Cambio hipsométrico de la vegetación en un bosque nublado de
los Yungas de Bolivia - metodología y cinturones altitudinales
Vegetational hypsometric change of a cloud forest in Bolivian Yungas
Methods and altitudinal belts
Kerstin Bach1'2 Stephan Robert Gradstein2
'Philipps-Uniyersitát Marburg, Fachbereich Geographie, Deutschhausstr. 10, 35037 Marburg, Alemania,
bach.kerstin@gmx.de
2Albrecht-von-Haller-Institut für Pflanzenwissenschaften, Abteilung Systematische Botanik,
Untere Karspüle 2, 37073 Góttingen, Alemania.
Se estudiaron los límites vegetales de cinturones altitudinales en un bosque nublado de los Yungas de Bolivia. Entre 1.700 y 3.400 m en una distancia altitudinal de 50 m, se analizaron 105 parcelas de 400 m2 en base a grupos elegidos (Pteridophyta, Melastomataceae, Araceae, Bromeliaceae, Palmae y Cactaceae). Se aplicaron diferentes métodos: Clasificación fítosociológica, análisis multivariado (DCA), de cluster y de parsimonia. Para un análisis de la diversidad beta (recambio de especies) se calculó el índice de Wilson-Shmida y se comprobó la significancia de los resultados con una simulación de la distribución de especies. En la clasificación fítosociológica se encontraron muchas transiciones, tanto entre grupos de parcelas como entre grupos de especies. En la DCA la mayoría de las parcelas formó una línea sin distancia amplia entre ellas. Solamente un grupo se distinguió por encima de los 3.050 m. Los resultados del análisis de cluster no fueron significativos y fue impossible distinguir grupos de parcelas en el análisis de parsimonia. Los índices de WilsonShmida se acercaron rara vez a un valor de uno y en ningún caso fueron significativos. La mayor concordancia de límites se encontró a 2.000 m (± 100 m) para los límites de distribución superiores de las especies y a 3.050 m. Sin embargo, no se pueden definir límites altitudinales precisos válidos para todos los grupos florísticos y armonizados con todos los métodos aplicados. Estos límites imprecisos reflejan la sinecología y autoecología de los grupos florísticos a lo largo de un transecto con cambios mayormente grad uales. Por ende, al momento de escoger un método de clasificación a lo largo de gradientes es muy importante tomar en consideración el objetivo del estudio.
Palabras clave: Bosques nublados, Cinturones altitudinales, Recambio de especies, Diversidad beta, PN-ANMI Cotapata.
Abstract
We studied elevational limits of vegetation belts in the Yungas of Bolivia, in montane cloud forest between 1,700 and 3,400 m based on an assessment of species diversity of six vascular plant groups (aroids, bromeliads, cacti, ferns, melastoms, and palms). We analysed the similarity of 105 relevees of 400 m2, taken at eleva tional intervals of 50 m. We applied phytosociological analysis (Tabwin), multivariate ordination (DCA), cluster analysis (Ward), and parsimony analysis (PA UP*). Species turnover was determined by means of the Wilson-Shmida index and sta tistica 1 significance by means of a Monte Carlo simulation of species distributions. Phytosociological analysis, cluster analysis, and parsimony analysis failed to resolve any significant elevational limits; while ordination revealed a major distance between groups of plots at 3,050 m. Significant borders at ca. 2,000 m (± 100 m) and at 3,050 m were based on a congruence of upper/ lower elevational limits of species ranges. Wilson-Shmida index values rarely approached 1.0 and none was significant. The study showed that sharply defined elevational belts for all groups of plants investigated could not be detected with the majority of the methods applied . The lack of borders presumably reflects the variation in species diversity patterns of the studied plant groups in relation to the gradual changes in abiotic factors along the gradient. Each group showed its own elevational pa tterns, presumably influenced by taxon-specific ecological traits. Also, the outcome of vegetation classification studies in species-rich tropical montane cloud forests seems to be strongly determined by applied methods and should be accurate with the study objectives.
Key words: Tropical montane cloud forests, elevational belts, species turnover, G-diversity, Cotapata National Park.
Introducción
Alexander von Humboldt inició la investigación sobre cinturones altitudinales en bosques montanos tropicales con su popular publicación en 1811. Posteriormente una enorme cantidad de clasificaciones con diferentes límites fue postulada (Frahm & Gradstein 1991). En general, estas clasificaciones se basan en investigaciones climáticas (p.e., Troll 1959, Lauer 1986) y/o en observaciones fisionómicas de la vegetación (p.e., Herzog 1923, Grubb 1974, Ellenberg 1975). Por la alta biodiversidad en bosques tropicales, solo escasos estudios se basan en características florísticas (Van der Hammen & Dos Santos 1995). Como terminología general se estableció una clasificación basada en Grubb (1974) recién utilizada por la UNESCO (Bruijnzeel & Hamilton 2000) con los siguientes pisos: Bosques de tierras bajas (en inglés: lowland forests), bosques montanos bajos (lozver montane forests), bosques nublados monta nos bajos (lower montane cloud forests), bosques nublados montanos superiores (upper montane cloud forests) y bosques nublados subalpinos (subalpine cloud forests). Los límites entre pisos varían en su altitud por diferentes factores (Frahm & Gradstein 1991, Bruijnzeel & Hamilton 2000). En primer lugar se menciona la influencia de la humedad con la latitud (distancia a la ITC), con la continentalidad (distancia al mar) y con el relieve orográfico. Además se describen como factores importantes el sustrato y la extensión máxima del monte. Recientes estudios no encuentran límites claramente marcados entre pisos altitudinales determinados por las comunidades vegetales (Lieberman et al. 1996, Vázquez & Givnish 1998, Kessler 2000). Se supone que la vegetación cambia gradualmente así como muchos factores abióticos a lo largo de un gradiente altitudinal y que solamente cuando existen discontinuidades abióticas también se presentan límites acentuados en la vegetación (Kessler 2000, Hemp 2002, Tuomisto eta!. 2003). El objetivo de este estudio es la descripción de los límites vegetales en un gradiente altitudinal, utilizando varios métodos de clasificación.
]]> Área de estudioEl área de estudio se ubica en el Parque Nacional y Área Natural de Manejo Integrado (PNANMI) Cotapata sobre la Cordillera Oriental, aproximadamente a 80 km al noreste de ciudad de La Paz. Ocupa diferentes formaciones montanas de vegetación en una topografía muy abrupta con pendientes entre 21 y 55%. El clima pertenece al tipo mesotérmico húmedo con lluvias de verano e inviernos secos y cálidos. Entre 2.300 y 3.400 m se observa la constante afluencia de neblinas todo el año.
Geológicamente, el área se caracteriza por rocas sedimentarias, de lutitas, limonitas y areniscas (Ribera 1995). Ribera eta!. (1994) describen tres zonas de vegetación incluidas en el PN-ANMI Cotapata: Páramo yungueño, ceja de monte y bosque montano húmedo. Una descripción más detallada del PN-ANMI se encuentra en Morales (1995) y también en Paniagua et al. (2003).
Para este estudio se eligió un transecto en el lado sureste del cerro Hornuni ( 3.647 m) entre 1.700 y 3.400 m (Bach et al. 2003). En la parte inferior de este transecto, el bosque montarlo húmedo está en un estado más o menos primario y se extiende por una vasta región. En la parte superior el transecto cruza un camino antiguo hacia una mina abandonada. La transición entre bosque y páramo se extiende en una parte amplia por encima de 3.150 m.
Métodos
Trabajo de campo
Por la alta diversidad florística en bosques montanos húmedos, la vegetación se estudió en base a grupos elegidos: Pteridophyta, Melastomataceae, Araceae, Bromeliaceae, Palmae y Cactaceae. Estos grupos representan en conjunto el 21.6% de la flora completa en bosques andinos (Van der Hammen Sz Dos Santos 1995) y como grupo tienen una correlación estrecha con la composición florística (r = 0,81; Bach 2004). Además, estos grupos tienen una distribución amplia en una variedad de hábitats (Bach et al. 1998 y Kessler tlz Bach 1999). A lo largo del transecto se estudiaron 105 parcelas homogéneas de 20 x 20 m en niveles de altitud cada (25-) 50 m. El área mínima de las parcelas se determinó por análisis de la relación entre especies y área a 2.000 m y 3.100 m (Bach et al. en prep.). En casos en que no fue posible establecer una parcela rectangular, p.c. por la influencia de árboles caidos, se investigó un área cercana homogénea de 400 m2. La distancia altitudinal entre parcelas fue de 50 m en general. Sin embargo, en el bosque montano con vegetación densa y alta diversidad reducimos la distancia a 25 m y contrariamente en la parte superior del transecto se registró un salto altitud mal de 100 m por la falta de bosque continuo. En el bosque montano fue posible investigar tres transectos paralelos. Además, a tres niveles ( 1.850 m, 2.550 m, 3.000 m) se estudiaron parcelas en un tramo horizontal con exposiciones oeste hasta noreste (Fig. 1). Para el análisis de la diversidad florística es importante (i) verificar que el área total de cada parcela investigada sea siempre el mismo tamaño ( 400 m2 en este estudio) e (ii) incluir una mínima cantidad de parcelas en cada intervalo altitudinal (por lo menos cinco). En cada parcela se anotó la cobertura de especies encontradas según la escala clásica de Braun-Blanquet y la cobertura y estructura de la vegetación en general.
Análisis de los datos
Para determinar los límites entre formaciones vegetales se aplicaron los siguientes métodos: Clasificación fítosociológica, análisis multivariado (DCA), análisis de cluster, análisis de parsimonia y análisis de la diversidad beta (recambio de las especies). La clasificación fítosociológica se realizó según el método de Mueller-Dombois Sz Ellenberg (1974). El objetivo de este método es generar una tabla comparativa con grupos de parcelas y de especies, las cuales representan las formaciones vegetales con especies características. Para el análisis multiva hado se eligió una ordenación (análisis de componentes principales) del tipo Detrended Correspondence Analysis (DCA). Este método corrige las agrupaciones en herradura, los resultados típicos de ordenaciones simples a lo largo de gradientes. Se utilizó un análisis de cluster, calculando la distancia euclideana entre las parcelas y varianza mínima entre los grupos (Ward). Este análisis siempre produce agrupaciones a causa de su fundamento matemático. Por eso es importante comprobar la
significancia de estos grupos, método diseñado por Pillar (1999). Otra metodología para comprobar la significancia de las agrupaciones es el análisis de parsimonia. Este método se origina por agrupaciones de especies en un sistema filogenético y recientemente se está aplicando a estudios biogeográficos. Igual que con las características genéticas de las especies en un análisis filogenético, se ordenan las parcelas por presencia o ausencia de especies en una tabla. Esta tabla se analiza con el programa PAUP (Swofford 2001) y da como resultado un cladograma, el cual comprende una agrupación de las parcelas con un mínimo de ramificaciones. Las dicotomías terminales representan las parcelas que alojan especies exclusivas en otras parcelas. En general existen varias posibilidades para la construcción de un cladograma con un mínimo de ramificaciones. Por eso se construye al final un promedio árbol de consenso estricto (en inglés: strict consensus tree) y la significancia de cada ramificación se calcula con un procedimiento de "atar con correa" (en inglés: boostrap) para inferencia estadística, utilizada para estimar la distribución de muestreo con el reemplazo de la muestra original.
El análisis de la diversidad beta muestra el recambio en la composición de las especies entre hábitats. En este estudio se analizó el cambio entre intervalos altitudinales con distancia de 100 m y de 200 m. Para cada intervalo se contó el número total de especies, número de estas especies con límite superior de distribución por intervalo y número de especies con límite inferior de distribución por intervalo. Para estos límites se calificaron los registros mínimos y máximos de cada especie a lo largo del transecto sin contar intervalos vacíos, porque frecuentemente existen otros motivos de su ausencia aparte de las razones bióticas. Además, se calculó el índice de Wilson-Shmida entre intervalos vecinos (Wilson & Shmida 1984):
b + c
R t = 2a + b c
Donde, 'a' significa el número de las especies presentes en ambos intervalos, 'b' representa el número de especies exclusivas del intervalo inferior (con límite superior de distribución/ intervalo) y 'c' es el número de especies exclusivas del intervalo superior (que inician su distribución en este intervalo). Cuando este índice se acerca a un valor de uno existen pocas especies comunes, es decir, se muestra un cambio marcado. Para excluir efectos artificiales en los bordes del transecto no se tuvieron en cuenta los intervalos extremos en la parte superior e inferior del transecto. Por ejemplo, el número de límites inferiores de las especies al borde inferior del transecto se refiere al límite artificial del transecto y no al límite natural de las especies. Además, no se consideraron las especies que se encontraban en sólo un intervalo. Este análisis se realizó en tres niveles: Primero todas las especies juntas, segundo los grupos florísticos por separado y en último lugar, diferenciando especies epífitas de especies terrestres. Las palmeras y cactáceas no se tomaron en cuenta por tener un número muy reducido de especies.
Para comprobar la significancia de los resultados se aplicó una simulación de distribución de especies en base a los datos observados con una prueba de Monte Carlo. Fue necesa ria la simulación porque no existe un modelo matemático que muestre el patrón de la distribución de especies en gradientes altitudinales. Por eso, para cada intervalo altitudinal en la simulación se refiere a un número máximo de especies que presenta el número registrado en el campo estimado como la diversidad alfa según el procedimiento de MMMean. Este método se base en la fórmula de Michaelis-Menten y da como resultado un promedio de varias repeticiones. En este estudio se calcularon 1.000 repeticiones con el programa de computadora EstimateS 6.0b1 (Colwell 1997).
En el modelo, una cantidad de especies, como observadas en el campo, se coloca aleatoriamente por orden y por posición altitudinal. La distancia máxima de distribución (del límite inferior hasta el superior) por cada especie se refiere al estado observado en el campo. No está permitido colocar más especies en un intervalo que el máximo definido por el procedimiento de MMMean. En último lugar, se cuenta la distribución de las especies por intervalo (número total, número de límites superior e inferior) y se calcula directamente el índice de Wilson-Shmida entre intervalos vecinos. Para obtener promedios y desviaciones estándar para cada intervalo, la simulación se repite 1.000 veces (prueba de Monte Carlo). Con estos promedios se compararon las distribuciones observadas mediante una prueba de X2 (acumulaciones) y prueba de Kolmogoroff-Smirnov (valores del índice de Wilson-Shmida).
Resultados
]]> Seencontra ron 342 especies de los gru pos elegid os en el área de estudio: 228 Pteridophyta, 64 Melastoma tacea e, 22 Araceae, 21 Bromeliaceae, 5 Palmae y 2 Cactaceae. Dentro de las Pteridophyta la diversidad más alta se observó en las Lomariopsidaceae (Elaphoglossum spp.) y Grammitidaceae. El número más alto de especies se registró entre 1.900 y 2.000 m.La clasificación fitosociológica comprende 129 especies con una frecuencia 10% y 109 especies con una frecuencia 1%. Solamente cinco especies aparecieron en más del 50% de los relevamientos (Blechnum lechleri, Hymenophyllum fucoides, H. plumieri, Melpomene firma y Polypodium fraxinifolium). En la tabla ordenada no se reconocieron tipos claramente agrupados con especies características. Más bien se evidenció un ordenamiento gradual de altura (Anexo 1). Se encontraron muchas transiciones, tanto entre grupos de parcelas como entre grupos de especies. Solamente se distinguió claramente la composición florística en los extremos del transecto. En la parte inferior se encontraron especies que aparecen también en bosques secos (Anthurium paraguayense, Asplenium bangii, Blechnum occidentale y Epiphyllum phyllanthus) y encima del límite de bosque se diferenciaron relevamientos por la presencia de especies exclusivas (p.e., Miconia mandonii, Elaphoglossum mathewsii, Lellingeria pseudocapillarís y Melpomene pilosissima).
Este grupo de relevamientos por encima del límite de bosque se encontró dentro de parcelas claramente separadas del resto en el análisis multivariado (DCA) (s3, Fig. 2). Sin embargo, el límite inferior de este grupo a 3.050 m coincide parcialmente con el grupo s2 (2.900- 3.075 m). Además, se separó a lo largo del eje uno y dos un grupo (Si) que comprende parcelas con diferentes exposiciones sobre el tramo horizontal a 3.000 m. La parte superior del bosque (s2) no se dividió claramente de las otras parcelas inferiores (hm, mo), más bien formó una línea a lo largo del eje 1 sin distancia amplia entre relevamientos. La distinción entre grupos inferiores ('hm' y 'mo') se realizó de manera subjetiva. El valor propio del eje 1 se expresa a 0.69 con una longitud de 5.77, lo que significa que se realiza un solo recambio completo entre los extremos del transecto (Leyer & Wesche 2007).
En el análisis de cluster se mostraron tres grupos a 20% de la distancia máxima entre relevamientos. Estos grupos comprenden parcelas que se colocan a altitudes como sigue: 1.700- 2.400 m, 2.350- 2.850 m y 2.800- 3.400 m. Sin embargo, la significancia de estos grupos fue negativa según la prueba de Pillar (P = 0.092).
El análisis de parsimonia resultó en un árbol de consenso estricto de 189 cladogramas con 1.956 pasos, el cual mostró sólo dos ramas robustas con bootstrap > 70%. Sin embargo, comprendió en ambos lados solamente dos parcelas vecinas y todas las parcelas restantes se agruparon como indistinguibles.
La diversidad beta calculada en base al índice de WilsonShmida con intervalos de 100

Fig. 2: Ordenación de las parcelas en el DCA proyectado con el primer y segundo eje. Los
grupos marcados comprenden parcelas en las alturas siguientes: mo (1.700- 2.300 m),
hm (2.300- 2.850 m), si (2.820-3.015), s2 (2.900- 3.075 m) y s 3 (3.050- 3.400 m). Las parcelas
]]> del grupo s l se encuentran en su mayoría en el tramo horizontal a 3.000 m con diferenteexposición.
m mostró valores menores a 0.17 en promedio y máximos entre 0.22 y 0.71 (Fig. 3). Los valores máximos, que indican cambios marcados en la composición de especies, se encontraron a 2.600 y 2.800 m, principalmente para especies terrestres (incluyendo especies terrestres de Pteridophyta, Melastoniataceae, Araceae y Bromeliaceae) y también para Bromeliaceae epífitas. Sin embargo, la mayoría de las epífitas tenía máximo del índice de WilsonShmida a 3.000 m (incluyendo epífitas de Pteridophyta y especies de Elaphoglossum). Los máximos para todas las especies juntas reflejan estas altitudes: 2.600 m, 2.800 in y 3.000 ni. Además se mostraron máximos a 2.300 m (Araceae), 2.400 ni (Hymenophyllaceae) y 3.200 ni (Melastomataceae). Con intervalos de 200 m resultaron valores del índice de WilsonShmida un poco más elevados que en el análisis con intervalos de 100 m (0.25 en promedio y máximos entre 0.4 y 0.7). Los valores máximos no fueron presentados por epífitas, excepto Bromeliaceae a 2.700- 2.800 m. La mayoría de los grupos analizados presentó valor máximo a 2.700 m (todos los grupos, Pteridophyta, terrestres y terrestres de Pteridophyta). Los valores máximos se encontraron a 2.300 ni (Araceae), 2.400-2.500 (Hymenophyllaceae) y 2.800- 2.900 m (Melastomataceae).
La acumulación de límites de distribución inferior fue máxima en niveles altitudinales menores a 2.400 ni con intervalos de 100 ni (Fig. 4). Por encima de esa altitud, el número de límite inferior disminuyó lentamente de manera continua para todos los grupos. Los límites de distribución superior no mostraron un patrón opuesto, sino fluctuaron con varios máximos de acumulación, reflejando diferentes extensiones de distribución de las especies (Fig. 5). Las especies terrestres mostraron un máximo
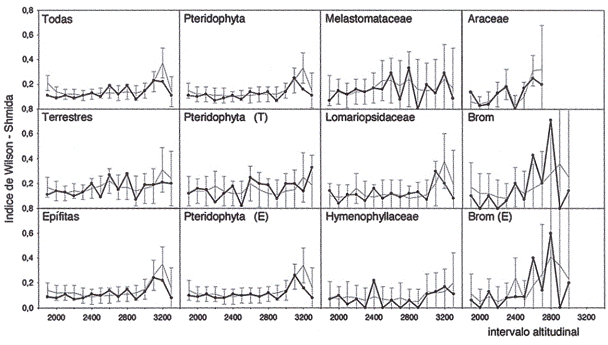
Fig. 3: Índice de Wilson-Shmida (diversidad beta) a lo largo del transecto con intervalos
altitudinales de 100 m (línea negra = valores observados, línea gris = valores media de
la simulación con desviación estándar doble, Brom = Bromeliaceae, E = epífitas, T =
especies terrestres).
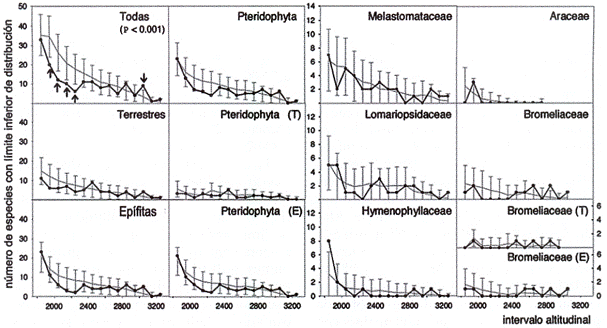
con intervalos de 100 m (línea negra = valores observados, línea gris = valores de la
simulación con desviación estándar doble, E = epífitas, T = especies terrestres). Las
flechas indican intervalos altitudinales con divergencias significantivas entre valores
observados y simulados.
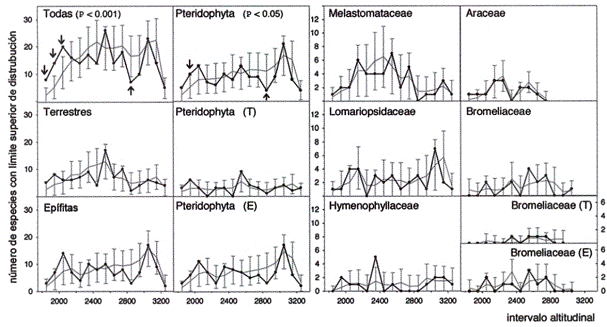
Fig. 5: Acumulación de los límites de distribución superior de las especies a lo largo del
transecto con intervalos de 100 m (véase Fig. 4 para las abreviaciones).
a 2.550 m, así como las Bromeliaceae. Las epífitas tenían máximos a 2.050 y 3.050 m, comparable a los máximos del índice de WilsonShmida. Las Melastomataceae presentaron varias acumulaciones elevadas entre 2.150 y 2.750 m y las Araceae entre 2.150 y 2.250 m.
Sin embargo, comparando estos valores del índice de Wilson-Shimda con los resultados de las simulaciones (prueba de Kolmogoroff-Smirnov), se mostró que todos los cambios mencionados arriba no difieren significativamente del caso aleatorio. Para los límites de distribución superior de especies con intervalos de 100 m se encontraron solamente acumulaciones significativas para todas las especies juntas (P < 0.001) y Pteridophyta (P < 0.05). Para los límites de distribución inferior se confirmaron significativamente sólo las acumulaciones para todas las especies juntas (P < 0.001). Además, con intervalos de 200 m resultaron significativas no más que dos acumulaciones: Límite superior de todas las especies juntas (P < 0.05) y de epífitas (P < 0.05). En relación con las Araceae, Bromeliaceae y Melastomataceae no se encontraron límites significativos de la diversidad beta a lo largo del transecto (Tabla 1).
]]> DiscusiónTodos los resultados según diferentes métodos aplicados indican claramente el carácter gradual del transecto en el área de estudio. Los índices de Wilson-Shmida se acercaron rara vez al valor de uno y en ningún caso fueron significativos. La tendencia de valores más altos con intervalos de 200 m refleja la heterogeneidad más alta dentro y entre los intervalos más amplios. Pero tampoco se mostraron significativos.
La mayor concordancia de límites se encuentra a 2.000 m (± 100 m) y a 3.050 m (Tabla 1). Según la hipótesis sobre gradientes (Kessler 2000, Hemp 2002, Tuomisto et al. 2003), se supone que a estas altitudes existen discontinuidades paralelas con los factores abióticos. Efectivamente y comparando la diversidad beta y factores edafológicos, se corroboran relaciones significativas con la saturación de bases en el horizonte A (r = 0.93; P < 0.05 corregido de Bonferroni). Sin embargo,
Tabla 1: Cuadro sintético de resultados según diferentes métodos aplicados que consideran estos intervalos altitudinales donde se comprobó el significativo cambio de la composición de especies (R, = diversidad beta).

existe la hipótesis que plantea que los factores efectivos a lo largo de gradientes no son los mismos en los límites opuestos (Dobzhansky 1950, McArthur 1972, Grubb 1977, Kaufman 1995, Stohlgren Bachand 1997). Se supone que en los límites superiores los factores bióticos tienen mayor efecto y que en los inferiores el factor más importante corresponde a las condiciones abióticas. Tenemos indicios que en el área de estudio los factores abióticos fueron la influencia más fuerte en la parte inferior del transecto; a esa altitud, el tipo de suelo cambia de cambisol a podzol y empieza la afluencia de neblinas. En consecuencia, la humedad aumenta y disminuye la temperatura junto con la radiación. Es posible que en las Hymenophyllaceae el máximo del índice de WilsonShmida a 2.400- 2.500 m refleje el recambio entre especies con tomento, adaptadas a la aridez y especies más cutáneas. Además, se mostró que los límites de distribución inferior de las especies (que indican factores bióticos según esta hipótesis) se acumularon a esta altitud con una cantidad significativa menor que los valores simulados. Es decir, el recambio es menor contra la previsión por la simulación.
Al contrario, en el límite opuesto del gradiente, a 3.050 m, se encontró una acumulación de límite inferior significativamente mayor que los valores simulados (lo que indica factores abióticos según esta hipótesis). A esta elevación la acumulación de humedad en el suelo bajo condiciones casi pantanosas se realiza de manera corriente y no tan abrupta como a 2.200 m. Además, tenemos todavía el efecto de la neblina hasta los 3.400 m y posiblemente la acumulación significativa del límite inferior muestra en este caso un límite biótico de especies del páramo, las cuales no se mantienen frente a especies del bosque húmedo montan° (competencia entre especies, cantidad de los nichos disponibles fuera y dentro del bosque). Por otra parte, se podría tratar también de un límite abiótico cuando se toma en cuenta el cambio de la radiación.
En conclusión, no se pueden definir límites altitudinales precisos válidos para todos los grupos florísticos y armonizados con todos los métodos aplicados. Estos límites imprecisos reflejan la sinecología y autoecología de los grupos florísticos a lo largo de un transecto con cambios mayormente graduales. Esto apoyaría la hipótesis de Gleason (1926) en un contexto biogeográfico frente a la de comunidades discretas apoyada por BraunBlanquet (1964).
Los varios máximos de acumulaciones de límites de distribución inferior de las Melastomataceae indican el cambio sucesivo entre especies de diferentes hábitats a lo largo de un transecto altitudinal. Al contrario, los máximos de las Araceae muestran la distribución principal de este grupo en regiones calientes húmedas con pocas especies en áreas altimontanas.
Sin embargo, las diferencias de los tipos vegetales se observan cualitativamente en el campo también por la estructura del bosque (Fig. 6). Sin determinar los límites precisos, se distinguen tres tipos los cuales obedecen a la clasificación de la UNESCO (bosque nublado montano bajo - lozver montanecloud forestbosque nublado montano superior- upper montane cloud forest- y bosque nublado subalpino, subalpine cloud forest). En la terminología de Beck et al. (1993) y Paniagua et al. (2003), esos tipos se refieren al bosque húmedo montan°, bosque nublado y ceja de monte de Yungas. Navarro (2002) dividen el bosque nublado en dos tipos (altimontano y basimontano) dentro de bosques húmedo hiperhúmedos pluviales de los Yungas de Coroico. Ribera (1995) también describe sólo dos formaciones bajo el páramo yungueño (bosque nublado en ceja de monte y bosque húmedo montañoso de Yungas) con una transición perhúmeda y de neblina amplia (Fig. 7). En el cerro Hornuni, la ceja de monte (aproximadamente grupo s, en el análisis de DCA) se extiende en una región angosta, dando lugar a la transición amplia entre bosque y páramo por encima de los 3.050 m (grupo s3).
]]>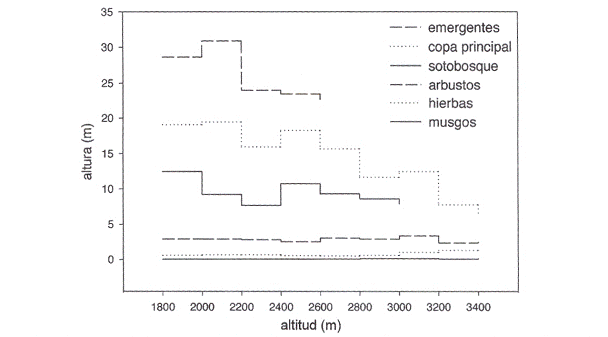
Fig. 6: Estructura del bosque a lo largo del transecto calculando los promedios en intervalos de 200 m. Se distingue en la parte inferior un bosque con tres estratos (1.700- 2.600 m), seguido por un bosque con dos estratos (2.600- 3.000 m) yen último lugar una vegetación con sólo un estrato de árboles (3.000- 3.400 m).
Paniagua et al. (2003) describen ese tipo de vegetación como subpáramo yunguerio.
Para la parte opuesta del transecto, inferior al bosque húmedo montan°, Navarro (2002) describen un bosque húmedo pluviestacional subandino de los Yungas, que reemplaza al bosque pluvial yunguerio subandino en lugares subhúmedos. Efectivamente, las especies de lugares más secos (veáse arriba) indican la transición a este tipo pluvioestacional del fondo del valle Huarinilla. Las especies típicas según Navarro (2002) estuvieron presentes solo con pocos individuos (a excepción de Cyathea delgadii, Dictyocaryum lamarckianum con solo un individuo) - comparando las especies de los grupos considerados en este estudio.
Conclusiones
Estas diferencias en la descripción de los cinturones tanto en la terminología como en la altitud correspondiente reflejan los problemas en determinar exactamente los límites de las formaciones de vegetación. Mientras que un investigador evalúa la estructura, otro se refiere a relevamientos en zonas típicas y analiza parcelas separadas por una gran distancia altitudinal (más de 200 m). Un tercero utiliza métodos no apropiados para gradientes como el análisis de cluster. Pese a que se entiende el deseo de delimitar en forma precisa a las formaciones, es imposible ajustarlas a cambios graduales como en el área de estudio. Por ende, al momento de escoger un método de clasificación a lo largo de gradientes es muy importante tomar en cuenta el objetivo del estudio. Si es para orientarse principalmente en el área, el análisis de cluster se presta adecuadamente. En caso de comparar la composición florística con factores abióticos, entonces es favorable aplicar el análisis de componentes principales (DCA). A fin de empezar a describir los límites con mayor nitidez, se impone un análisis de parsimonia o calcular la diversidad beta considerando

Fig. 7: Límites altitudinales de la vegetación en el área de estudio por diferentes autores y
métodos, 11 resp. U indican los límites significativos de las distribuciones superior
e inferior. Las áreas rayadas marcan zonas de transición. LI Vegetación de los Yungas
]]> subandinos pluviestacionales.Véase fig.2 para las abreviaciones.una prueba de significancia. Una clasificación fitosociológica con tabla ordenada muestra claramente el carácter gradual altitudinal. La fuerza de este método se encuentra más en relación con los objetivos sin transiciones y se necesitan más investigaciones si es adecuado utilizarlo en regiones con diversidad tan elevada.
Agradecimientos
Este trabajo forma parte del proyecto conjunto entre la Universidad de Góttingen y el Instituto de Ecología de La Paz "Cambio hipsométrico de ecosistemas de bosques montañosos húmedos en los Yungas de Bolivia" financiado por la DFG. Agradecemos por las identificaciones A.R. Smith (Pteridophyta), M. Kessler (Cactaceae y Elaphoglossum), T.B. Croat y A. Acebey (Araceae), H.E. Luther y T. Krómer (Bromeliaceae), S. Renner y F. Almeda (Melastomataceae), M. Moraes (Palmae), M. Lehnert (Cyathea) y J. Gonzales (Sticherus). Ala comunidad del Chairo y todos los estudiantes y científicos por su ayuda en el campo. Al personal del Herbario Nacional de Bolivia en La Paz por el apoyo logístico y al revisor por sus notas valiosas.
Referencias
Bach, K. 2004. Vegetationskundliche Untersuchungen zur Hóhenzonierung tropischer Bergregenwálder in den Anden Bol iviens. Górich Weiersháuser, Marburg. 123 p.
Bach, K., M. Kessler & J. Gonzales. 1998. Clasificación preliminar de los bosques deciduos andinos de Bolivia en base a grupos indicadores botánicos. Ecología en Bolivia 32: 47-65.
Bach, K., M. Schawe, S. R. Gradstein, G. Gerold, S. Beck & M. Moraes. 2003. Vegetación, suelos y clima en los diferentes pisos altitud males de un bosque montan() de Yungas, Bolivia: Primeros resultados. Ecología en Bolivia 38: 3-14.
Bach, K., S. G. Beck, N. Thompson &A. Palabral. en prep. Species-area relationship at an elevational gradient in the Bolivian Andes. Ecotropica.
Beck, S. G., T. J. Killeen & E. García E. 1993. Vegetación de Bolivia. pp. 6-24. En: T. J. Killeen, E. García E., S. G. Beck (eds.). Guía de Árboles de Bolivia. Herbario Nacional de Bolivia, Missouri Botanical Garden, Edit. Quipus, La Paz.
]]> Braun-Blanquet, J. 1964. Pflanzensoziologie. Springer, Viena. 865 p.Bruijnzeel, L. A. & L. S. Hamilton 2000. Decision time for cloud forests. UNESCO, Paris.
39p.
Colwell, R. K. 1997. EstimateS: statistical estimation of species richness and shared species from samples, version 6. 10nline.] Available at / /purl.ocic. org/estimates.
Dobzhansky, T. 1950. Evolution in the tropics.
American Scientist 38: 209-211.
Ellenberg, H. 1975. Vegetationsstufen in perhu miden bis perariden Bereichen der tropischen Anden. Phytocoenologia 2(3/4): 368-587.
Frahm, H.-J. & S. R. Gradstein 1991. An altitudinal zonation of tropical rain
forests using bryophytes. Journal of Biogeography 18: 669-678.
Gleason, H. A. 1926. The individualistic concept of the plant association. Bull. Torrey Botan. Club 53, 7-26.
]]> Grubb, P. 1974. Factors controlling the distribution of forest types on tropical mountains: New facts and new perspective. pp. 13-46. En: J. R. Flenley (ed.). Altitudinal Zonation in Malaysia . Trans. 3rd Aberdeen-Hull Symp. on Malaysian Ecology. Univ. Hall, Dept. Geogr., Miscell. Ser. 16.Grubb, R 1977. Control of forest growth and distribution on wet tropical mountains: with special reference to mineral nutrition. Ann. Rey. Ecol. Syst. 8: 83-107.
Hemp, A. 2002. Ecologyof the pteridophytes on the southern slopes of Mt. Kilimanjaro. I. Al titudinal distribution. Plant Ecology 159(2): 211-239.
Herzog, T. 1923. Die Pflanzenwelt der bolivischen Anden und ihres óstlichen Vorlandes. En: A. Engler & O. Drude (eds.). Die Vegetation der Erde, Bd. 11, Leipzig.
Humboldt, A. v. 1811. Atlas geographique et physique du royaume de la NouvelleEspagne: Ifonde sur des observations astronomiques, des mesures trigonometriques et des nivellemens barometriquen Paris.
Kaufman, D. M. 1995. Diversity of New World mammals: universality of the latitudinal gradient of species and bauplans. Journal of Mammalogy 76: 322-334.
Kessler, M. 2000. Altitudinal zona tion of Andean cryptogam communities. Journal of Biogeography 27: 275-282.
Kessler, M. & K. Bach. 1999. Using indicator families for vegetation classification in species-rich Neotropical forests. Phytocoenologia 29(4): 485-502.
Lauer, W. 1986. Die Vegetationszonierung der Neotropis und ihr Wandel seit der Eiszeit. Ber. Deutsch. Bot. Ges. 99: 211-235.
Leyer, I. & K. Wesche. 2007. Multivariate Statistik in der Ókologie. Springer, Berlin. 221 p.
]]> Lieberman, D., M. Lieberman, R. Peralta & G. S. Hartshorn. 1996. Tropical forest structure and composition on a largescale altitudinal gradient in Costa Rica . Journal of Ecology 84: 137-152.McArthur, R. H. 1972. Geographical ecology: patterns in the distribution of species. Harper and Row, Nueva York. 269 p.
Morales, C. B. (ed.) 1995. Caminos de Cotapata. Instituto de Ecología, La Paz. 174 p.
Mueller-Dombois, D. & H. Ellenberg 1974. Aims and methods of vegeta tion ecology. Wiley & Sons, Nueva York. 547 p.
Navarro, G. 2002. Provincia biogeográfica de los Yungas.Pp.279-350.En:Navarro,G.&M. Maldonado (eds.). Geografía Ecológica de Bolivia. Centro de Ecología Simón I. Patirio - Departamento de Difusión, Cochabamba.
Paniagua,N.,C.MaldonadoG.&C.Chumacero. 2003. Mapa de vegetación de los alrededores de la Estación Biológica de Tunquini, Bolivia. Ecología en Bolivia 38: 15-26.
Pillar, V. D. 1999. How sharp are classifications? Ecology 80(8): 2508-2516.
Ribera, M. O., M. Liberman, S. Beck & M. Moraes R. 1994. Mapa de vegetación y áreas protegidas de Bolivia. Escala 1:1.000.000. La Paz.
Ribera, M.O. 1995. Aspectos ecológicos, del uso de la tierra y conservación en el Parque Nacional y Área Natural de Manejo
Artículo recibido en: Enero de 2005. Manejado por: Mónica Moraes. Aceptado en: Marzo de 2007.
]]> Integrado Cotapata. pp. 1-84. En: C. d. Morales (ed.). Caminos de Cotapata. Instituto de Ecología, La Paz.Stohlgren T. J. & R. R. Bachand . 1997. Lodgepole pine Pinus conforta ecotones in Rocky Mountain National Park, Colorado, USA . Ecology 78: 632-641.
Swofford, D. L. 2001. PAUP* Phylogenetic analysis using parsimony (*and other methods). Version 4. Sinauer Associates, Sunderland, Massachusetts.
Troll, C. 1959. Die tropischen Gebirge - ihre dreidimensionale klimatische und pflanzengeographische Zonierung. Bonner Geographische Abhandlungen 25.
Tuomisto, H., K. Ruokolainen, M. Aguilar & A. Sarmiento. 2003. Floristic patterns along a 43-km long transect in an Amazonian rain forest. Journal of Ecology 91(5): 743-756.
Van der Hammen, T. & A. G. Dos Santos (eds.) 1995. La Cordillera Central Colombiana. Transecto Parque Los Nevados. Studies on tropical andean ecosystems. Studies on Tropical Andean Ecosystems (ECOANDES). Vol. 4. Cramer, Berlin. 613 p.
Vázquez, J. A. & T. J. Givnish. 1998. Altitudinal gradients in tropical forest composition, structure and diversity in the Sierra de Manantlán. Journal of Ecology 86(6): 999-1020.
Wilson, M. V. & A. Shmida. 1984. Measuring beta diversity with presence-absence data. Journal of Ecology 72: 1055-1064.



Anexo 2
Alsophila erinacea 9:+, 10:1, 13:1, 16:+, 25:+, 35:1; Anthurium paraguayense 6:+, 7:+, 15:+, 20:+, 23:+; Anthurium scandens var. scandens 13:r, 22:+, 24:+, 36:+, 37:+, 45:+, 67:+, 72':+; Anthuritun soukupii 11:+; Asplenium abscissum 2:+; Asplenium (dation 5:1; Asplenium bangii 2:+, 3:+, 5:1, 13:+, 20:+, 21:1; Asplenium cuspidatum 2:+, 5:1, 9:+, 10:+, 14:+, 21:+; Asplenium cuspidatum vel aff. 12:+, 16:+; Asplenium flabellulaturn 5:+; Asplenium fragrans 76:+, 86:+; Asplenium raddianum 1:1, 8:+, 24:+; Asplenium rigídum 35:1, 53:+, 57:+; Asplenium rutaceum 9:+; Asplenium sp. ]]> ' ; Elaphoglossum paleaceum ve! aff. 85:+, 100:1; Elaphoglossum cf. pannosum 27:+; Elaphoglossum petiolosum 58:+, 69:+; Elaphoglossum puberulentum 9:+, 10:+, 12:+, 13:1, 16:+; Elaphoglossum pulchrum 26:+; Elaphoglossum cf. rimbachii 86:+, 91:1, 100:+, 103:+; Elaphoglossumsrnithii vel aff. 1:+, 3:+, 7:+, 17:+; Elaphoglossum sp. nov. (E 4455) 54:+; Elaphoglossum sp. nov. aff. amplum 66:+, 71:+, 72:+, 76:+, 85:1, 88:+, 89:+, 103:1; Elaphoglossum tabanense 1:+, 3:+, 4:+, 7:+, 8:+, 15:1, 21:+, 24:+, 29:+; Elaphoglossum unduaviense ve! aff. 84:1, 104:1, 105:1; Eriosorus orbignyanum 27:+, 45:+, 46:+, 49:+, 50:+, 51:+, 64:+, 66:+; Geonoma densa 66:+, 76:+, 79:+, 80:1; Graffenrieda cf. gracilis 8:+, 20:+; Granunitis bryophila 51:+, 62:+, ]]> ' phylla 25:+; Pteris sp. 4:+; Racinaea pardina 32:1, 36:+, 40:1; Racinaea tetrantha var. scarlatina 76:1, 81:1, 88:+, 95:+, 100:+; Radiovittaria gardneriana 84:+, 87:1, 90:+, 95: r, 98:+; Radiovittaria stipitata 14:+, 18:+, 21:+, 22:+, 26:+, 27:+, 29:+, 31:+; Saccoloma inaequale 29:+; Selaginella moritziana var. moritziana 54:1, 57:+, 58:+, 59:1, 66:+, 75:+, 78:+; Selaginella ]]>
]]>