Modelización de la evolución de la materia orgánica en suelos en descanso (Altiplano seco boliviano)
Modelling the transformations of soil organic matter in fallow (dry Bolivian Altiplano)
Pierre Bottner1, Dominique Hervé2 , Ruben Callisaya3, Klaas Metselaar4 & Marc Pansu2
1CEFE-CNRS, 1919 route de Mende, 34293 Montpellier Cedex 5, Francia, email: bottner.pierre@wanadoo.fr
2IRD, 911 Avenue Agropolis BP 64501, 34394 Montpellier Cedex 5, Francia, email: herve@mpl.ird.fr, pansu@mpl.ird.fr ]]>
Resumen
La paja de trigo marcada con 14C y 15N ha sido incubada durante dos años en el horizonte 5-10 cm de un suelo arenoso luego de un descanso de tres años (Jo por joven) y de ocho años (Vi por viejo) en el Altiplano central de Bolivia. Dos calidades de paja han sido utilizadas: N- con N = 0.3% , N+ con N = 1.6%. Mediciones en nueve fechas de 14C, C, 15N y N totales del suelo proveen datos para el modelo de descomposición de la materia orgánica MOMOS-6 con cinco compartimientos. Se calcula la tasa de N de cada compartimiento, dividiendo su tasa de C por su tasa C/N. En la entrada del modelo, la necromasa (NC) se divide en material vegetal lábil (VL) y estable (VS) en base a sus características bioquímicas; carbono y nitrógeno que alimentan a la biomasa microbiana (BM) y que a su vez alimenta a los compartimientos del humus lábil (HL) y estable (HS). Las salidas son el CO2 producido por BM y el nitrógeno mineral. Cada constante de velocidad se multiplica por una función de respuesta a la temperatura y humedad del suelo. El modelo SAHEL predice esta humedad. Después de dos años de incubación para ambas calidades de paja, no subsistía más que un 1/4 del 14C inicial. Con N+, el 60-70% y con N- el 55% del 14C inicial se mineralizó durante los 100 primeros días. Después de dos años, subsistió el 45 y 60% del 15N inicial en el suelo para N+ y N-, respectivamente. La edad del descanso tiene un efecto mucho menos importante que el tenor en N de la paja que frena la mineralización de 15N, pero también la mineralización inicial de 14C. La biomasa microbiana predicha por el modelo llega inicialmente a 30 y 13% del 14C total y 75 y 50% del 15N total para las pajas N+ y N-, respectivamente. Durante los primeros 100 días, estos valores bajaron en ambos tipos de paja menos del 8% para 14C y entre 10-12% para 15N. Durante esta fase inicial, 55-65% de 14C y 65-75% de 15N remanentes, se acumulan progresivamente en el humus lábil como fuente de C y N para los microorganismos y de nitrógeno mineral para las plantas. El humus estable no excede nunca en el 2% de 14C ni en el 2.5% de 15N remanentes. El coeficiente metabólico de la biomasa microbiana (q14CO2) fue más elevado con N- que con N+, por su demanda más importante en energía de mantenimiento.
Palabras clave: Altiplano, Materia orgánica, Modelización, 14C, 15N.
Abstract
14C and 15N labelled wheat straw was incubated over 2 years in the 5-10 cm layer of a sandy soil, under a young 3 year old fallow plot ( =Jo) and an older 8 year old one (= Vi), in the central Bolivian Altiplano. Two kinds of straw were used: with N = 0.3% called N- and with N = 1.6% called N+. The analyses of total 14C, C, 15N and N at nine sampling dates were used in a five-compartment model of organic decomposition, MOMOS-6. The N content of each compartment was calculated using the C content divided by its C/N ratio. The model input (necromass = NC) is split into labile (VL) and stable (VS) plant material compartments, using the biochemical characteristics of NC. VL and VS are used by the microbial biomass (BM) which provides labile (HL) and stable humus (HS) compartments with C and N. The outputs are CO2 released from BM and inorganic N. Each decomposition rate is multiplied by a temperature- and a soil moisture- response function. The soil moisture was predicted using SAHEL model. For both straw types, only 1/4 of the initial 14C remained in the soil after two years of incubation. With the N+ treatment, 60-70 % and with N- 55 % of the initial 14C were mineralised within the first 100 days. After 2 years of incubation, 45 and 60% of the initially added 15N remained in the soil for N+ and N-, respectively. Thus the N content of the straw lowered the mineralisation of 15N but also the initial mineralisation of 14C. The fallow age affected the decomposition rates much less than the straw quality. At the beginning of incubation, the predicted microbial biomass amounted to 30 and 13% of the remaining 14C and 75 and 50% of the remaining 15N for N+ and N- respectively. During the first 100 days, these values decreased to less than 8% for 14C and to 10-12% for 15N for both straw types. During this initial phase, 55-65% of the remaining 14C and 65-75% of the remaining 15N, accumulated progressively in the labile humus compartment (HL), which became the essential source of 14C and 15N for the micro-organisms and of inorganic N for plants. The stable humus compartment (HS) never exceeded 2% of the remaining 14C and 2.5 % of the remaining 15N. The microbial metabolic quotient (q 14CO2) was higher with the N- treatment than with N+, due to a higher demand of maintenance energy for N-.
Key words: Altiplano, Organic matter, Modelling, 14C, 15N.
De la paille de blé marquée au 14C et 15N a été incubée durant 2 ans dans l’horizon 5-10 cm d’un sol sableux sous une jachère de 3 ans (Jo pour Jeune) et de 8 ans (Vi pour Vieille) de l’Altiplano central de Bolivie. Deux qualités de paille ont été utilisées: la paille appelée N- avec N = 0.3 % et N+ avec N = 1.6 %. Les analyses de 14C, C, 15N et N totaux du sol prélevé à neuf dates sont utilisées dans un modèle de décomposition à 5 compartiments: MOMOS-6. Le taux de N de chaque compartiment est calculé d’après son taux de C divisé par son rapport C/N. L’entrée du modèle, la nécromasse (NC), est divisée en matériel végétal labile (VL) et stable (VS), sur la base de ses caractères biochimiques. Ses C et N sont utilisés par la biomasse microbienne (BM) qui alimente les compartiments d’humus labile (HL) et stable (HS). Les sorties sont le CO2 produit par BM et N minéral. Chaque constante de vitesse est multipliée par une fonction de réponse à la température et à l’humidité du sol. Le modèle SAHEL prédit cette humidité. Pour les 2 qualités de paille, après 2 ans d’incubation, il ne subsistait plus que 1/4 du’14C initial. Avec N+, 60 à 70 % et avec N-, 55 % du 14C initial ont été minéralisés durant les 100 premiers jours. Après 2 ans, il subsistait 45 et 60 % du 15N initial pour N+ et N- respectivement. L’âge de la jachère a un effet nettement moins important que la teneur en N de la paille qui freine la minéralisation de 15N mais aussi la minéralisation initiale de 14C. La biomasse microbienne prédite par le modèle atteint initialement 30 et 13 % du 14C et 75 et 50 % du 15N qui subsistent dans le sol pour les pailles N+ et N respectivement. Durant les premiers 100 jours, ces valeurs descendent à moins de 8 % pour 14C et entre 10 et 12 % pour 15N pour les deux types de pailles. Durant cette phase initiale, 55 à 65 % du 14C et 65 à 75 % du 15N qui subsistent, s’accumulent progressivement dans l’humus labile qui devient la source principale de C et N pour les microorganismes et d’azote minéral pour les plantes. L’humus stable ne dépasse jamais 2 % du 14C restant et 2.5 % du 15N restant. Le quotient métabolique de la biomasse microbienne (q CO2) était plus élevé avec N- qu’avec N+, du fait d’une demande plus importante en énergie de maintenance pour N-.
Mots clés: Altiplano, Matière organique, Modélisation, 14C, 15N.
Introducción
El Altiplano central de Bolivia se caracteriza por una agricultura de secano con descanso largo, donde alternan un año de cultivo de papa y hasta dos años de cereal o quinua con 510 años de descanso pastoreado, dominado por plantas anuales, perennes y finalmente una vegetación arbustiva característica de la puna, que no cubre más del 40% de la superficie del suelo. El clima es seco con 400 mm de precipitación promedio, distribuida en 3-4 meses y una temperatura media anual de 8.7°C (con fuerte amplitud térmica diaria). El Altiplano seco se extiende del Norte de Chile hasta Bolivia. Bajo estas condiciones climáticas, se desarrolla poco el suelo y su contenido en materia orgánica es muy bajo. Contiene generalmente menos de 0.7% de C orgánico, con C/N muy bajo y el pH es neutro a ligeramente ácido (Hervé 1994). En estos sistemas de cultivo con descanso, que usan escasos entrantes, el objetivo de este trabajo consistía en entender el papel de la materia orgánica como stock de nutrientes (en particular del nitrógeno). Esta preocupación era justificada por tres cuestiones: (1) Estos sistemas se caracterizan por una disminución rápida de la fertilidad del suelo (después generalmente de tres años de cultivo) y por la necesidad de un descanso largo para reconstituir la fertilidad. Sin contribución mineral, el ciclo de N en estos sistemas es administrado exclusivamente por los flujos entre el cultivo, el suelo, la vegetación natural que se desarrolla durante el descanso y el estiércol de los animales. El objetivo consiste en cuantificar estos flujos integrando también el carbono que es el soporte de la energía en la dinámica de la materia orgánica. (2) La comprensión del funcionamiento de los suelos pobres en materia orgánica, en particular bajo condiciones secas, tropieza con un problema que no ha sido aun aclarado: tienen generalmente valores de C/N excepcionalmente bajos de 5-9, comparados a los valores de 10 a 12 en los suelos a mull activos de las condiciones templadas. (3) Por razones geológicas se desarrolla una parte de los suelos del Altiplano central de Bolivia sobre material esencialmente arenoso. En el horizonte 0-20 cm, la ausencia de arcillas hace que la materia orgánica no sea protegida de la biodegradación y tenga una evolución rápida. Esto explica en parte el escaso contenido orgánico de estos suelos. El mismo trabajo se realizó también en descansos del páramo de los Andes de Mérida en Venezuela, bajo condiciones tropicales de alta montaña más húmedas, en suelos ácidos desarrollados sobre esquistos, de textura más fina y que presentan una fuerte acumulación de materia orgánica. Ambas experiencias rigurosamente idénticas permiten examinar el papel de las condiciones físico-químico del suelo sobre la dinámica de la materia orgánica. En ambos lugares, venezolano y boliviano, el sistema de cultivo dominante es de papa después de varios años de descanso del suelo.
En este trabajo, se ha incubado un material vegetal normal de paja de trigo maduro, marcado con 14C y 15N en un joven y un viejo descanso. Se utilizaron dos tipos de paja sobre la base del contenido en N: una rica (C/N = 26), la otra pobre (C/N = 130) en N. El experimento se llevó simultáneamente durante dos años en la puna seca del Altiplano central de Bolivia y en el páramo húmedo de Venezuela. Los objetivos eran los siguientes: (1) Modelar los flujos de C y N a través de los compartimentos de la materia orgánica del suelo, (2) Evaluar el papel de la edad del descanso sobre los procesos orgánicos, (3) Definir el papel de la riqueza en N del material vegetal, y finalmente (4) Comparar los dos tipos de suelo en descanso, muy diferentes en el páramo y la puna, para determinar el papel del ambiente físico-químico del suelo.
Material y métodos
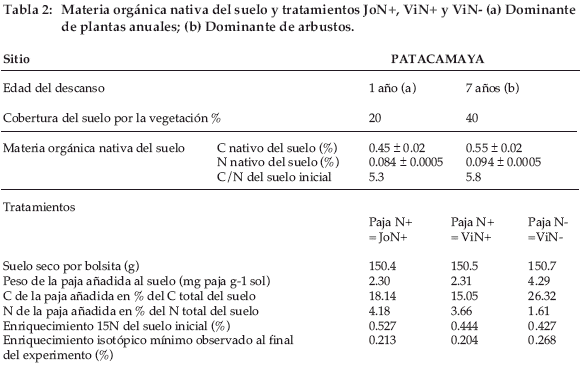
]]> Área de estudioEl experimento se realizó durante dos años (1998-2000) en la estación experimental de Patacamaya (17.15 S, 67.57 W, altitud 3.789 m) del Instituto Boliviano de Tecnología Agropastoral (IBTA) situada en el Altiplano central de Bolivia. Durante estos dos años empezando en noviembre, la temperatura media fue de 8.6°C, la media de la mínima de 0.2°C y la media de la máxima de 17.2°C; la precipitación media fue de 438 mm. El suelo es poco evolucionado; el horizonte 0-20 cm es arenoso (arena = 78.8%; limo = 14.8%; arcilla = 6.4%) y sin estructura, el pHH20 = 6.34 (Hervé 1994), C orgánico total = 0.5 ± 0.02%, N total = 0.09 ± 0.005% (C/N = 5.5), la capacidad de campo = 0.35 (v/v) y el punto de marchitez = 0.06 (v/v). Simultáneamente el mismo experimento se realizó en Gavidia, en el páramo de Mérida en Venezuela, con un clima más húmedo (precipitación = 1.352 mm por año), una temperatura (media anual = 8.4°C) más elevada y más constante a través de las temporadas y un suelo muy orgánico con un C/N más elevado (C del horizonte 0-20 cm = 7.5%, N = 0.53%).
Introducción de los trazadores isotópicos
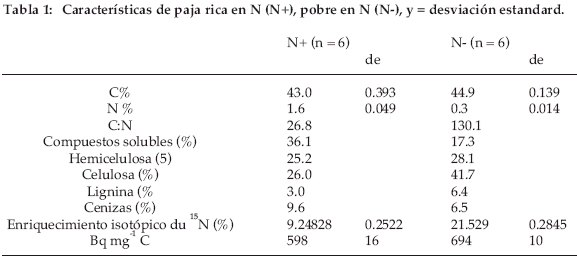
Las pajas están marcadas con 14C y 15N en una cámara de marcado isotópico desde el germen hasta la completa madurez. Se utilizaron dos tipos de paja: una paja rica en N (llamada paja N+) con N total = 1.6% y una paja pobre en N (llamada N-) con N = 0.3% (Tabla 1). Solamente se utilizaron los tallos y las hojas, que fueron machacadas groseramente con el fin de obtener partículas que pasan a través de un tamiz de 5 mm. Con el fin de obtener una señal medible de 15N hasta el final del experimento, el marcado en 15N de la paja N- era tres veces más elevado que para la paja N+ (enriquecimiento isotópico del 15N = 21.53% para N- y 9.25% para N+). La paja N- contenía también menos compuestos solubles y más celulosa y lignina (Tabla 1).
El experimento se instaló en un descanso de tres años (Jo por joven) con la paja N+ (= tratamiento JoN+) solamente y en un descanso de ocho años ( Vi por Viejo) con la paja N+ (= tratamiento ViN+) y N- (= tratamiento ViN-). Las muestras de suelo tomadas en el horizonte 5-10 cm, secadas al aire y homogenizadas, se colocaron en bolsas en poliéster de 8-10 cm. La cara superior de estas bolsas estaba constituida por mallas de 1 mm para permitir la penetración de las raíces, la cara inferior era con mallas de 0.25 mm con el fin de evitar la pérdida de suelo. Cada bolsa contenía 150 g de suelo (Tabla 2). Para los tratamientos JoN+ y ViN+, 347 mg de paja N+ se añadieron a cada bolsa y para el tratamiento ViN-, 646 mg de paja N-. El enriquecimiento isotópico inicial en 15N era siempre superior al 0.5% y los valores al final del experimento eran aún superiores a 0.2%. El ratio isotópico natural era de 0.3729±0.002 (n=4x2) para Patacamaya y 0.3713±0.0001 (n=4x2) para Gavidia. El enriquecimiento isotópico seguía siendo suficiente para ser medido en espectrometría de masa. El carbono de la paja añadida representaba un 15% (JoN+ et ViN+) y un 26% (ViN-) del C total de la mezcla suelo-paja y el nitrógeno total añadido era 1.6 y un 4.2% de la mezcla, respectivamente. La paja se añadió individualmente a cada bolsa y se homogenizó con el suelo. Las bolsas se instalaron en el suelo, a 5-10 cm de profundidad y a una distancia de 20 cm entre ellas. Fueron cubiertas por 5 cm de suelo de la capa 0-5 cm. Puesto que el suelo instalado en las bolsas se había secado previamente al aire, al final de la instalación cada bolsa enterrada fue regada lentamente con una cantidad de agua que correspondía a la capacidad al campo de la capa 0-10 cm. El experimento se instaló el 28/ 11/1998 y duró dos años. Durante este periodo, se efectuaron nueve muestreos para cada tratamiento con cuatro repeticiones por tratamiento. En los muestreos, las bolsas se colocaron individualmente en embalajes herméticos para el transporte. Luego su contenido fue pesado húmedo y secado al aire en el Instituto Boliviano de Técnicas Nucleares (IBTEN) de Viacha (Bolivia) y finalmente enviado al Cefe-CNRS de Montpellier (Francia) para su análisis.
Análisis
]]> Se homogenizó la muestra de suelo de cada repetición de terreno y se machacó una fracción alícuota (Retsch MM2). El C total y 14C fueron analizados, combinando un Carmograph 12A (Wösthoff Bochum, Alemania) y una espectrometría a centelleo líquida (Packard tricarb 1500) siguiendo a Bottner & Warembourg (1976). Se analizaron cuatro repeticiones para cada repetición de terreno, lo que representa 16 análisis para cada muestreo. Los N total y 15N fueron determinados, combinando un análisis CHN y una espectrometría de masa (Finnigan delta S) en el Centro de Análisis del CNRS de Solaize (Francia) con dos repeticiones por repetición de terreno o sea ocho análisis por muestreo. Se midieron 14C y 15N de la biomasa microbiana solamente en el experimento de Venezuela por fumigación-extracción (Joergensen 1996, Joergensen & Mueller 1996), espectrometría a centelleo líquida (ULA, Mérida, Venezuela) y espectrometría de masa (CNRS Solaize, Francia). El fraccionamiento bioquímico del material vegetal se realizó siguiendo el método Van Soest et al. (1991).
Modelización
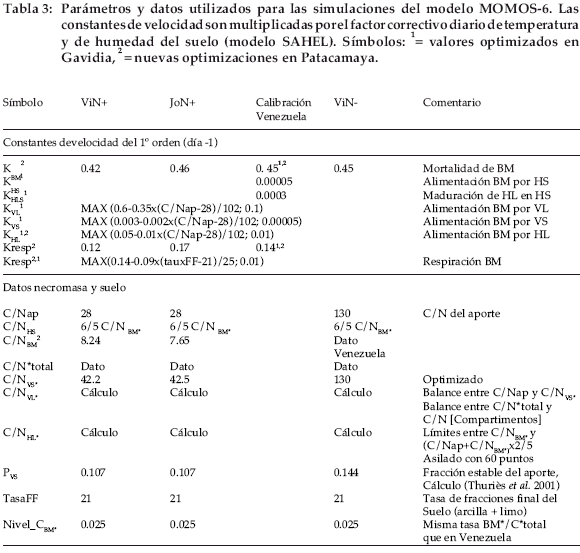
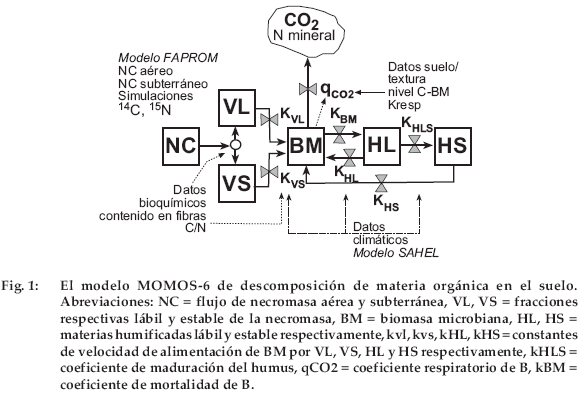
El modelo Momos-6 está constituido por cinco compartimientos como en la versión inicial de Pansu et al. (1998), pero con un diagrama de flujo muy diferente (Fig. 1, tabla 3). La necromasa (NC) constituye la entrada de C y N del material vegetal. El flujo de NC se divide entre el material vegetal lábil (VL) y el estable (VS) sobre la base de las fórmulas propuestas por Thuriès et al. (2002), según la composición bioquímica del material vegetal (Van Soest et al. 1991). Sólo se utilizaron aquí las fórmulas por las que se define la fracción estable de NC (Thuriès et al. 2001). La entrada de VL y VS en la biomasa microbiana (BM) es controlada por dos constantes de velocidad de primer orden: kVL y kVS. La respiración microbiana, es decir, el CO2 que deja el compartimento de biomasa microbiana (BM), constituye la única vía de salida de C; la cual es controlada por el cociente respiratorio q CO2 de BM, que es dependiente de BM y de la textura del suelo. Las otras constantes de velocidad kHL y kHS controlan la alimentación de BM por los compartimientos humus lábil (HL) y humus estable (HS). Las constantes kVL, kVS y kHL se conectan a la tasa C/N de la contribución orgánica inicial. La constante kBM representa a la tasa de mortalidad de los microorganismos y kHLS es una constante de maduración que cuantifica la transformación lenta de una pequeña fracción de HL en HS. Una particularidad de Momos-6 es la de ser controlado solamente por constantes de velocidad (dimensión T-1), con exclusión de otros parámetros sin dimensión generalmente utilizados para dividir los flujos de materia, por ejemplo en los modelos propuestos por Parton et al. (1987), Bradbury et al. (1993) o Pansu et al. (1998).
Cada constante de velocidad (Tabla 3) está multiplicada por una función de respuesta a la temperatura f(è) y a la humedad del suelo f(μ) f(è) es una ley Q10 = 2 para una temperatura óptima de actividad microbiana de 20°C para estos suelos de montaña (Kätterer et al. 1998); f(μ) es una función linear de la humedad actual del suelo con relación a la capacidad de campo. La humedad actual para el horizonte 5-10 cm fue calculada por el modelo SAHEL (Penning de Vries et al. 1989); f(è) y f(μ) varían de 0 a 1.
El 15N de cada compartimiento se calcula según su 14C, dividido por su tasa 14C/15N. La estimación de las tasas 14C/15N resulta entonces muy importante. Proviene de: (1) Medidas directas para 14C/15N[NC], 14C/15N[BM] y 14C/ 15N [total] del suelo, (2) Una optimización en base a los datos medidos para 14C/15N[VS ] y 14C/15N[HS ], y (3) Cálculos de balances para 14C/15N[VL] (conexión con 14C/15N[NC] y 14C/ 15N[VS]) y 14C/15N[HL ] (conexión entre 14C/ 15N[total] y 14C/15N de los cuatro compartimientos restantes). Algunos de los valores obtenidos están alisados dentro de los límites para evitar irregularidades de predicciones. El nitrógeno mineral producido es calculado por diferencia entre el 15N total inicialmente aportado y la suma del 15N de los diferentes compartimientos orgánicos. En este trabajo realizado en Patacamaya no se midió la biomasa microbiana; los valores de14C/15N[BM] proceden del experimento conducido en Gavidia, Venezuela.
Resultados
Modelización
]]> Los parámetros del modelo se ajustaron con el experimento de Gavidia que implicaba también las medidas del compartimento BM (Fig. 1, tabla 3). Con los datos experimentales 14C y15N se estimaron los parámetros kVL, kVS, kBM, kHL y kresp. A partir de los únicos datos 14C y 15N de este experimento que solo ha durado dos años no fue posible ajustar con suficiente precisión los parámetros kHLS y kHS que regulan la entrada y salida del compartimento HS a evolución muy lenta. Así, el kHLS se ha ajustado con un C total medido en las mismas muestras que los trazadores; kHS se ha ajustado en la aplicación de este modelo a una estimación de la evolución de la materia orgánica de estos suelos a largo plazo. Aun siendo ajustado con el C total, este compartimiento podía ser integrado al modelo para mejorar las predicciones de 14C y 15N al final de la incubación (poco subestimadas en ausencia de HS, comparando con los valores medidos).El modelo pudo a continuación aplicarse al experimento de Patacamaya únicamente mediante reajuste del parámetro kresp que regulaba el coeficiente respiratorio (q CO2) de BM. Se propuso una relación lineal entre kresp y la textura del suelo. El modelo requiere también del conocimiento del valor medio del carbono de la biomasa microbiana del suelo (niveau_C[BM ], fig. 1, tabla 3). La influencia de la calidad de los dos tipos de paja marcada fue tomada en cuenta de dos maneras: (1) Cálculo de la fracción estable por las fórmulas de Thuriès et al. (2002) y (2) Propuesta de una relación lineal de corrección de las constantes de velocidad kVL, kVR, kHL en función de los C/ N respectivos de VL y VS de la paja aportada.
14C y 15N totales
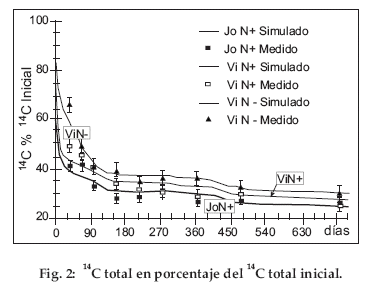
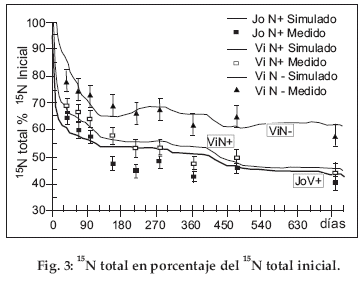
En las figuras 2 y 3, el 14C y el 15N que quedan en el suelo se expresan en un porcentaje de los 14C y 15N iniciales. Para los tres tratamientos, el 14C se mineraliza rápidamente, puesto que después de dos años ya no subsiste más que 1/4 del 14C inicial (Fig. 2).
Para ambos tratamientos con paja rica en N (JoN+ y ViN+) la mineralización de 14C es especialmente rápida al inicio, puesto que se mineraliza el 35-40% del 14C inicial en menos de 100 días. Esto genera en la curva de mineralización una forma característica en dos fases: una fase rápida que corresponde a la desaparición de los compuestos lábiles durante los primeros 100 días y una fase lenta que corresponde a la mineralización de los compuestos más estables. La misma forma de curva es aplicable al tratamiento con paja pobre en N (ViN-), sin embargo con una mineralización claramente más lenta durante la fase rápida inicial. El escaso contenido en N de la paja frena a la mineralización inicial de 14C. Para los tres tratamientos, las curvas muestran también una neta influencia de las condiciones climáticas entre el 28/11/1998 y el 27/11/2000. Por ejemplo, la mineralización es más activa entre los días 400 y 480 (enero, febrero y marzo de 2000) que entre los días 150 y 400 y que más allá de 480. Estas curvas en escalera reflejan la influencia de las temporadas húmedas que alternan con las temporadas secas. Esto no es visible para enero, febrero, marzo de 1999, puesto que este período húmedo corresponde también a la mineralización rápida inicial de los compuestos lábiles. La fuerte actividad inicial encubre al efecto de la temporada húmeda.
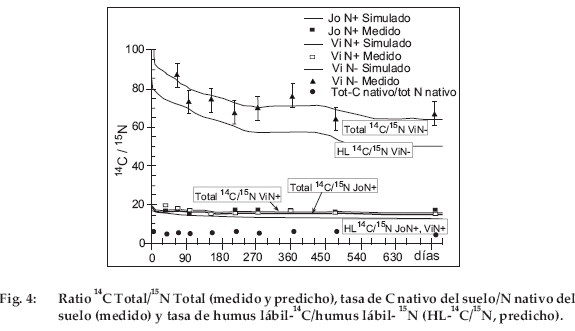
Los valores medidos muestran una mineralización ligeramente más rápida para JoN+ que para ViN+. Es generalmente significativa durante los 150 primeros días. La edad del descanso solo afecta débilmente a las velocidades de mineralización de14C. El aspecto de las curvas de mineralización de 15N en porcentaje del 15N inicial (Fig. 3) es bastante comparable a la de 14C en porcentaje del 14C inicial (Fig. 2), con una fase de mineralización inicial rápida y una fase más lenta más allá de 120 días y también con formas en escalera que reflejan el efecto del clima. Sin embargo, diferencias importantes aparecen entre 14C y 15N. Aunque la forma de las curvas sea comparable, 15N que queda en el suelo no desciende nunca por debajo de un 40% del 15N inicial contra un 25% para 14C. La diferencia entre JoN+ y ViN+ en los valores medidos es generalmente significativa durante el primer año. La diferencia entre ViN+ y ViN- es significativa durante todo el tiempo del experimento. Entonces para la mineralización de 15N, la edad del descanso y el contenido en N de la paja son discriminantes. El 14C/15N inicial de la paja N+ era 27 (Tab. 1); en JoN+ y ViN+ la tasa 14C/15N total del suelo disminuye en menos de 10 días a 17, luego a 16 después de 150 días (Fig 4). Más allá, su disminución es lenta puesto que es aún aproximadamente de 16 después de 750 días. Para los dos tratamientos con N+, la tasa de mineralización de 14C es pues claramente más importante que para 15N durante los 10 primeros días; pero más allá de este tiempo las tasas para 14C y 15N son idénticas. La paja pobre tiene un elevado 14C/15N de 130 (Tabla 1). Como para JoN+ y ViN+, en el tratamiento ViN- la tasa disminuye mucho durante los 10 primeros días cayendo de 130 a 90. Pero, a diferencia de los dos tratamientos anteriores y más allá de este tiempo, 14C/15N disminuye constantemente; alcanzando todavía 60 al final del experimento. El 14C/15N procedente de la paja N+ y N- se comporta de manera muy distinta.
]]> La figura 4 presenta también el (C nativo del sol)/(N nativo del suelo), es decir (C total -14C)/(N total -15N) medido al momento de los muestreos y se sitúa a lo largo del experimento entre 5 y 6. Eso significa que después de los dos años de incubación para los tres tratamientos, el 14C/ 15N dista mucho de haber alcanzado al C/N nativo del suelo. La diferencia es más importante para la paja N- que para la paja N+.
Biomasa microbiana, humus lábil y humus estable
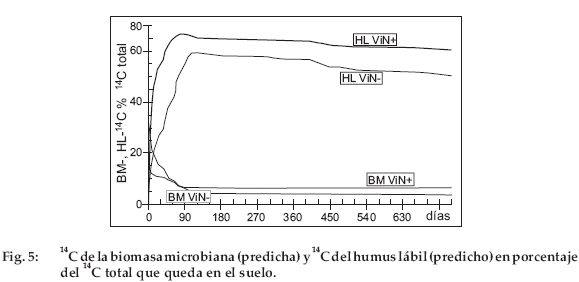
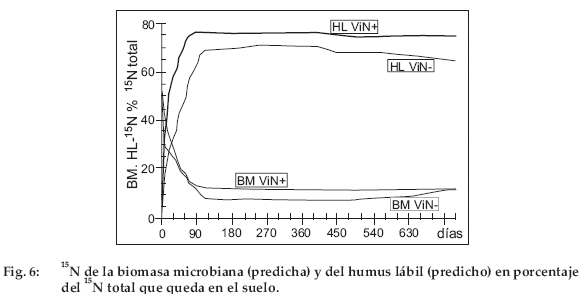
La desaparición del 14C y del 15N del suelo se debe a la mineralización en 14CO2 atmosférico y 15N mineral utilizado por las raíces. La pérdida por percolación es probablemente escasa porque las precipitaciones son escasas. Las figuras 5 para 14C y 6 para 15N ilustran las predicciones para BM y HL. Como las predicciones para JoN+ y ViN+ son muy cercanas, estas figuras presentan solamente a ViN+ y ViN-. Desde los primeros días, BM-14C alcanza un 30% para ViN+ y un 12% para ViNdel 14C total. Para BM-14C y -15N, así como para HL-14C y -15N, las curvas predichas presentan dos etapas muy distintas: una fase rápida de disminución para BM-14C y -15N y simultáneamente una fase de aumento rápido para HL-14C y -15N, luego una fase de escasa actividad, durante la cual HL-14C y -15N disminuyen lentamente y BM-14C y -15N evolucionan poco. El límite entre ambas fases tanto para BM como para HL se sitúa entre 80 y 100 días. El comportamiento de estos dos compartimientos es explicado por las fracciones lábiles (VL) y estables (VS) de la paja (no presentadas en las figuras). Según el modelo, la paja N+ contiene inicialmente 90% y la paja N86% de VL, el complemento siendo VS. Después de 80 a 100 días, la fracción VL desapareció completamente. Más allá de este tiempo, solamente la fracción VS del material vegetal queda en el suelo y se descompone muy lentamente (Tabla 3). Es entonces la calidad del material vegetal (materializada por la proporción de VL y VS) que explica la dinámica de BM y HL. Durante la fase activa, la caída rápida de la biomasa acompaña el agotamiento rápido de VL y simultáneamente a la acumulación rápida de HL. Luego durante la fase de escasa actividad, HL disminuye lentamente. Durante la fase de evolución lenta, por encima de 80-100 días, la actividad del suelo es debida esencialmente a la descomposición de la reserva de HL que se ha constituido a partir de VL.
Para 15N (Fig. 6) es VL durante el primer mes y luego HL que constituyen la fuente esencial de nitrógeno mineralizable. A principios de la fase de escasa actividad, HL-14C y HL-15N representan entre 55 y 65% del 14C total y entre 65 y 75% del 15N total. El humus estable (HS-14C et -15N) no se presentó en las figuras, pero aumenta de 0 a 2% del 14C total al final del experimento y de 0 a 2.5% del 15N total. Después de los dos años de experimento, este compartimento solo tiene una escasa fracción inmovilizada de 14C y 15N.
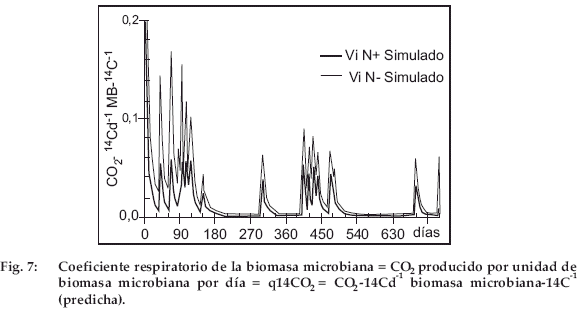
Cociente respiratorio de la biomasa microbiana (q14CO2)
La diferencia entre la evolución de la paja N+ y N- se ilustra por el cociente respiratorio de la biomasa microbiana (Fig. 7, la diferencia entre JoN+ y ViN+ es escasa, solo son presentadas ViN+ y ViN-), definida aquí por el 14CO2 producido por unidad de biomasa microbiana en un día (q14CO2 = 14CO2 día-1 MB-14C-1). Para la paja N- (ViN-), el 14C total disminuye menos rápidamente (Fig. 2) y BM-14C es más escaso (Fig. 5) que para la paja N+ (ViN+). Con N-, la respiración-14C y la biomasa microbiana-14C son más bajas que con ViN+. Sin embargo, la biomasa microbiana es más activa, puesto que en ViN- el qCO2 es más elevado que en ViN+ (Fig. 7). Esto significa que la biomasa gasta más energía para sobrevivir en este medio pobre en N. El comportamiento de la biomasa frente a 15N es más difícil de interpretar, puesto que una parte del 15N mineralizado es parcialmente reinmovilizada.
]]>
Efecto de la edad del descanso sobre las velocidades de descomposición
Al comparar el descanso de tres años (JoV +) y de ocho años (ViN+) todo indica que el potencial de mineralización de C no fue muy alterado por la duración del descanso, puesto que la mineralización del 14C se modifica poco. Para 15N, la mineralización es ligera pero significativamente más rápida en JoN+ que en ViN+, pero la diferencia es también escasa, puesto que 14C/15N no es significativamente diferente entre ambos tratamientos. El modelo no prevé tampoco diferencia significativa entre JoN+ y ViN+ para los otros parámetros examinados en este trabajo. Los datos de Gavidia (que se encuentran en preparación) han logrado el mismo resultado.
Como lo mostraron Hervé et al. (1994) para Patacamaya y Abadin et al. (2002) para Gavidia, en ambos sitios la variabilidad del C y N total y del C/N es muy elevada, pero no permite detectar una variación en función de la edad del descanso y si hay una variación vinculada a la edad, es inferior a la variabilidad. El presente trabajo puso de manifiesto que para una contribución idéntica de paja de calidad idéntica, la edad del descanso poco o no interfiere sobre los procesos (en caso de la mineralización de N). Estos suelos conservan pues la parte fundamental de su capacidad biológica de descomposición, que sean resultantes de un período de descanso de ocho años o de una fase de cultivo abandonada desde tres años.
Papel del contenido en N de la necromasa
Dos criterios discriminan a los dos tipos de paja:
(1) El C/N de la paja N- es 130, el de la paja N+ es 27; (2) Según las ecuaciones de Thuriès et al. (2002) y sobre la base de sus caracteres bioquímicos (Tabla 1) la paja N- contiene un 86% de los compuestos inestables mientras que la paja N+ contiene un 90%. En la diferenciación del efecto de ambos tipos de paja, el modelo asigna un papel claramente más importante al contenido en N que a la proporción de VL y VS en la paja. Las medidas ponen de manifiesto que con la paja N-, la mineralización de 14C y 15N totales se frena significativamente. En el modelo, las constantes de velocidad de mineralización de 14C (kVL, kVS, kHL) disminuyen linealmente en función del aumento de 14C/15N de la paja inicial (Tabla 3). Así el aumento del 14C/15N de la paja implica una disminución del 14C y 15N de la biomasa microbiana y del humus lábil y una desaparición más lenta de la necromasa (VL + VS). Finalmente, en presencia de la paja N-, el modelo predice una actividad de los organismos más elevada, revelada por q14CO2. El modelo explica entonces que la falta de N para los microorganismos frena la actividad de mineralización de la necromasa y de las materias humificadas. En el estado actual del modelo, la relación entre el 14C/15N y las constantes de velocidad se supone lineal, puesto que en el experimento solamente se estudiaron dos valores extremos de C/N. En un estudio bibliográfico, Fog (1988) había demostrado que el estímulo de la mineralización de la materia orgánica por N se debía esencialmente a una aceleración de la mineralización de compuestos lábiles (como la celulosa) y no de los compuestos estables (como la lignina). Esto se comprueba en este modelo: el efecto del14C/15N sobre kVS y kHS es desdeñable, comparado al efecto sobre kVL y kHL. Sin embargo, Fog avanza también otras explicaciones, como la competencia entre organismos por el N. El medio defectuoso en N implica un aumento del q CO2, es decir que se incrementa la actividad microbiana por unidad de biomasa. Se sugieren varias explicaciones como un acortamiento de la duración de vida con una alta mortalidad o la mineralización de compuestos húmificados complejos que requiere una mayor diversidad enzimática. Anderson & Domsch (1993) pusieron de relieve efectos comparables para microorganismos que se desarrollaban en medios de pH ácido desfavorable. Concluyen que bajo tales condiciones se desvía una parte de la energía normalmente atribuida al crecimiento hacia una más importante demanda para el mantenimiento. A propósito de la nutrición nitrogenada de las plantas, conviene sacar a la luz la importancia del humus lábil que controla la parte fundamental del N como fuente de N mineral. Es un stock que se constituye durante los 80-100 primeros días y luego disminuye lentamente. Su C/N es más elevado que el de la biomasa microbiana pero es menos elevado que el de la necromasa y del humus estable.
Ahora bien la separación entre el humus lábil y el humus estable es solamente predicha por el modelo, pero no se puede medir hasta ahora. Convendría en el futuro materializar las fracciones predichas por técnicas de fraccionamiento físicas o químicas con el fin de acercar lo mejor posible medidas y predicciones conceptuales.
]]>La maduración de la materia orgánica
Después de dos años de incubación y según las predicciones del modelo, la proporción de 14C y 15N que está en el humus estable HS no sobrepasa al 2 y 2.5% de 14C y 15N total, respectivamente (HS no presentado en las figuras). La parte fundamental del material marcado humificado se encuentra en el humus lábil. Si se aplica el modelo Momos-6 al C y N nativo del suelo (C, N nativo del suelo = C, N total menos 14C, 15N), el modelo prevé una distribución muy diferente: más del 50% de C y N se encuentra en el humus estable (trabajo en preparación). Eso significa que el humus que deriva del material marcado, incubado in situ, está muy lejos de haber alcanzado su maduración. Esto es ilustrado también por la diferencia que subsiste entre 14C total/15N total y C nativo/N nativo del suelo (Fig. 4). El paso de HL hacia HS (Fig. 1) es un fenómeno muy lento, aunque el contenido en arcilla y limo probablemente no protege mucho al material humificado lábil. No se tienen en la actualidad datos sobre la edad de la materia orgánica de estos suelos.
Papel de las condiciones físico-químicas del suelo
Dos tipos de características físico-químicas distinguen al suelo de Gavidia del de Patacamaya. (1) Los suelos de Patacamaya son ligeramente ácidos (pH = 6.1); los de Gavidia son más ácidos (pH = 4.5) y en altura, una fuerte acidez se acompaña generalmente con la presencia de aluminio libre procedente de los productos de alteración de los minerales primarios o secundarios (Abadin et al. 2002). Así en Gavidia, la fuerte acidez del medio y la presencia tóxica del Al libre probablemente frenan a la actividad biológica (Gonzales-Prieto et al.1996); (2) Los suelos de Gavidia son limosos y estructurados, mientras que en Patacamaya son arenosos y de partículas. La textura arenosa de los suelos de Patacamaya favorece a la mineralización, porque facilita la circulación del oxígeno. Con bajos contenidos en arcilla y limo, la materia orgánica es poco protegida de la biodegradación y entonces es más accesible a los microorganismos. El clima diferencia también a estas dos situaciones. En Patacamaya las condiciones más secas y frías frenan a la mineralización durante algunos periodos del año (Fig.2). A pesar de ello, la mineralización de 14C y 15N total es más rápida que en Gavidia, donde probablemente son las condiciones de acidez, Al libre y granulometría las que frenan la descomposición. El modelo describe la influencia de las condiciones físico-químicas por una relación lineal negativa entre kresp (la respiración de la biomasa microbiana) y la tasa de limo + arcilla. Fue puesto de manifiesto varias veces experimentalmente, por ejemplo por Ladd et al. (1995) y Saggar et al. (1999), que estas partículas finas tomadas en su conjunto frenan la mineralización. Como la mayoría de los modelos publicados, Momos-6 solo tiene en cuenta al efecto de la granulometría (arcilla + limo). Este efecto ejerce específicamente aquí sobre la constante de respiración kresp, en vez de ejercer una acción global (Jenkinson 1990) o gradual (Molina 1998) sobre las constantes de velocidad de todos los compartimentos o solamente de los estables (Parton et al. 1987). Por otra parte, Saggar et al. (1996) han demostrado que el tipo de arcilla, expresado en superficie específica, controlaba estrechamente al tiempo de residencia del carbono. Pero es actualmente difícil el integrar este factor, así como otros factores importantes como pH, Al libre o la hidromorfía. Sucede lo mismo con el efecto de la temperatura. En este trabajo, la respuesta a la temperatura fue definida por una relación, donde Q10 = 2. Esto, que a menudo está admitido en los modelos, es arbitrario. En efecto, Dalias et al. (2002) han demostrado que el valor de Q10 puede variar con los tipos de suelo y en un mismo suelo con los tipos de mecanismos biológicos en juego (mineralización de la necromasa o compuestos humificados).
Conclusión
Este trabajo se llevó a cabo en un medio extremo de puna seca, relativamente poco explorado del Altiplano central de Bolivia. La productividad es baja tanto durante la fase de cultivo como durante el descanso, con escasos entrantes de residuos de cultivo, de necromasa natural o de estiércol animal. Por lo tanto, el suelo es muy pobre en materia orgánica. Además, en una gran parte de esta región, el suelo esencialmente arenoso es poco propicio para la retención del agua. Con el uso de trazadores y gracias a la modelización, este trabajo tenía por objeto examinar los flujos de C y N en la materia orgánica. A pesar de una larga temporada seca, la mineralización de C y N es rápida, con 75% y un 55% de C y N iniciales mineralizados, después de dos años de incubación con una necromasa de C/N bajo. Esta rápida descomposición se debe sobre todo a la textura arenosa del suelo, es decir, a la ausencia de protección física. Una fuerte actividad del suelo tiene lugar solamente durante los tres meses húmedos del año. El modelo divide a la necromasa en una fracción lábil y una estable y esto explica en parte la dinámica de descomposición: una fase rápida seguida de una fase lenta. El contenido en N de la necromasa es otro parámetro importante; que controla la mineralización no sólo del N total sino también del C total. El modelo explica este control de una manera relativamente simple: por una modificación de la actividad mineralizadora de los microorganismos sobre las fracciones de la materia orgánica. En la gestión de la mineralización de N y en consecuencia de la nutrición nitrogenada de las plantas, el modelo asigna un papel importante al humus lábil, por el tamaño del compartimento y por su C/N relativamente bajo. En una próxima etapa de este trabajo, una buena valoración cuantitativa y cualitativa de los residuos de cultivo y de la necromasa natural, debería permitir una predicción a largo plazo del funcionamiento de estos sistemas de alternancia cultivo/descanso.
]]> Agradecimientos
La comisión de la Unión Europea financió este trabajo incluido en el programa TROPANDES (contrato INCO-DC XII: ERBIC18CT98-0263). Los autores agradecen a la Dra. Tarsy Carballas quién garantizó eficazmente la coordinación del proyecto Tropandes, al Dr. Stephan Beck del Instituto de Ecologia del Universidad Mayor de San Andrés de La Paz (Bolivia) y al Centro IRD de La Paz por haber facilitado las condiciones materiales de realización del trabajo. Los autores agradecen también a Bruno Buatois (Cefe-CNRS de Montpellier) y Md Peretti (SCA-CNRS Solaize) por haber garantizado los análisis de 14C y 15N.
Referencias
Abadin, J., S.J. Gonzales-Prieto, L. Sarmiento, M.C. Villar & T. Carballas. 2002. Successional dynamics of soil characteristics in a long fallow agricultural system of the high tropical Andes. Soil Biology & Biochemistry 34: 1739-1748.
Anderson, T. H. & K. H. Domsch. 1993. The metabolic quotient for CO2 (qCO2) as a specific activity parameter to assess the effects of environment conditions, such as pH, on the microbial biomass of forest soils. Soil Biology & Biochemistry 25: 393-395.
Bottner, P. & F. Warembourg. 1976. Method for simultaneous measurement of total and radioactive carbon in soils, soil extracts and plant materials. Plant and Soil 45: 273-277.
Bradbury, N.J., A.P. Whitmore, P.B.S. Hart & D.S. Jenkinson. 1993. Modelling the fate of nitrogen in crop and soil in the years following application of 15N labelled fertiliser to winter wheat. Journal of Agricultural Science 121: 363-379.
Dalias, P., J.M. Anderson., P. Bottner & M.M. Coûteaux. 2002. Temperature responses of net nitrogen mineralization and nitrification in conifer forest soils incubated under standard laboratory conditions. Soil Biology & Biochemistry 34: 691-701.
Fog, K. 1988. The effect of added nitrogen on the rate of decomposition of organic matter. Biol. Rev. 63: 433-462.
[ Links ]Gonzales-Prieto, S. J., A. Cabaniero, M.C. Villar, M. Carballas & T. Carballas. 1996. Effect of soil characteristics on N mineralization capacity in 112 native and agricultural soils. Biol. Fertil. Soils 22: 252-260.
Hervé, D. 1994. Respuestas de los componentes de la fertilidad del suelo a la duracion del descanso. Pp. 155-170. En: Hervé, D., Genin D. & G. Rivière (eds.) Dinámicas del Descanso de la Tierra en los Andes, Ed., Orstom, Paris-La Paz.
[ Links ]Jenkinson, D. S. 1990. The turnover of organic carbon and nitrogen in soil. Phil. Trans.R. Soc. Lond. B. 329: 361-368.
[ Links ]Joergensen, R. G. 1996. The fumigationextraction method to estimate soil microbial biomass: calibration of the kEC value. Soil Biology & Biochemistry 28: 25-31.
[ Links ]Joergensen, R. G. & T. T. Mueller. 1996. The fumigation-extraction method to estimate soil microbial biomass: calibration of the kEN value. Soil Biology & Biochemistry 28: 33-37.
Kätterer, T., M. Reichstein, O. Andrén. & A. Lomander. 1998. Temperature dependence of organic matter decomposition: a critical review using literature data analysed with different models. Biol. Fertil. Soils 27: 258-262.
Ladd, J.N., M. Amato, P.R. Grace & J.A. van Veen. 1995. Simulation of C-14 turnover through the microbial biomass in soils incubated with C-14-labelled plant residues. Soil Biology & Biochemistry 27: 777-783.
Molina, J.-A.E., B. Nicolardot & H.H. Cheng. 1998. Influence of clay content and time on soil organic matter turnover and stabilization/Influence de la teneur en argile et du temps sur le taux de renouvellement et de stabilisation de la matière organique du sol. En: IUSS (ed.), 16th World Congress of Soil Science, Montpellier, http://www.iuss.org, symposium 7, scientific registration 1939, 8 p.
Pansu, M., Z. Sallih & P. Bottner. 1998. Modelling of soil nitrogen forms after organic amendments under controlled conditions. Soil Biology & Biochemistry 30: 19-29.
Parton, W.J., D.S. Schimel, C.V. Cole & D.S. Ojima. 1987. Analysis of factors controlling soil organic matter levels in Great Plains grasslands. Soil Science Society of America Journal 51: 1173-1179.
]]>Penning de Vries, F. W. T., D.M. Jansen & H. F. M. ten Berge. 1989. Simulation of ecophysiological processes of growth in several annual crops. Simulation monographs29, Wageningen Pudoc. 271 p.
[ Links ]Saggar, S., A. Parshotam, G.P. Sparling, C.W. Feltham & P.B.S. Hart. 1996. 14C-labelled ryegrass turnover and residence times in soils varying in clay content and mineralogy. Soil Biology & Biochemistry 28: 1677-1686.
Saggar, S., A. Parshotam, C. Hedley & G. Salt. 1999. 14C- labelled glucose turnover in New Zealand soils. Soil Biology & Biochemistry 31: 2025-2037.
Thuriès, L., M. Pansu, C. Feller, P. Hermann & J.C. Rémy. 2001. Kinetics of added organic matter decomposition in a mediterranean sandy soil. Soil Biology & Biochemistry 33: 997-1010.
Thuriès, L., M. Pansu, M.C. Larre-Larrouy & C. Feller. 2002. Biochemical composition and mineralisation kinetics of organic inputs in a sandy soil. Soil Biology & Biochemistry 34: 239-250.
Van Soest, P.J., J.B. Robertson & B.A. Lewis. 1991. Symposium: Carbohydrate methodology, metabolism, and nutritional implications in dairy cattle. Journal of Dairy Science 74: 3583-3597.
]]>