Descomposición de hojarasca y raíces en un sistema de descanso largo (Altiplano de Bolivia)
Decomposition of plant litter and roots in a long fallow system (Bolivian Altiplano)
Marie-Madeleine Coûteaux1, Dominique Hervé2 & Stephan Beck3
1Centre d’Ecologie Fonctionnelle et Evolutive, CNRS, 1919 route de Mende, 34293 Montpellier Cedex 5, Francia, marie-madeleine.couteaux@cefe.cnrs.fr
2Institut de Recherche pour le Développement, IRD, BP 64501, 34394 Montpellier Cedex 5, Francia, herve@mpl.ird.fr ]]>
Resumen
El sistema de cultivo con descanso se basa sobre el principio de la pérdida de materia orgánica del suelo durante el cultivo y su recuperación posterior al dejar las parcelas sin cultivar durante un período de descanso más o menos largo. Durante este período, el stock de materia orgánica del suelo se reconstituirá con la incorporación de restos de la vegetación natural sucesional. En un ensayo experimental en la puna semiárida de Bolivia (Patacamaya, S La Paz, 3.800 m), se estudió la descomposición de 12 tipos de material vegetal (diferentes especies y órganos) en un terreno en descanso desde hace tres años y otro desde hace ocho años, bajo condiciones naturales, incubando bolsas con material vegetal (litterbags) durante uno o dos años. Un análisis en componentes principales ordenó la calidad de hojarasca inicial en tres grupos: (i) partes aéreas de gramíneas, (ii) hojas y tallos de arbustos y de dicotiledóneas herbáceas, (iii) raíces.
El contenido de lignina fue superior en las plantas del terreno en descanso largo. Concluimos que las reservas de materia orgánica del suelo (MOS) se reconstituyen debido fundamentalmente a los aportes de sistemas radiculares y gramíneas, que son los materiales que presentaron la menor tasa de descomposición. Las hojas de los arbustos de “tola” (Baccharis incarum) perdieron un 60% de su masa en los primeros tres meses. Esta alta tasa de descomposición se explica principalmente por un alto contenido de componentes no estructurales (NS). Se proponen modelos para predecir la tasa de descomposición y la pérdida de peso durante el primer año.
Palabras clave: Sistema de descanso, Materia orgánica del suelo, Calidad de la hojarasca, Descomposición.
Abstract
The fallow agriculture system is based on the fact that after depletion of soil organic matter due to cultivation, the soil can recover its initial level of fertility by leaving the fields for a long fallow period. During this period, the soil organic matter stock will be reconstituted by the inputs of the residues of the natural vegetation succession. The decomposition of 12 kinds of plant material (different organs and species) from a young three-years old and an old eight-years old fallow was studied in the semiarid Bolivian puna (Patacamaya, South of La Paz, 3,800 m asl.) by incubating litterbags over one or two years in field conditions. A PCA ordinated the initial litter quality in three groups: (i) the grass shoots, (ii) the leaves and stems of non-grass plants, and (iii) the roots.
The lignin content was higher in the plants of the old fallow. The plant materials that decomposed the slowest were the grasses and the roots, and it is assumed that these plant materials are the main source to recover the SOM (Soil organic matter) lost during the cultivation period. Mass loss of leaves of shrubs “tola” (Baccharis incarum) reached about 60% during the first three months. This high decomposition rate was mainly due to non-structural compound content (NS). Models were proposed for predicting decomposition rate and first year mass loss.
]]> Keywords: Fallow system, Soil organic matter, Litter quality, Decomposition.
Introducción
La agricultura tradicional de secano en los Andes centrales se basa en un sistema de cultivo que alterna descansos largos con pocos años de cultivo. Durante el período de descanso, se supone que el suelo recupera su fertilidad natural, pero los mecanismos de tal recuperación no son bien comprendidos. En el Altiplano de Bolivia, el período de descanso solía durar más de diez años, pero el incremento de la población y la presión por una mayor cosecha tienden a modificar las estrategias de manejo, extendiendo la superficie cultivada y reduciendo la duración del descanso. Sin embargo, el descanso del suelo no puede ser reducido, a menos que una función esencial – la recuperación de la fertilidad del suelo pueda ser substituida por prácticas alternativas. La tendencia general para compensar la pérdida de fertilidad debido a la reducción de la duración del descanso, es el uso de diferentes técnicas de fertilización que buscan mantener o restablecer el balance de nutrientes del suelo en los campos (Wezel & Haigis 2002).
La pérdida de fertilidad tras el cultivo es generalmente atribuida a la pérdida de materia orgánica (Weisbach et al. 2002). Durante el período de descanso, la restitución de esta materia orgánica proviene de los aportes de residuos del cultivo previo y de la vegetación nativa. La cantidad y calidad de los aportes de materia orgánica de diferentes tipos de vegetación controlan la estabilización de materia orgánica del suelo (MOS) bajo distintos sistemas de cultivo. La tasa de descomposición y liberación de nutrientes y su sincronía con la demanda por parte de la vegetación son aún objeto de estudio (Tiessen et al. 2001).
Después del período de cultivo, la tierra es abandonada e invadida por vegetación natural sucesional. Durante el primer año, la especie dominante es una gramínea anual -Chondrosum simplex – que es rápidamente seguida por herbáceas dicotiledóneas, como las malezas Erodium cicutarium (Geraniaceae), Tarasa tenella (Malvaceae) y la nativa perenne, Oxalis bisfracta (Oxalidaceae). Después aparecen la leguminosa Lupinus otto-buchtienii y otras herbáceas dicotiledóneas, como las compuestas Schkuhria multiflora, Erigeron lanceolatus, algunos arbustos perennes de “tola” (Baccharis incarum y Parastrephia lepidophylla) y gramíneas perennes en matas densas de Stipa ichu, Festuca dolichophylla y matas abiertas de gramíneas de varias especies de Nassella. Todas estas plantas producen una amplia variedad de biomasa que después de la senescencia retorna al suelo como hojarasca.
El objetivo de este estudio fue determinar la tasa de descomposición de la hojarasca de las plantas más representativas de la sucesión de descanso, en función de su calidad química inicial, con el fin de apreciar su potencial en la estabilización de MOS. Nuestras hipótesis fueron que (i) el contenido de lignina de la hojarasca aumenta con la duración del período de descanso, (ii) la tasa de descomposición puede reducirse disminuyendo la duración del período de descanso y (iii) el secuestro potencial de MOS será mayor en la descomposición de las plantas leñosas que en las herbáceas.
Área de estudio
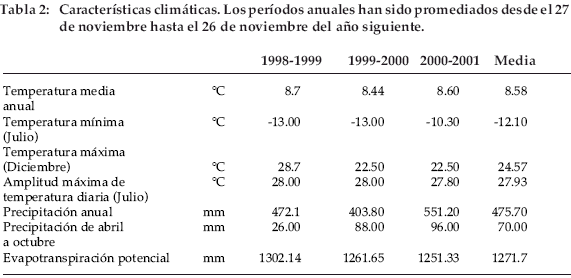
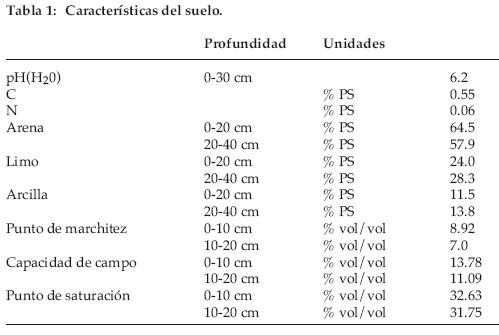
]]> El área de campo fue localizada en el Altiplano boliviano, en parcelas cercadas de la antigua Estación Experimental de Patacamaya, aproximadamente a 110 km al sur de La Paz y cerca del camino a Oruro (Long. 67°55’ W, Lat. 17°15 S, altitud: 3.800 m). El Altiplano con la vegetación de puna puede ser considerado como una estepa tropical alpina semidesértica, con cobertura vegetal limitada y una cobertura arbustiva cuya altura aumenta con la duración del período de descanso. El suelo es mayormente coluvial y sus características se resumen en la tabla 1. Se trata de un suelo arenoso con un bajo potencial de retención de agua, como lo indican la capacidad de campo y el punto de marchitez.El sitio está expuesto a condiciones climáticas diurnas típicas de la montaña alta tropical. Los parámetros de temperatura de los tres años fueron muy similares (Tabla 2) con 8.6°C de temperatura anual promedio, -12.1°C de temperatura diaria mínima en julio, 24.6°C de temperatura diaria máxima en diciembre y 28°C de amplitud diaria máxima de temperatura en julio. Las precipitaciones fueron bajas, con una estación seca de abril a octubre en el primer año y de marzo a octubre en el segundo. Las precipitaciones fueron levemente más altas durante el tercer año y con una época seca menos severa. Las condiciones relativas de extrema sequía se reflejan en la elevada evapotranspiración potencial, debida a la radiación global elevada en estas elevadas altitudes.
Material y métodos
Terminología
Las abreviaciones aplicadas en este capítulo se presentan en la tabla 3. En el lenguaje cotidiano de la zona, las parcelas que se dejan en descanso tras el cultivo se llaman descansos. En este trabajo respetaremos esta terminología y hablaremos en lo sucesivo de descanso corto para referirnos a las parcelas experimentales que llevan poco tiempo sin cultivarse y de descanso largo para referirnos a las parcelas que llevan mucho tiempo sin cultivarse.
]]> Selección del material vegetal
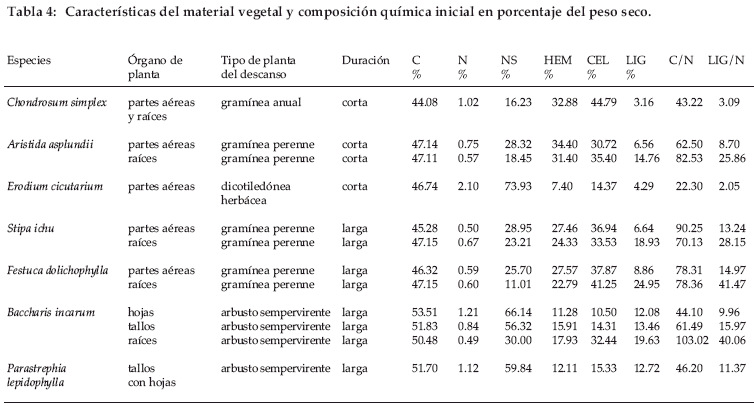
Siete especies de plantas, encuadrables en tres formas de vida – herbáceas dicotiledóneas, gramíneas y arbustos - fueron seleccionadas como representativas de descansos cortos a medianos y de medianos a antiguos. Chondrosum simplex, una gramínea anual pionera y Erodium cicutarium, una herbácea anual o bianual con raíz pivotante, representaron la vegetación de descansos cortos a medianos de aproximadamente cinco años, con una importante biomasa y cobertura de suelo. Como representativas del descanso mediano a largo, se seleccionaron Aristida asplundii, una herbácea nativa perenne y agresiva que se distingue por elevadas cobertura del suelo y biomasa porque no es pastoreada por el ganado, y matas densas perennes de Stipa ichu y Festuca dolichophylla, acompañadas hasta los siguientes estadios de descanso por Baccharis incarum y Parastrephia lepidophylla como especies arbustivas perennes.
Las hojas y tallos senescentes de las tres primeras especies fueron recolectados al final de la estación seca en octubre de 1999, en tanto que los de Stipa ichu y Festuca dolichophylla se recolectaron de la parte central muerta de las matas densas en noviembre de 1998. Para Baccharis incarum las hojas y tallos senescentes se recolectaron por separado. En cuanto a Parastrephia lepidophylla las hojas cilíndricas se confunden con los tallos: por ello, hojas y tallos fueron recolectados conjuntamente de la parte muerta de la planta (necromasa erecta) antes de noviembre de 1998. Las raíces fueron recolectadas de plantas arrancadas, sin distinguir necromasa de biomasa. El material obtenido fue secado al aire y luego guardado hasta su utilización. Doce tipos de material vegetal fueron estudiados (Tabla 4).
Bolsas de hojarasca
Cinco gramos de material vegetal secado al aire fueron introducidos en bolsas de 13 x 15 cm. Cuando fue necesario, el material fue previamente fragmentado en porciones de 2 cm. La red de la parte superior de la bolsa fue de 2 mm para permitir la entrada de mesofauna en las bolsas, y la red de la parte inferior fue de aproximadamente 0.1 mm para minimizar pérdidas durante la manipulación. Para cada tipo de material vegetal, 80 bolsas fueron dispuestas en 10 parcelas replicadas en el campo (2 x 2 m); en cada parcela se incubaron ocho bolsas para cada material vegetal, una para cada fecha de muestreo (10 réplicas, ocho fechas de muestreo). El material vegetal obtenido de partes aéreas (hojas, tallos) fue fijado a la superficie del suelo con clavos. Debido a la fuerte insolación que daño a las bolsas, una red de material resistente a la radiación fue instalada por encima de las bolsas en toda la superficie de cada parcela. El material de raíces fue enterrado a 10 cm de profundidad. El primer muestro se realizó un mes después de la instalación, posteriormente el intervalo de tiempo entre dos muestreos se incrementó progresivamente a medida que la incubación avanzaba.
Lugares y períodos de incubación
Las bolsas de hojarasca fueron incubadas en dos parcelas, un descanso corto y un descanso largo. Las incubaciones realizadas fueron las siguientes:
Análisis de la hojarasca en descomposición
En cada muestreo, se recolectaron 10 bolsas de hojarasca para cada material vegetal. En el Laboratorio de Calidad Ambiental (LCA) del Instituto de Ecología en La Paz, las muestras fueron limpiadas de contaminación de suelo o de raíces vivas y desecadas a temperatura ambiente hasta lograr un peso constante. Las bolsas de hojarasca fueron luego enviadas al CEFE-CNRS (Montpellier, Francia) por servicio de correo internacional (DHL). El contenido de las bolsas de hojarasca fue pesado para la determinación de la pérdida de masa. Debido a la contaminación mineral especialmente en bolsas enterradas, era importante corregir el peso para expresarlo libre de cenizas. La muestra fue luego molida (1 mm) usando una trituradora centrífuga (Cyclotec, Perstorp Analytical) y su contenido en ceniza determinado por la espectrofotometria NIR (NIRSystems 6500, Perstorp Analytical) (Coûteaux et al. 1998, McTiernan 1998, Kurz- Besson 2000). Se procedió a una selección de las muestras, sacando aquellas con espectros similares, identificadas por un análisis de componentes principales. En las muestras seleccionadas se analizó el contenido de cenizas tras seis horas a 650°C y el contenido de C mediante un analizador elemental CHN (Carbo Erba). Se construyeron ecuaciones usando regresiones múltiples entre la ceniza medida o el contenido en C y las longitudes de onda del espectro. El modelo se basó en 139 y 133 datos con una R2= 0.984 y 0.982 para ceniza y C, respectivamente. El cociente (error estándar de calibración/error estándar de la población de referencia) fue de 0.13 y 0.15, respectivamente, que son inferiores al valor umbral (0.2) utilizado para evaluar la bondad de los modelos. Estos modelos permitieron predecir el contenido de ceniza y de C para todas las muestras de hojarasca.
Calidad inicial de hojarasca
C y N fueron cuantificados con un analizador elemental CHN (Carbo Erba). Se aplicó el protocolo van Soest para la determinación del contenido de hemicelulosa, celulosa y lignina (van Soest & Robertson 1985) en el CIRAD en Montpellier, siguiendo la norma francesa AFNOR V18-122 y usando bolsas fibertec. Este método tiene mejor reproducibilidad que la técnica de filtración convencional (D’heer et al. 2000). Las muestras de suelo fueron primero tratadas con acetona para remover las resinas y taninos. Digestiones sucesivas con detergentes neutros y ácidos, así como ácido sulfúrico dan tres tipos de residuos de fibras: NDF, ADF y ADL, respectivamente. NDF es el residuo estructural que incluye hemicelulosa (HEM), celulosa (CEL) y lignina (LIG), siendo ADF la lignocelulosa y ADL el residuo final. NDF – ADF corresponde a la hemicelulosa junto a ciertos residuos de pared celular de la biomasa microbiana, ADF – ADL es la celulosa y el ADL es la verdadera lignina más los productos secundarios de la descomposición, cuya recalcitrancia es similar a la de la lignina. Los compuestos no estructurales (NS) son calculados como 100 – NDF y pueden ser mayormente considerados como compuestos solubles. Todas las concentraciones se expresan libres de cenizas.
]]> Estadísticas
Se realizó un análisis de componentes principales con StatBoxPro 5 (Grimmersoft, Francia), usando los tipos de material vegetal como individuos y sus composiciones químicas iniciales como variables. El tipo de órgano, el tipo de planta y el tiempo de descanso fueron tomados como variables adicionales. Los datos de masa remanente se ajustaron mediante Prism 3.00 (Graphpad software) a modelos exponenciales. El modelo exponencial simple se expresa por la ecuación:
![]()
siendo MR la masa remanente, en porcentaje respecto de la masa inicial, k la tasa de descomposición anual y t el tiempo en años. El modelo exponencial doble se expresa por la ecuación:
![]()
en que A es la fracción lábil de la materia orgánica, k1 la tasa anual de descomposición de esta fracción y k2 la tasa anual de descomposición de la fracción recalcitrante de la materia orgánica. Se asume que esta fracción recalcitrante es 100-A. Al igual que en la ecuación anterior, t es el tiempo en años.
Se estudiaron las relaciones entre la calidad inicial de hojarasca y las tasas de descomposición k y el primer año de pérdida de masa (PM1), mediante correlaciones de Pearson y el procedimiento de regresiones múltiples seriadas (Sigmastat, SPSS Inc.). Se aplicaron varias pruebas ANOVA de un factor para probar diferencias entre órganos de plantas o tipos de plantas. Todas las significaciones son registradas a un nivel del 5%, salvo que se indique de otra forma.
Resultados
Calidad inicial de hojarasca
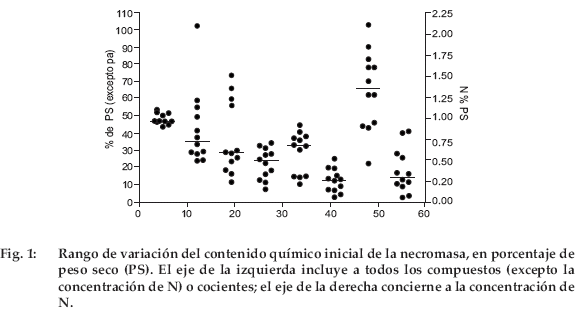
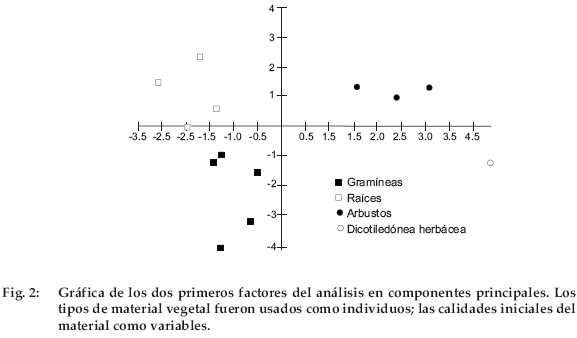
]]> Las concentraciones de C fluctuaron entre cerca de 44 hasta 52%, dándose los valores más altos en las especies arbustivas (Tabla 4, Fig.1).La concentración de N osciló entre 0.50 y 2.1%, siendo los valores más bajos en raíces y gramíneas, y el más elevado para Erodium cicutarium, muy superior al siguiente de 1.21% para las hojas de Baccharis incarum. Las concentraciones de NS fueron de 16-74%; los valores más bajos se dieron en gramíneas y el más alto, nuevamente para Erodium cicutarium. Se pueden distinguir dos grupos: (i) Erodium cicutarium, hojas y tallos de Baccharis incarum y tallos de Parastrephia lepidophylla con una concentración de NS más elevada que el 50% y (ii) el resto del material vegetal con una concentración de NS inferior al 50%. Las concentraciones de HEM fluctuaron entre 7 y 34%, el valor más bajo correspondió a Erodium cicutarium y el mayor para las gramíneas. Las concentraciones de CEL oscilaron entre 10 y 45%, los valores menores para hojas y tallos de especies arbustivas y para Erodium cicutarium, mientras que los mayores correspondieron a las gramíneas. Se observó una baja concentración de CEL (inferior al 20%) para Erodium cicutarium, hojas y tallos de Baccharis incarum y tallos de Parastrephia lepidophylla, mientras que la concentración de CEL fue mayor para las demás especies. Es notable observar que el grupo con baja concentración de CEL coincide con el grupo con alta concentración de NS. Las concentraciones de LIG fueron de 3-25%. Los valores C/N fluctuaron homogéneamente entre 22 y 103, y los valores LIG/N entre 3 y 41. Se realizó un análisis de componentes principales tomando los tipos de material vegetal como individuos y los componentes químicas de la hojarasca como variables. En una gráfica donde el primer y segundo factores explicaron el 88% de la varianza, el material vegetal quedó distribuido en tres grupos: hojas de arbustos y Erodium cicutarium, partes aéreas de gramíneas y las raíces de todas las plantas (Fig. 2). El primer eje se relaciona con los contenidos de N, NS, HEM y CEL, mientras que el segundo eje se relaciona con los contenidos en C y LIG. Se observó una correlación positiva y significativa entre N y NS (R=0.767) y negativa de N con HEM (R=0.642) y CEL (R=-0.633). La correlación entre N y LIG no fue significativa.
Contenido en cenizas
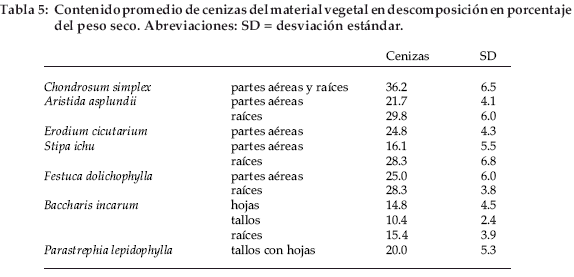
La determinación del contenido en cenizas es particularmente importante debido a la contaminación por partículas minerales. Las bolsas fijadas a la superficie del suelo sufrieron una contaminación de origen eólico, mientras que las bolsas enterradas fueron contaminadas por el suelo circundante. El contenido promedio de ceniza por material vegetal fluctuó entre 10 y 36% (Tabla 5).
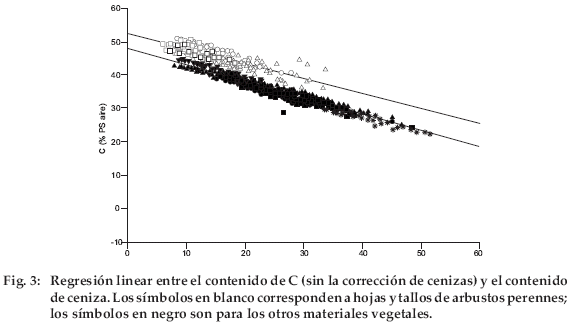
La gráfica del contenido de C (sin la corrección de cenizas) frente al contenido de cenizas (Fig. 3) muestra dos grupos bien diferenciados: un primer grupo, con hojas y tallos de Baccharis incarumy tallos deParastrephia lepidophylla, y un segundo grupo con todas las demás especies. El primer grupo fue identificado ya por su elevado contenido inicial de C (con la corrección de cenizas). Las pendientes de ambos grupos no son significativamente diferentes. El ajuste fue mejor para el segundo grupo (R2=0.95) que para el primero (R2=0.74).
]]>Cinética de la masa remanente (MR)
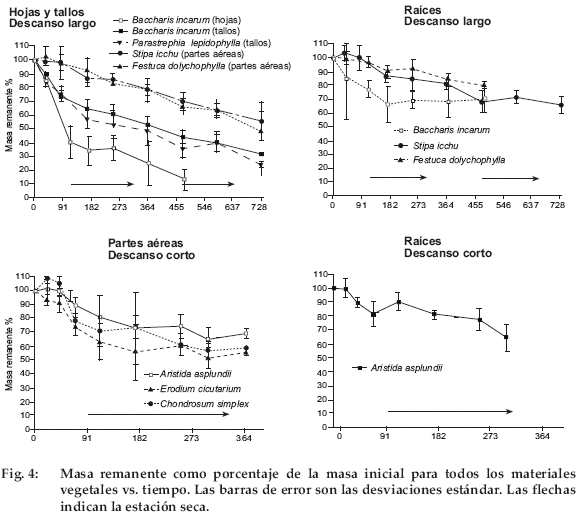
En descanso largo, la incubación duró unos dos años. Las hojas de Baccharis incarum fueron las más rápidas en descomponerse y perdieron el 60% de su masa durante los primeros tres meses (Fig. 4). La estación seca que siguió fue una fase de estabilización, seguida por una nueva fase de descomposición con el retorno de las lluvias. Después de 460 días de incubación, la masa remanente era sólo el 20% de la inicial. Los tallos de Baccharis incarum y Parastrephia lepidophyllase descompusieron más lentamente. Durante la primera época de lluvias, se perdió cerca del 40% de la masa inicial. Posteriormente, no se detectaron efectos estacionales. La masa remanente declinó linealmente hasta fines del segundo año. Stipa ichu y Festuca dolichophylla fueron las especies que se descompusieron más lentamente. Sus masas remanentes disminuyeron linealmente hasta el 60% después de dos años sin ningún aparente efecto estacional. Pese a que fueron enterradas, las raíces se descompusieron menos que la hojarasca por encima del suelo. Las raíces de Baccharis incarum fueron las más rápidas en descomponerse, con una pérdida de masa del 35% durante la primera estación lluviosa. Después, la masa remanente se estabilizó. Las raíces de Stipa ichu y Festuca dolichophylla mostraron un patrón similar al de sus partes aéreas.
En descanso corto, la incubación duró un año. Erodium cicutarium mostró la descomposición más rápida, con una pérdida de masa inicial en la época de lluvias del 40%. Las partes aéreas de gramíneas -Aristida asplundii y Chondrosum simplex – no sufrieron descomposición el primer mes, a pesar de encontrarse en la estación lluviosa. Durante la segunda fase de la estación lluviosa empezó una descomposición lenta, que se interrumpió para Aristida asplundii con la estación seca. En cambio, la descomposición de Chondrosum simplex continuó; al final de la incubación su masa remanente estaba aproximadamente al mismo nivel que la de Erodium cicutarium.
Regresiones no lineares
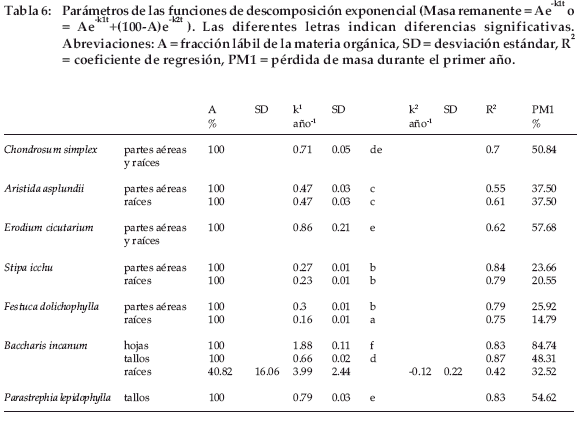
Se ajustó a los datos un modelo exponencial simple, excepto para raíces de Baccharis incarum para las cuales los parámetros no convergieron (Tabla 6). Para este material se obtuvo un ajuste adecuado mediante un modelo exponencial doble. Las tasas de descomposición fluctuaron entre 0.16-1.88 por año y en el primer año la pérdida de masa fue del 15% para raíces de Festuca dolichophylla hasta 84% para hojas de Baccharis incarum. Las tasas de descomposición de las raíces fueron significativamente inferiores a las de las partes aéreas (ANOVA de un factor). Las gramíneas mostraron las tasas más bajas de descomposición. No se observó ninguna influencia de la duración del descanso.
]]> Correlaciones y regresiones múltiples seriadas
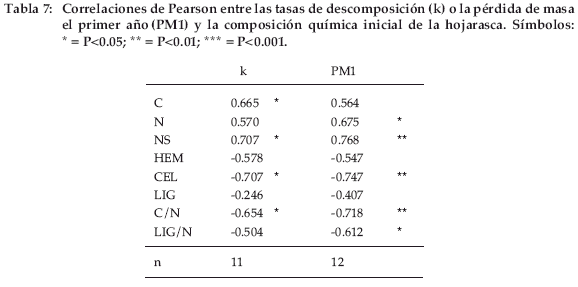
Las correlaciones entre las tasas de descomposición (k) y la composición química inicial de la hojarasca fueron positivas (P<0.05) para C y NS, mientras que fueron negativas para CEL y C/N (Tabla 7).
Las correlaciones con la pérdida de masa en el primer año (PM1) fueron positivas para N (P<0.05) y NS (P<0.01), y negativas para CEL (P<0.01), C/N (P<0.01) y LIG/N (P<0.05). Sin embargo, NS no es independiente de HEM y CEL, porque son los componentes dominantes expresados como proporciones de la masa total. Cuando NS es elevada HEM y CEL son bajas y viceversa. El porcentaje de LIG fue siempre bajo, y poco afectado por las variaciones de los otros componentes. En el análisis seriado de la regresión múltiple (Tabla 8), los componentes correlacionados no fueron nunca probados juntos.
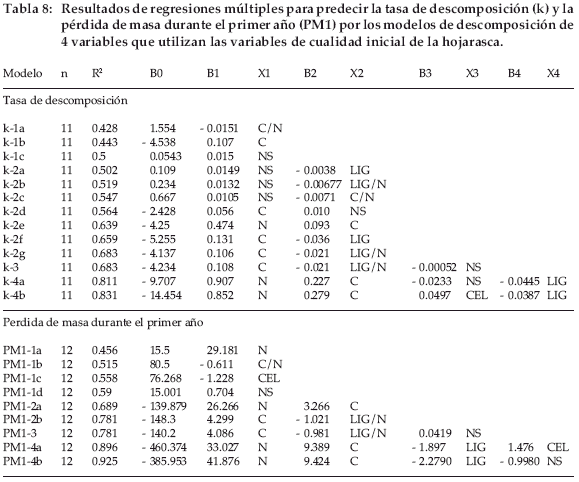
Los modelos que incluían a las mismas variables fueron generalmente mejores para PM1 que para k. Los mejores modelos con dos variables incluyeron a C y LIG/N para k y PM1. La mejora de los modelos con tres variables se debe a la adición de NS. Finalmente, se consiguió una otra mejora para estos modelos al añadir una cuarta variable:
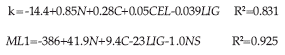
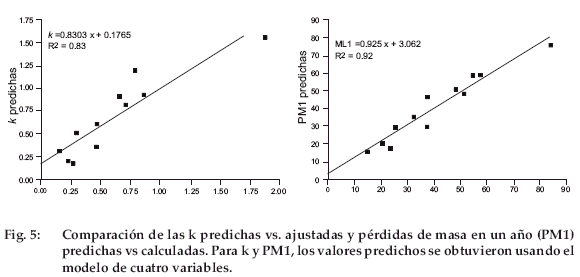
La figura 5 muestra los valores de k y PM1 predichos por los modelos de cuatro variables (k4b, PM1-4b) comparados con los mismos valores obtenidos mediante el modelo exponencial. Ambas pendientes fueron levemente inferiores a 1 con una intercepción positiva, indicando sobreestimaciones para valores bajos y subestimaciones para valores elevados. Al excluir el valor k de las hojas de Baccharis incarum, para el cual la predicción parece ser subestimada, la intercepción se trasladó a casi 0, pero la pendiente aumentó a 1.2, lo que indica una sobreestimación del modelo.
]]> Comparación entre tipos de material vegetal
Una ANOVA de un factor mostró que las tasas de descomposición k y la pérdida de masa durante el primer año (PM1) de las hojas y tallos de arbustos y deErodium cicutarium fueron mayores que las de gramíneas y raíces (Tabla 9). A pesar de una descomposición de raíces más lenta que la de gramíneas, las diferencias no fueron significativas.
Discusión
Efecto de la forma vital
La base de datos cubrió un amplio rango de variación de calidad de hojarasca, incluyendo arbustos perennes, una dicotiledónea herbácea y gramíneas con sus diferentes órganos: hojas o partes aéreas, tallos y raíces. Los arbustos y la dicotiledónea herbácea mostraron un elevado contenido en N y un bajo contenido en LIG, los partes aéreas de gramíneas un bajo contenido en NS y LIG, mientras que las raíces un elevado contenido en LIG. Los cocientes C/N y LIG/N – que a menudo son considerados buenos predictores de la tasa de descomposición (Aber & Melillo 1982, Pérez-Harguindeguy et al. 2000) – cubrieron un rango homogéneo de 22-103 y 3-41, respectivamente. Por el contrario, el rango de variación de N, NS y CEL fue discontinuo. La especie que se diferenció netamente de las demás fue Erodium cicutarium, con un muy alto contenido en N (2.1%) y que mostró la descomposición más rápida, después de las hojas de Baccharis incarum. Sin embargo, el contenido de N por sí solo no se correlacionó significativamente con la tasa de descomposición. NS y CEL diferenciaron a arbustos y a la dicotiledónea herbácea de las gramíneas y raíces. En este caso, la proporción C/N fue un mejor predictor que LIG/N. En un desierto de Norteamérica, Schaefer et al. (1985) también mostraron que LIG/N fue un mal predictor de la descomposición.
Algunos autores han mostrado que las diferencias en la calidad de hojarasca de las especies se relacionan con la forma vital. Como hemos mostrado en este trabajo, hay una relación entre forma vital y tasa de descomposición: las especies con una misma forma vital tienen una tasa de descomposición similar, en tanto que especies con formas vitales distintas tienen tasas de descomposición muy distintas. Hemos vistos que las hojas y tallos de arbustos y la dicotiledónea herbácea se descomponen más rápidamente que las partes aéreas de gramíneas. Esto contradice a Shaw & Harte (2001), quienes reportaron una tasa de reducción similar para las gramíneas y hojarasca de arbustos, así como a Hobbie (1996), quien observó en la tundra de Alaska una tasa de descomposición mayor para graminoides que para arbustos caducifolios. En Shaw & Harte (2001), la gramínea Festuca thurberi tenía un alto contenido en lignina (34%) y una alta propoción LIG/N (34), que explican la baja tasa de descomposición comparada con la observada por Hobbie (1996), que obtuvo para Carex bigelowii un contenido de lignina de 12.36% y de LIG/N de 20. También Trofymow et al. (2002) encontraron que la gramíneaFestuca halli se descompone más rápido que varias hojarascas de planifolios o de coníferas. Sin embargo, Cornelissen (1996), en un análisis sobre las tasas de descomposición de varias especies, mostró que las gramíneas se descomponen más lentamente que los arbustos caducifolios y más rápidamente que los arbustos perennes.
En nuestro estudio, los partes aéreas de gramíneas fueron los materiales más recalcitrantes, pese a su bajo contenido en lignina (3-9%) y su bajo cociente LIG/N (3-15). Por lo tanto, su recalcitrancia no puede ser atribuida a contenidos en lignina o N. Tal vez su baja proporción de compuestos lábiles como NS y sus consecuentes contenidos altos en HEM y CEL – que son comparativamente más recalcitrantes – pueden explicar su resistencia a la descomposición, superior a la de los otros materiales vegetales estudiados. La baja tasa de descomposición de Stipa ichu y Festuca dolichophylla puede ser atribuida a que el material vegetal fue obtenido de matas secas, que pueden haber pasado una larga etapa de senescencia. En tales condiciones, pueden haber sufrido importantes pérdidas de aminoácidos libres y carbohidratos solubles, por lixiviación (Marstop 1996). En cambio, Chondrosum simplex y Aristida asplundii fueron recolectados inmediatamente después de su senescencia. Este material vegetal queda verde muy poco tiempo y una parte se puede dispersar rápidamente con el viento, por lo que fue necesario obtenerlo lo más pronto posible después de su senescencia para poder contar con material suficiente para el experimento. Presentaron tasas de descomposición superiores a las de las dos especies previamente mencionadas de gramíneas, si bien inferiores a las de las hojas y tallos de arbustos y a las de Erodium cicutarium.
La pérdida de masa de Erodium cicutarium, así como de hojas y tallos de arbustos, alcanzó el 60% después de un año de incubación. Es un valor muy alto si se considera el clima frío y el efecto de una estación seca muy marcada. La elevada tasa de descomposición de Erodium cicutarium se explica fácilmente por su elevado contenido inicial de N y NS. La rápida descomposición de la hojarasca de arbustos fue más sorprendente, porque considerábamos a este material como similar a la hojarasca de árboles, que a menudo se descompone lentamente. Sin embargo, al igual que Erodium cicutarium, muestran altos contenidos en N y NS, que son factores estimulantes de la descomposición. El contenido de C es también notable. En general, el contenido de C en la materia orgánica vegetal varía en torno al 47%. Para estos materiales vegetales, el contenido en C excedió al 50%. Como el contenido en lignina – polímero rico en C – fue bajo, debemos suponer que los compuestos ricos en C se hallan en la fracción NS; si son compuestos lábiles, podrían contribuir a explicar la alta tasa de descomposición de este material.
]]>Descomposición de las raíces
Las raíces fueron inicialmente más ricas en lignina que su hojarasca correspondiente (19.6 ± 4.1 y 8.5 ± 2.6%, respectivamente, prueba t bilateral pareada, P=0.011). Fueron obtenidas sin distinguir las raíces muertas de las vivas y fueron enterradas a 10 cm de profundidad. Por lo tanto, su descomposición pudo ser afectada por su calidad inicial, por factores climáticos y por el medio ambiente microbiano. Gill & Burke (2002) estudiaron en zonas templadas de Norteamérica la descomposición de raíces de la gramínea perenne en matas Bouteloua gracilis, a profundidades de 10, 40, 70 y 100 cm. Observaron que la tasa de descomposición de raíces fue aproximadamente 50% más lenta a 1 m que a 10 cm; la pérdida de masa disminuyó linealmente con la profundidad. Estos autores atribuyen esa disminución a condiciones climáticas y especialmente a la reducción de la humedad disponible con la profundidad. Sin embargo, en condiciones mediterráneas, Rovira & Vallejo (1997, 2000) al incubar material vegetal a 5, 20 y 40 cm mostraron que las tasas de descomposición aumentan de 5 a 20 cm, mientras que Schaefer et al. (1985), en un desierto norteamericano, observaron que las especies de hojarasca tuvieron una pérdida de masa más elevada en la superficie que aquellas similares que fueron enterradas. En este último estudio, el material vegetal enterrado difería en calidad de la hojarasca de la superficie, ya que correspondía a raíces de las especies incubadas en la superficie. El efecto de la calidad de la hojarasca se añadió al efecto climático. Sin embargo, incluso si la tasa de descomposición y la perdida de masa durante el primer año de las raíces fueran más bajas que sus correspondientes en la superficie, la diferencia no fue significativa. El efecto del clima puede haber compensado el de las diferentes calidades del material incubado.
Conclusiones
La hipótesis inicial de este trabajo era que el contenido de LIG de las hojas es mayor en descanso largo, y que ello puede ser atribuido a la presencia de especies leñosas. En consecuencia, dado que los materiales ricos en LIG parecen tener una descomposición más lenta, se esperaba una tasa de descomposición menor en el descanso largo. Este no fue el caso. Observamos aquí que el principal factor determinante de la tasa de descomposición fue el contenido de compuestos no estructurales (NS) que probablemente incluyen una parte importante de las sustancias lábiles. Como las gramíneas se caracterizan por un muy bajo contenido en NS, compensado con un elevado contenido en HEM + CEL, mostraron una baja tasa de descomposición pese al hecho que su contenido en LIG fue bajo. Las raíces también se descompusieron lentamente. La fuente principal para la acumulación de MOS debería ser atribuida a los sistemas radiculares y a las gramíneas, especialmente Stipa ichu y Festuca dolichophylla, que mostraron las tasas de descomposición más bajas. Proponemos la hipótesis que la recuperación de la fertilidad del suelo en los períodos de descanso se debe a dos hechos simultáneos: (1) la presencia de hojarasca rica en N (Baccharis incarum, Parastrephia lepidophylla), que liberaría nutrientes en abundancia debido a su rápida descomposición, y (2) la presencia de hojarasca y raíces de gramíneas pobres en N, de descomposición lenta, que contribuirían a la recuperación del stock de materia orgánica.
Agradecimientos
Agradecemos la colaboración de Bruno Buatois (CEFE, CNRS, Francia) y a Claudio Rosales (Instituto de Ecología, La Paz, Bolivia), asistentes técnicos en Francia y Bolivia para este estudio. También agradecemos a Klaas Metselaar (PRI, Wageningen, Holanda) el habernos facilitado el modelo Sahel para la estimación PET y a Monica Moraes (Editora de la revista Ecología en Bolivia, Instituto de Ecología) y Pere Rovira (Universidad de Barcelona) por la traducción del texto al castellano. Este trabajo fue realizado en el marco del programa TROPANDES titulado ‘Fertility management in the tropical Andean mountains: agroecological bases for a sustainable fallow agriculture’ financiado por la Unión Europea (INCO-DCXII ERBIC18CT98-0263).
]]> Referencias
Aber, J. & J. Melillo. 1982. Nitrogen immobilization in decaying hardwood leaf litter as a function of initial nitrogen and lignin content. Canadian Journal of Botany 60: 2263-2269.
Cornelissen, J. H. C. 1996. An experimental comparison of leaf decomposition rates in a wide range of temperate plant species and types. Journal of Ecology 84: 573-582.
[ Links ]Coûteaux, M. M., K. B. McTiernan, B. Berg, D. Szuberla, P. Dardenne & P. Bottner. 1998. Chemical composition and carbon mineralisation potential of Scots pine needles at different stages of decomposition. Soil Biology & Biochemistry 30: 583-595.
C. V. Boucqué. 2000. The filter bag versus the conventional filtration technique for the determination of crude fibre and Van Soest cell wall constituents. Journal of Animal and Feed Sciences 9: 513-526.
Gill, R. A. & I. C. Burke. 2002. Influence of soil depth on the decomposition of Bouteloua gracilis roots in the shortgrass steppe. Plant and Soil 241: 233-242.
[ Links ]Hobbie, S. E. 1996. Temperature and plant species control over litter decomposition in Alaskan tundra. Ecological Monographs 66: 503-522.
[ Links ]Marstop, H. 1996. Influence of carbohydrates, free amino acids, and protein content on decomposition of Lolium multiflorum shoots. Biology and Fertility of Soils 21: 257-263.
[ Links ]Kurz-Besson, C. 2000. Décomposition de litières de pin (Pinus sylvestris, P. halepensis, et P. pinaster) dans un transect climatique européen: rôle de la qualité des litières et du climat. PhD Thesis, Universidad de Paris XI, Orsay. 280 p.
[ Links ]McTiernan, K. B. 1998. The effect of climate on the decomposition of chemical constituents of tree litters. PhD, Universidad de Lancaster, Lancaster. 175 p.
[ Links ]Pérez-Harguindeguy, N., S. Diaz, J. H. C. Cornelissen, F. Vendramini, M. Cabido & A. Castellanos. 2000. Chemistry and toughness predict leaf litter decomposition rates over a wide spectrum of functional types and taxa in central Argentina. Plant and Soil 218: 21-30.
[ Links ]Rovira, P. & V. R. Vallejo. 1997. Organic carbon and nitrogen mineralization under mediterranean climatic conditions: the effects of incubation depth. Soil Biology & Biochemistry 29-4: 1509-1520.
[ Links ]Rovira, P. & V. R. Vallejo. 2000. Decomposition of Medicago sativa debris incubated at different depths under Mediterranean climate. Arid Soil Research and Rehabilitation 14: 265-280.
[ Links ]Schaefer, D., Y. Steinberger & W. G. Whitford. 1985. The failure of nitrogen and lignin control of decomposition in a North American desert. Oecologia 65: 382-386.
[ Links ]Shaw, M. R. & J. Harte. 2001. Control of litter decomposition in a subalpine meadowsagebrush steppe ecotone under climate change. Ecological Applications 11: 12061223.
[ Links ]Tiessen, H., E. Sampaio & I. H. Salcedo. 2001. Organic matter turnover and management in low input agriculture of NE Brazil. Nutrient Cycling in Agroecosystems 61: 99-103.
[ Links ]Trofymow, J. A., T. R. Moore, B. Titus, C. Prescott, I. Morrison, M. Siltanen, S. Smith, J. Fyles, R. Wein, T. C. Camir, L. Duschene, L. Kozak, M. Kranabetter &
S. Visser. 2002. Rates of litter decomposition over 6 years in Canadian forests: influence of litter quality and climate. Canadian Journal of Forest Research 32: 789-804.
van Soest, P. J. & J. B. Robertson. 1985. Analysis of forage and fibrous foods: A laboratory manual for animal science. Cornell University Pub., Ithaca. 613 p.
[ Links ]Weisbach, C., H. Tiessen & J. J. Jimenez Osornio. 2002. Soil fertility during shifting cultivation in the tropical Karst soils of Yucatán. Agronomie 22: 253-263.
[ Links ]Wezel, A. & J. Haigis. 2002. Fallow cultivation system and farmers’ resource management in Niger, West Africa. Land Degradation and Development 13: 221-231.
]]>