El ciclo global del carbono y sus consecuencias en la fotosíntesis en el Altiplano boliviano
Carbon global cycle and its consequences on photosynthesis in the Bolivian Altiplano
B. Saugier1 & J.Y. Pontailler2
1Ecologie, Systématique et Evolution, bât. 362, Université Paris-Sud, 91405 Orsay, Cedex, Francia e-mail: bernard.saugier@ese.u-psud.fr
2jean-yves.pontailler@ese.u-psud.fr
Resumen
El ciclo global del carbono se describe tomando en cuenta los intercambios entre la atmósfera y la biosfera continental, así como las modificaciones causadas por las actividades humanas: deposiciones nitrogenadas, aumento del CO2 atmosférico y el calentamiento climático inducido. Estos cambios globales tienen como consecuencia un aumento de producción vegetal y un almacenamiento de carbono en la biomasa de los árboles y en la materia orgánica del suelo. Como todos los ecosistemas terrestres, el Altiplano boliviano con sistemas agrícolas que respetan un descanso largo está sometido a estos cambios globales. La respuesta fotosintética a la luz y al CO2 ha sido medida en hojas de algunas especies en la sucesión vegetal post-cultivo en el Altiplano boliviano. La fotosíntesis máxima es alta para las plantas pioneras y es baja para plantas del final de la sucesión, como se podía esperar. Las especies espontáneas (Chondrosum simplex, Erodium cicutarium, Lupinus otto-buchtienii, Baccharis incarum, Parastrephia lepidophylla, Stipa ichu, Aristida asplundii) tienen una fotosíntesis superior a la medida en una especie cultivada, la quinua (Chenopodium quinoa), lo que es sorprendente. En todos los casos, la fotosíntesis de la hoja aumenta fuertemente con el CO2.Al duplicar artificialmente la tasa de CO2incide en una duplicación de la fotosíntesis de la hoja. Un aumento del CO2atmosférico podría contribuir al aumento de la producción vegetal durante el descanso y entonces en la reducción de la duración del descanso en el Altiplano boliviano.
Palabras clave: Balance de carbono, Cambio climático, Biomasa, Fotosíntesis, Altiplano boliviano.
Abstract
The global carbon cycle is described considering the carbon fluxes between the atmosphere and the terrestrial biosphere, and the perturbations due to human activity: nitrogen deposition, increased atmospheric CO2 and induced climatic warming. These global changes increase plant production and then carbon storage in tree biomass and soil organic matter. The Bolivian Altiplano with long fallow agrosystems is affected by these global changes as all other terrestrial ecosystems. The leaf photosynthetic response to light and CO2 was measured for a few plant species of fallow in the Bolivian Altiplano. As expected, maximum photosynthesis is high for pioneer plants and low for plants of late succession. Wild species (Chondrosum simplex, Erodium cicutarium, Lupinus otto-buchtienii, Baccharis incarum, Parastrephia lepidophylla, Stipa ichu, Aristida asplundii) exhibit higher photosynthetic rates than quinoa (Chenopodium quinoa) crop, which is surprising. In each species leaf photosynthesis strongly increases with CO2, almost doubling when CO2 is artificially doubled. A rise in atmospheric CO2 may increase plant productivity during fallow and thus induce a reduction of fallow duration in the Bolivian Altiplano.
Key words: Carbon balance, Climatic change, Biomass, Photosynthesis, Bolivian Altiplano.
]]> Introducción
La puna, como todos los ecosistemas terrestres, está sometida a cambios globales tales como el aumento de la concentración de CO2 atmosférico, el recalentamiento que lo acompaña, entre otros. Estos cambios globales se traducen en un aumento de la fijación de CO2 por las especies vegetales tanto cultivadas como espontáneas. Es interesante predecir su comportamiento futuro en sistemas de cultivo con descanso largo (unos 10 años) que son practicados de manera tradicional en los Andes. Uno de los objetivos de hacer descansar el suelo es la restauración parcial de la fertilidad del suelo que existía antes del cultivo. Bien es sabido que la reserva de materia orgánica de un ecosistema disminuye rápidamente tras su puesta en cultivo. En los ecosistemas tropicales basta con unos diez años de cultivo para que la reserva de carbono del suelo se reduzca a la mitad, lo que provoca una disminución de la capacidad de intercambio catiónico y más generalmente de la fertilidad. La función del descanso es restablecer una fracción de la reserva de carbono perdida y de la microflora asociada que garantice el reciclaje de los elementos minerales. Las especies espontáneas, que colonizan el terreno después de los cultivos, contribuyen a este balance de carbono y podrían responder fuertemente al aumento vigente del contenido de CO2 atmosférico.
Por esta razón, presentamos primero nuestros conocimientos sobre el ciclo global del carbono, sobre su perturbación por las actividades del hombre y sobre el papel de la vegetación en el balance de carbono mundial. Deducimos luego algunas consecuencias para el Altiplano boliviano y medimos la fijación del carbono por algunas especies que colonizan el terreno en descanso. Estas mediciones en hojas de la respuesta fotosintética a la luz y al CO2 permiten predecir el comportamiento futuro de estas especies frente a un aumento de la concentración del CO2 atmosférico.
Análisis del ciclo global del carbono
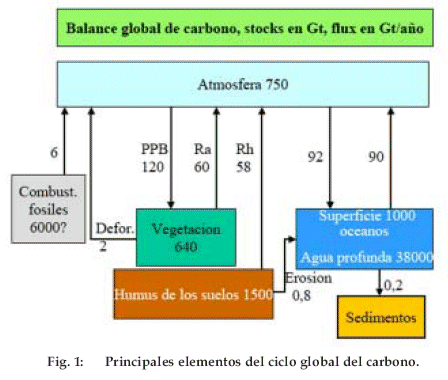
La composición de la atmósfera terrestre ha variado mucho en el curso de tiempos geológicos. Antes de la aparición de la vida, la atmósfera estaba dominada por gas carbónico producido por las erupciones volcánicas, como es aún el caso para los planetas sin vida que nos rodean, Venus y Marte (Tabla 1).
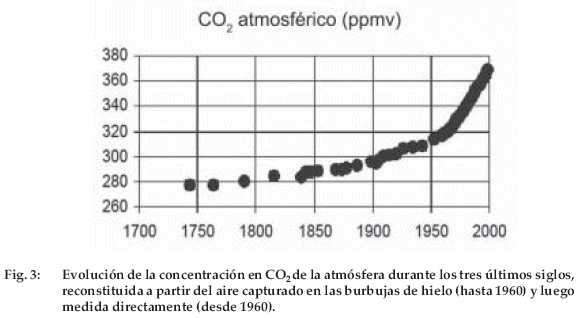
La fotosíntesis, que comenzó hace por lo menos tres mil millones de años, fijó cantidades considerables de carbono y liberó oxígeno que primero oxidó la superficie terrestre, primitivamente reductora y permitió la acumulación de oxígeno libre desde hace aproximadamente 2 mil millones de años (Schlesinger 1991). En el curso de tiempos geológicos, la absorción del CO2 por la fotosíntesis y la incorporación de grandes cantidades de carbono en las rocas sedimentarias, redujeron progresivamente el contenido de CO2 del aire. Al final de la era secundaria, cuando se extinguieron los dinosaurios, aún había aproximadamente dos veces más de CO2 que en 1800 antes de la era industrial (Berner 1994). La composición actual de la atmósfera resulta de un equilibrio entre procesos biológicos como la fotosíntesis y la respiración, así como de procesos fisicoquímicos como la absorción del CO2 en aguas frías océanicas, subsaturadas de CO2 y su liberación por aguas calientes, sobresaturadas de CO2. Es sabido, por el análisis de las burbujas de aire atrapadas en los hielos de la Antártica desde hace 400.000 años (Fig. 3), que el contenido de CO2 del aire varió al mismo tiempo que la temperatura de la Tierra, pasando de aproximadamente 200 ppm (partes por millón en volumen) durante los periodos glaciales a 280 ppm durante los periodos más calientes, como el que conocemos desde hace casi 10.000 años (Barnola et al. 1991).
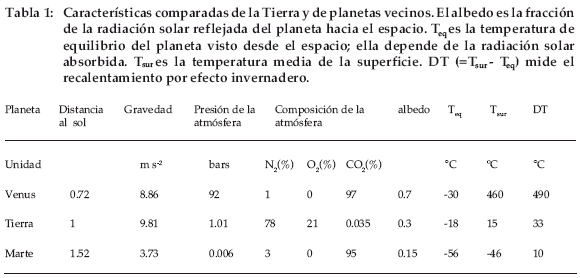
El equilibrio natural que condujo a estos bajos contenidos de CO2 está siendo actualmente modificado a escala global por las actividades del hombre, las cuales liberan cantidades crecientes de CO2 al quemar combustibles fósiles (carbón, petróleo, gas natural) y la madera de los bosques eliminados para la agricultura o el pastoreo. La figura 1 presenta el estado de nuestros conocimientos sobre los principales depósitos de carbono en la superficie de la Tierra y sobre los flujos anuales intercambiados entre estos depósitos. Podemos observar que la biomasa continental (principalmente la madera de los árboles) contiene 640 Pg o GtC (miles de millones de toneladas de carbono), es decir un poco menos que el carbono almacenado en la atmósfera en forma de CO2(750 GtC) y que la materia orgánica de los suelos (1.500 GtC) representa dos veces más CO2 que en la atmósfera. Los flujos intercambiados anualmente por la fotosíntesis (PPB = 120 GtC/año) y la respiración de las plantas (Ra = 60 GtC/año) y de los animales (Rh = 58 GtC/año), que provienen principalmente de los descomponedores del suelo, son elevados respecto al tamaño de los depósitos, lo que significa una renovación rápida del CO2 atmosférico. La figura 1 muestra también los flujos relativos a los combustibles fósiles (un promedio de más de 6 GtC/año de 1990 a 1999) y a la deforestación (2 GtC/año, cifra muy imprecisa) y los flujos intercambiados entre la atmósfera y la superficie de los océanos, con una absorción por las aguas frías de las altas latitudes un poco superior al desprendimiento por las aguas calientes a bajas latitudes. La absorción de CO2 por las aguas de superficie se transmite a las aguas profundas con un tiempo de renovación de varios siglos, lo que significa que el exceso de CO2 del océano con respecto a su valor preindustrial se transmitirá a la atmósfera durante varios siglos, incluso si hoy se detuviera toda expulsión de CO2 a la atmósfera.
]]>
Desde alrededor de 1990, un grupo de científicos coordina regularmente los datos relativos a los cambios climáticos y sus causas (ciclo del carbono); se conoce bajo el acrónimo de IPCC (Intergovernmental Panel on Climatic Change) y se divide en tres grupos. El primer grupo estudia el efecto invernadero, sus causas y consecuencias sobre el clima. El segundo grupo estudia las consecuencias de los cambios globales sobre los ecosistemas y las actividades del hombre, y el tercer grupo estudia las diversas posibilidades para limitar el efecto invernadero, con sus consecuencias socioeconómicas y políticas. En el sitio Web www.ipcc.ch se pueden consultar los últimos informes de estos grupos.
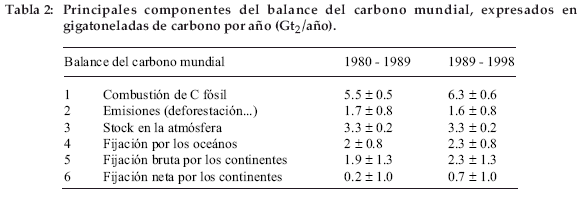
La tabla 2 muestra el balance del intercambio anual de carbono entre los principales depósitos, para los períodos recientes. Podemos observar que de las 7.9 mil millones de toneladas de carbono expulsadas anualmente por las actividades del hombre de 1990 a 1998, un poco menos de la mitad (3.3) se acumula en la atmósfera en forma de CO2, siendo el resto absorbido en partes iguales por los océanos y por la biosfera continental. ¿A dónde va el carbono absorbido por los continentes? Los únicos depósitos posibles son la biomasa y los suelos. La biomasa está formada principalmente de la madera de los árboles. ¿Hay algún aumento de la cantidad de madera viva en el mundo en las zonas no afectadas por la deforestación? ¿La reserva de materia orgánica de los suelos está aumentando? ¿Con qué distribución geográfica? ¿Este aumento será duradero? Para tratar de responder a estas interrogaciones, presentaremos las mediciones del CO2 atmosférico, seguido de los métodos que permiten estudiar los intercambios de CO2 a nivel de un ecosistema y luego de los continentes en su conjunto. Luego analizaremos lo que sucede en un ecosistema en equilibrio con su medio y en un ecosistema cuya producción primaria está en aumento. Finalmente, analizaremos los argumentos que sugieren un aumento de la producción primaria en el curso de las últimas décadas. El lector interesado podrá encontrar algunos desarrollos complementarios en Saugier (1996).
Las mediciones del CO2 atmosférico
En 1958, como resultado del año Geofísico Internacional, Keeling estableció la primera estación de medición del CO2 atmosférico en una isla del archipiélago Hawai, Mauna Loa. Echando un vistazo en un mapa, vemos que este archipiélago se encuentra en medio del Pacífico, a 20° N de latitud. El sitio se eligió por ser representativo de la concentración atmosférica promedio de esta latitud en el hemisferio norte. Es sabido que la vegetación terrestre, por los procesos de fotosíntesis y de respiración, modifica el contenido de CO2 del aire. Al instalarse en una pequeña isla, Keeling minimizaba este efecto. Además para reducir también los efectos locales, decidió instalar la estación a 3.000 metros de altitud en la cumbre de un volcán (lo que obliga a eliminar los datos cuando el viento desvía el humo del volcán hacia el sitio de medición, una situación afortunadamente rara). Logró mantener la estación en funcionamiento hasta hoy, a pesar de numerosos ataques de la administración que deseaba reducir los gastos de mantenimiento. Se constató rápidamente que el CO2 no era constante, sino que aumentaba regularmente. Cuando un número suficiente de años de medición estuvo disponible, se constató que este aumento era cada vez más rápido. Surgieron entonces varias preguntas: ¿Este aumento es un fenómeno general? ¿Cuál es la causa? ¿Desde cuándo se produce?
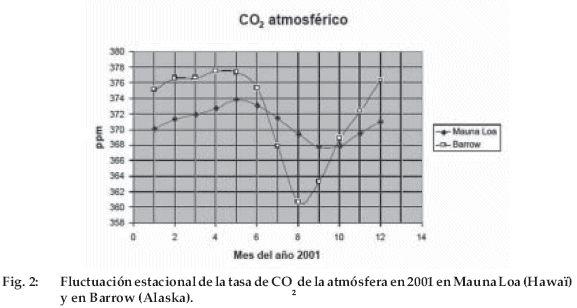
A primera vista, el fenómeno parecía general, ya que la atmósfera es un medio muy agitado por los vientos y su composición es relativamente estable de un punto al otro del planeta. Esto se pudo verificar instalando otras estaciones de medición, además de la de Mauna Loa. Actualmente existen aproximadamente sesenta; la mayoría toman muestras de aire en botellas que se envían a un laboratorio central para ser analizadas, las otras como Mauna Loa registran continuamente el contenido de CO2 del aire y establecen promedios diarios y mensuales. ¿Qué indican esas numerosas mediciones? Primero, que el aumento del CO2 atmosférico es general, con una tasa de aumento que no depende de la estación de medición. Luego se constató que el contenido de CO2 presentaba fluctuaciones estacionales muy marcadas y que la amplitud de estas fluctuaciones variaba con la latitud. Así, vemos en la figura 2 que en Mauna Loa se alcanza la concentración máxima en mayo y la mínima en septiembre-octubre. En Barrow (Alaska), la concentración aumenta ligeramente de diciembre a mayo y disminuye bruscamente de mayo a agosto. La amplitud estacional es de 17 ppm en Barrow, en vez de 6 ppm en Mauna Loa. En el hemisferio sur, las variaciones estacionales son muy pequeñas (<1 ppm) e invertidas con respecto a las del hemisferio norte. Para saber si estas fluctuaciones eran provocadas por los intercambios con los océanos o con los continentes, Keeling y sus colegas midieron la relación isotópica 13C/12C del CO2 atmosférico. Se sabe en efecto que los intercambios con los océanos prácticamente no modifican esta relación, mientras que en los continentes, la mayoría de las plantas - llamadas en C3 porque el primer producto de la fotosíntesis es una molécula de tres átomos de carbono - utilizan principalmente para su fotosíntesis el 12 C, lo que provoca un enriquecimiento del CO2 del aire en 13 C. La conclusión fue obvia: en su gran mayoría, las fluctuaciones estacionales del CO2 proceden de los intercambios con los continentes. Ahora entendemos fácilmente por qué la amplitud de estas variaciones depende de la latitud: en el hemisferio norte, que posee una gran superficie continental, las fluctuaciones aumentan con el carácter estacional del clima y por consiguiente con la latitud; en el hemisferio sur, hay menos continentes que en el hemisferio norte y sobre todo muy pocos continentes más allá de 40° S, lo que explica las fluctuaciones estacionales poco marcadas.
Para saber desde cuándo el CO2 aumenta, varios equipos de investigadores franceses y suizos tuvieron la idea de analizar la composición del aire atrapado en los hielos de la Antártica o de Groenlandia (que no se funden nunca debido a las bajas temperaturas que allí reinan). Cuando en esas regiones nieva, la nieve se acumula y progresivamente va formando hielo cuando es sometida a un peso suficiente de la columna de nieve que está por encima. En ese momento, el aire está atrapado en las burbujas sin contacto con el aire exterior y conserva la composición que tenía en el momento del cierre de las burbujas. Perforando el hielo en la Antártica que encierra más de 3.000 metros de espesor de hielo en puntos como Vostok (Barnola et al. 1991), se puede retroceder hasta aproximadamente 400.000 años en el tiempo. El análisis del aire atrapado recientemente permitió trazar la curva de la figura 3 que concuerda bien con las mediciones directas del CO2 atmosférico realizadas en Mauna Loa. Podemos observar que la concentración ha aumentado muy lentamente al principio a partir de 1750 y luego cada vez más rápidamente. Como la revolución industrial comenzó después de 1850, el aumento estaba sin duda asociado a los considerables desmontes que acompañaron el aumento de la población humana. El contenido, que antes de 1750 era de aproximadamente 280 ppm, sobrepasó 370 ppm en 2001 (Fig. 3). ¿Cuál es la causa de este aumento?
]]> Rápidamente se pensó que se debía al uso del combustible fósil, primero carbón y luego petróleo y gas natural. Se ha demostrado que el consumo de esos combustibles ha crecido exponencialmente desde aproximadamente burbujas sin contacto con el aire exterior y fotosíntesis el 12C, lo que provoca un conserva la composición que tenía en el enriquecimiento del CO2 del aire en 13C. La momento del cierre de las burbujas. Perforando conclusión fue obvia: en su gran mayoría, las el hielo en la Antártica que encierra más de fluctuaciones estacionales del CO2 proceden 3.000 metros de espesor de hielo en puntos de los intercambios con los continentes. Ahora como Vostok (Barnola et al. 1991), se puede entendemos fácilmente por qué la amplitud de retroceder hasta aproximadamente 400.000 estas variaciones depende de la latitud: en el años en el tiempo. El análisis del aire atrapado hemisferio norte, que posee una gran superficie recientemente permitió trazar la curva de la continental, las fluctuaciones aumentan con el figura 3 que concuerda bien con las mediciones carácter estacional del clima y por consiguiente directas del CO2 atmosférico realizadas en con la latitud; en el hemisferio sur, hay menos Mauna Loa. Podemos observar que la continentes que en el hemisferio norte y sobre concentración ha aumentado muy lentamente todo muy pocos continentes más allá de 40° S, al principio a partir de 1750 y luego cada vez lo que explica las fluctuaciones estacionales más rápidamente. Como la revolución poco marcadas. industrial comenzó después de 1850, el aumentoPara saber desde cuándo el CO2 aumenta, estaba sin duda asociado a los considerables varios equipos de investigadores franceses y desmontes que acompañaron el aumento de la suizos tuvieron la idea de analizar la población humana. El contenido, que antes de composición del aire atrapado en los hielos de 1750 era de aproximadamente 280 ppm, la Antártica o de Groenlandia (que no se funden sobrepasó 370 ppm en 2001 (Fig. 3). ¿Cuál es la nunca debido a las bajas temperaturas que allí causa de este aumento? reinan). Cuando en esas regiones nieva, la nieve Rápidamente se pensó que se debía al uso se acumula y progresivamente va formando del combustible fósil, primero carbón y luego hielo cuando es sometida a un peso suficiente petróleo y gas natural. Se ha demostrado que el de la columna de nieve que está por encima. En consumo de esos combustibles ha crecido ese momento, el aire está atrapado en las exponencialmente desde aproximadamente 1920 con periodos de relativo estancamiento debidos a ambas guerras mundiales y a los enormes aumentos del precio del petróleo (choques petroleros). Como el carbono fósil fue producido por la fotosíntesis de plantas en C3 (las únicas que existían hasta hace aproximadamente 10 millones de años), éste está empobrecido en 13C y su combustión debe provocar un empobrecimiento en 13C del CO2 atmosférico. Este fenómeno fue claramente demostrado. Sin embargo, no permite una conclusión definitiva, ya que la madera de los árboles también está empobrecida en 13C: no podemos entonces saber de esta manera si el CO2 en exceso procede del combustible fósil o de la deforestación. Hasta ahora, se ha podido establecer el consumo del combustible fósil con menos del 10% de error, sin embargo, las estimaciones de expulsiones de CO2 asociadas a la deforestación tienen un error del 50% (Tabla 2).
En la tabla 2, la absorción de CO2 por la biosfera continental se calcula por la diferencia entre las emisiones y la acumulación de carbono en la atmósfera y los océanos, habiendo sido calculado este último con ayuda de un modelo y con una posibilidad de error considerable. Ciertos investigadores pusieron en duda esta absorción de carbono por la biosfera continental, pero por lo menos dos trabajos confirman la formación de un gran pozo de carbono por la vegetación terrestre del hemisferio norte. Uno de ellos (Ciais et al. 1995) utiliza un modelo inverso para calcular las fuentes y pozos biosféricos y oceánicos de carbono por bandas de latitud a partir de la red de mediciones de CO2 y de la relación 13C/12C. El otro (Denning et al. 1995) calcula dichos pozos y fuentes comparando el gradiente latitudinal de CO2 con lo que se debería obtener teniendo en cuenta la distribución preferencial de las fuentes de carbono fósil en el hemisferio norte.
Así, los estudios del ciclo global del carbono muestran que en la atmósfera sólo se encuentra aproximadamente el 40% del CO2 liberado por la combustión del carbono fósil y la deforestación. El resto es absorbido por los océanos y ecosistemas terrestres en partes aproximadamente iguales. El almacenamiento en los océanos se efectúa primero en la parte superficial, una capa de alrededor de 100 metros de espesor bien mezclada por los vientos (Figura 1). El almacenamiento a mayor plazo se realiza en la parte profunda del océano y depende del intercambio de agua entre parte superficial enriquecida en CO2 y parte profunda, con un tiempo de respuesta de cinco a diez siglos. El almacenamiento en los ecosistemas terrestres está limitado principalmente a la madera de los árboles y a la materia orgánica de los suelos, que juntas representan una reserva de carbono que es alrededor de tres veces la reserva de carbono del CO2 atmosférico.
El papel principal de la vegetación en el balance de carbono mundial
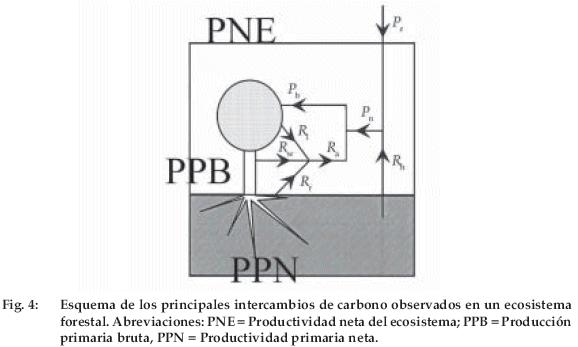
La figura 4 detalla el intercambio de carbono de un ecosistema forestal. Si llamamos PPB a la producción primaria bruta o suma de la fijación diurna de carbono por el follaje y Ra a la respiración autótrofa (que es la suma de la respiración nocturna de las hojas y de la respiración durante 24 horas de los otros órganos), vemos que la productividad primaria neta PPN es igual a PPB - Ra. Además, los organismos heterótrofos (principalmente los microorganismos del suelo) respiran también (Rh), de manera que podemos definir una productividad neta del ecosistema PNE, igual a la diferencia entre PPN y Rh. PNE representa la suma de 24 horas (o de un año) del flujo neto de carbono sobre el ecosistema FC, que se mide con un gran recinto que cubre una porción de suelo para una vegetación baja o con métodos micrometeorológicos para un bosque. En un mundo en equilibrio donde la concentración de CO2es constante (como ocurrió en los últimos 10.000 años hasta alrededor de 1750), no hay intercambio neto de CO2 entre océanos y atmósfera o entre continentes y atmósfera, por consiguiente la PNE de un año es nula. El aumento actual del CO2 atmosférico introduce a un desequilibrio que provoca una absorción neta por los océanos y continentes.
Esta absorción está asociada para los océanos a la diferencia de presión parcial de CO2 entre la atmósfera y aguas superficiales. Para los continentes es un fenómeno principalmente biológico que favorece la absorción de carbono: la estimulación de la productividad primaria neta que es seguida con retraso por un aumento de la respiración heterótrofa, resulta en una acumulación de carbono (PNE>0). Esta estimulación tiene varias causas posibles:
La PNE es pues un término fundamental para saber si un ecosistema particular absorbe o expulsa carbono a la atmósfera. Con frecuencia no es fácil sacar conclusiones, ya que la PNE resulta de una diferencia entre dos procesos de importancia comparable: una absorción neta de carbono por el ecosistema de día y una expulsión durante la noche. Un pequeño error en uno de esos procesos provoca entonces un gran error en PNE. Además, para predecir la evolución del balance de carbono de los ecosistemas con los cambios globales, hay que conocer no sólo el flujo global de carbono Fc a nivel del ecosistema, sino también los diversos procesos que componen este flujo y sus variaciones con estos cambios globales. Se emprendió un estudio de gran alcance para medir de manera continua este flujo Fc sobre varios tipos de ecosistemas; contamos actualmente con 150 sitios de mediciones en el mundo, reunidos en la base de datos FLUXNET.
Consecuencias sobre la fotosíntesis en el Altiplano boliviano
En el Altiplano boliviano, estos cambios globales podrían modificar las condiciones de crecimiento de las especies cultivadas, como la papa o la quinua, como de las especies espontáneas que colonizan el descanso. Se midió la fijación de carbono de algunas especies claves de la puna boliviana con la finalidad de predecir el comportamiento futuro de estas especies frente a un aumento de la concentración de CO2 atmosférico. Se estudiaron detalladamente siete especies nativas y se hicieron algunas mediciones en la quinua cultivada (Chenopodium quinoa). Erodium cicutarium y Chondrosum simplex son pioneras, Lupinus otto-buchtienii es una especie intermedia; Parastrephia lepidophylla y Baccharis incarum son arbustos a menudo co-dominantes en los descansos de edad; Stipa ichu y Aristida asplundii son gramíneas perennes; Aristida asplundii con amplio rango de tolerancia está más representada en descansos viejos que Stipa ichu.
Metodología
Utilizamos un sistema portátil para mediciones de fotosíntesis, el Li-Cor 6400 que estaba equipado con una cámara que contenía no más de 6 cm2 de hoja con la posibilidad de hacer variar la iluminación (diodos que emiten en rojo y en azul de 0 a 2000 µmol m-2 s-1), el contenido de CO2(de 0 a 2.000 ppm en volumen) y la temperatura (de –2°C a +10°C con respecto al ambiente). Los individuos representativos de las especies vegetales estudiadas estaban localizados en dos sitios - Patacamaya y Patarani -distantes de unos kilómetros y situados a 150 km al sur de La Paz (17° 15’ S, 67° 55’ W, altitud 3.950 m). La media anual de temperatura en Patacamaya es de 11.2°C y de precipitación es de 400 mm. Las mediciones se efectuaron en marzo 2001 al final de una estación de lluvias particularmente abundante, con un suelo todavía muy húmedo. El reducido tamaño de las hojas generalmente no permitió que se llenara la cámara de mediciones. La superficie contenida en la cámara era de aproximadamente 2 cm2 y se medía cuidadosamente tras las mediciones de fotosíntesis. La mayoría de las hojas eran planas, excepto Stipa ichu - una gramínea de hojas enrolladas sobre su eje - y Parastrephia lepidophylla, que presenta hojas cilíndricas. En ese caso, utilizamos la superficie de un semicilindro para el cálculo de fotosíntesis (igual a p/2 ó 1.57 veces la superficie proyectada).
El Li-Cor 6400 posee un sensor de presión y corrige los valores medidos de CO2 para la diferencia de presión con la presión normal al nivel del mar. Los valores medidos de asimilación del CO2, expresados en µmol m-2 s1, se utilizaron entonces sin corrección.
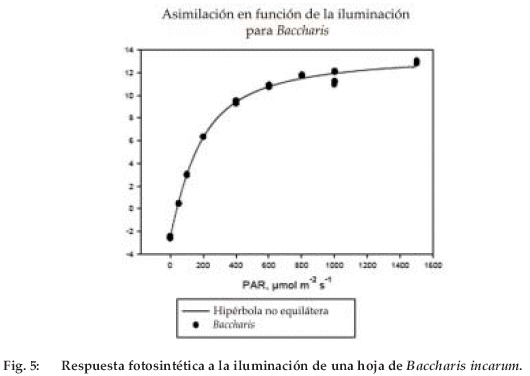
La figura 5 presenta una curva típica de respuesta fotosintética a la iluminación, ajustada con ayuda del software Sigmaplot por una hipérbola no equilátera, del tipo:
]]>Donde A es la asimilación neta de CO2(medida); Pm, la fotosíntesis bruta máxima; a, el rendimiento cuántico aparente; I, la iluminación; c, el parámetro de curvatura y R, la respiración en la oscuridad. El valor máximo de A es Am = Pm –R. Los parámetros Pm, a y R se ajustaron basándose en las mediciones de varias hojas de cada especie y se sacó el promedio de ellas para obtener valores representativos por especie. Como R varía en función de la capacidad fotosintética, preferimos presentar la relación R/Pm, cuyo valor es cercano a 0.1 para diversas especies de árbol (Ceulemans & Saugier 1991).
Resultados
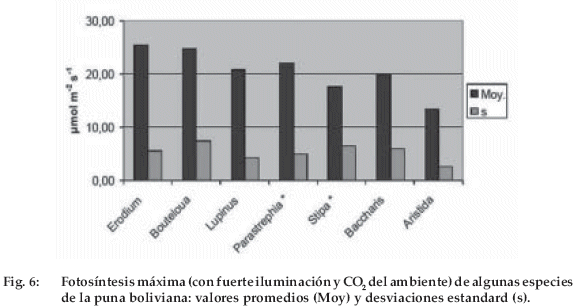
Los resultados se presentan en la figura 6, las especies están ordenadas de izquierda a derecha en función del orden supuesto de aparición en la sucesión ecológica. Los valores Pm son elevados y disminuyen de 25 a menos de 15 µmol m-2 s-1 en el curso de la sucesión. Los arbustos tales como Parastrephia lepidophylla y Baccharis incarum tienen una fotosíntesis tan elevada como la de las herbáceas, tales como Lupinus otto-buchtienii, Stipa ichu y Aristida asplundii. El coeficiente de variación de Pm está comprendido entre 0.2 y 0.3, salvo para Stipa ichu (0.4) cuya escasa superficie foliar es difícil de estimar.
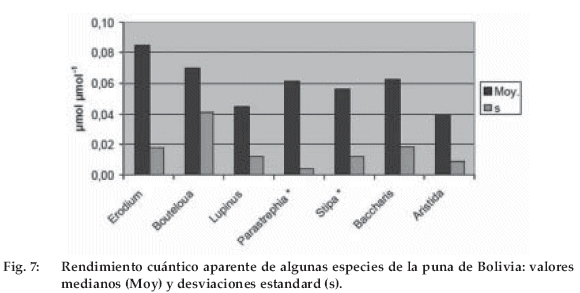
Los valores de rendimiento cuántico aparente (pendiente al origen de la relación asimilación/iluminación, Figura 7) varían entre 0.04 y 0.09 con un promedio de alrededor de 0.06 y un coeficiente de variación de 0.2-0.3, salvo para Stipa ichu (0.6). Estos valores son elevados pero compatibles con los valores publicados de rendimiento cuántico (promedio de 0.05–0.06 para las plantas que crecen al exterior). Como en el caso de los valores de fotosíntesis máxima, se nota que las especies pioneras tienen valores más elevados que las especies al final de la sucesión.
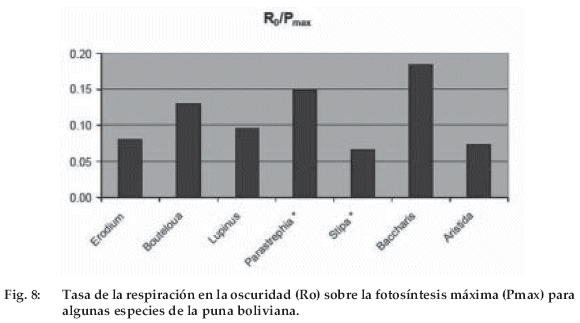
Los valores de la relación R0/Pmax (respiración en la oscuridad sobre fotosíntesis máxima, Figura 8) varían entre 0.06 (Stipa ichu) y 0.18 (Baccharis incarum). Son más elevados en los arbustos (Parastrephialepidophylla y Baccharis incarum) que en las herbáceas, con un valor intermediario para Lupinus otto-buchtienii que debe incorporar el costo de la fijación de nitrógeno en su respiración.
]]>
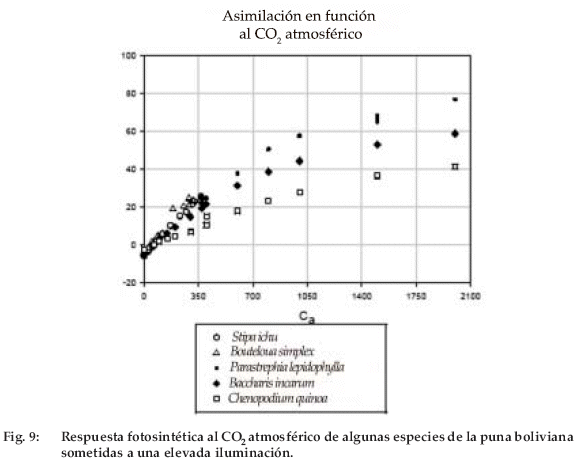
La respuesta al CO2 se estudió en cinco especies: Stipa ichu, Chondrosum simplex, Baccharis incarum, Parastrephia lepidophylla y Chenopodium quinoa. Ésta permite hacerse una a mostrar que esta hipótesis es relativamente idea de la manera en que estas especies se aceptable para árboles jóvenes (Besford et al. comportarán en respuesta al aumento en curso 1998), pero es más discutible para las plantas del CO2atmosférico, con la hipótesis fuerte de herbáceas, que carecen de órganos de que no hay aclimatación del aparato almacenamiento del carbono asimilado frente fotosintético al CO2elevado. Los experimentos al exceso de CO2. La figura 9 muestra estas en las herbáceas y en árboles jovenes tienden respuestas fotosintéticas al CO2 del aire para una hoja de cada una de las cinco especies, sometidas a una elevada iluminación (2.000 µmol m-2 s-1). Las curvas se prolongan a altas concentraciones para tres de esas especies. La forma general es idéntica para las diversas especies, solo difieren los valores absolutos de la asimilación a una determinada concentración. La relación A700/A350 de la asimilación a 700 ppm sobre la asimilación a 350 ppm varía entre 1.8-2. Así, si la aclimatación es relativamente baja a CO2 elevado, deberíamos obtener una fuerte respuesta de estas especies al aumento en curso del contenido atmosférico de CO2. La quinua tiene una asimilación más baja que las especies silvestres, pero presenta una relación A700/A350 de aproximadamente 2.0, como las especies silvestres. Esta relación (obtenida tras el ajuste de las curvas por una hipérbola no equilátera) es de 2.0 para Parastrephia lepidophylla, 1.86 para Baccharis incarum y 1.98 para la quinua, respectivamente. Podríamos entonces esperar un aumento de la presencia de Parastrephia lepidophylla, pero muchos otros factores son suceptibles de modificar esta hipótesis.
Discusión y conclusión
El ciclo global del carbono indica claramente que los ecosistemas terrestres se comportan actualmente como un pozo considerable de carbono, que representa alrededor del 30% de las emisiones de CO2 causadas por las actividades del hombre. La mayor parte de este pozo parece resultar de un aumento de la producción vegetal asociado al aumento del CO2 atmosférico y al recalentamiento global. Una parte viene del aumento de la producción de los cultivos, praderas y bosques aprovechados por el hombre.
En el Altiplano boliviano estos cambios globales pueden modificar de manera considerable las condiciones de crecimiento, tanto de las especies cultivadas, como la papa o la quinua, como de las espontáneas. Las especies de la puna confirman la tendencia esperada de una disminución de la capacidad fotosintética con el orden supuesto de aparición de las especies en la sucesión. Sin embargo, los valores obtenidos son muy elevados, particularmente para los arbustos como Parastrephia y Baccharis, que presentan una fotosíntesis netamente más elevada que la de la especie cultivada estudiada, la quinua. Estos valores se obtuvieron en un suelo particularmente húmedo, al final de una abundante estación de lluvias. Sería interesante verificar cómo evolucionan durante la estación seca. Es posible que estos valores elevados sólo se mantengan durante un corto periodo, pero la alimentación en agua depende también estrechamente del enraizamiento, que no estudiamos en esta oportunidad. Finalmente, observamos que una duplicación de contenido de CO2 con respecto al actual provoca casi una duplicación de la respuesta instantánea de la fotosíntesis, lo que deja suponer que las especies del Altiplano pueden responder fuertemente al aumento en curso del contenido atmosférico de CO2. El aumento del CO2 atmosférico podría contribuir al aumento de la producción vegetal durante el descanso y abrir entonces una posibilidad para reducir la duración del descanso.
Referencias
Barnola, J. M., P. Pimienta, D. Raynaud & Y. S. Korotkevich. 1991. CO2-climate relationship as deduced from the Vostok ice core: a re-examination based on new measurements and on a re-evaluation of the air dating. Tellus 43(B): 83-90.
]]>Berner, R. A. 1994. Geocarb II: A revised model of atmospheric CO2 over phanerozoic time. American Journal of Science 294: 56-91.
[ Links ]Besford, R.T., M. Mousseau & G. Matteucci. 1998. Biochemistry, physiology and biophysics of photosynthesis. Pp. 29-78. En: P.G. Jarvis (ed.). European Forests and Global Change: the Likely Impacts of Rising CO2 and Temperature. Cambridge University Press. Cambridge.
[ Links ]Ceulemans, R. & B. Saugier. 1991. Photosynthesis. Pp. 21-51. En: A. S Rajhavendra (ed.). Physiology of Trees, Vol. 2. J. Wiley and Sons, Nueva York.
[ Links ]Ciais, P., P. P. Tans, J.W.C. White, M. Trolier, R. J. Francey, J. A. Berry, D. R. Randall, P.J. Sellers, J. G. Collatz & D. S. Schimel. 1995. Partitioning of ocean and land uptake of CO2, as inferred by measurements of changes in 13C from the NOAA Climate Monitoring and Diagnostics Laboratory Global Air Sampling Network. J. Geophys. Research 100 (D3): 5051-5070.
Denning, A. S., I. Y. Fung & D. Randall. 1995. Latitudinal gradient of atmospheric CO2 due to seasonal exchange with land biota. Nature 376: 240-243.
Saugier, B. 1996. Végétation et atmosphère. Flammarion (Collection Dominos), Paris. 126 p.
[ Links ]Schlesinger, W. H. 1991. Biogeochemistry: An analysis of global change. Academic Press, San Diego, California.
[ Links ] ]]>