Dinámica sucesional de la vegetación en un sistema agrícola con descanso largo en el Altiplano central boliviano
Plant succesional dynamics of long term fallow in the central Altiplano of Bolivia
Teresa Ortuño1, Stephan Beck2 & Lina Sarmiento3
1Museo Nacional de Historia Natural, Herbario Nacional de Bolivia, Casilla 10077 - Correo Central, La Paz, Bolivia, email: casstol@yahoo.com
2Instituto de Ecología, Herbario Nacional de Bolivia, Universidad Mayor de San Andrés, Casilla 10077 - Correo Central, La Paz, Bolivia, e-mail: lpb.dir@acelerate.com ]]>
Resumen
Se estudió la dinámica sucesional de la vegetación en parcelas en descanso agrícola en el Altiplano central boliviano, donde se practica una agricultura con descansos largos en sectores manejados comunalmente. El objetivo fue analizar cambios sucesionales de la diversidad de especies, de la estructura vertical de la vegetación, de la abundancia de las diferentes formas de vida que intervienen en la sucesión. Los resultados indican pocos cambios en la riqueza florística y diversidad, contrariamente a lo que se ha reportado comúnmente en sucesiones secundarias. Esto se relaciona a que la vegetación dominante se ha adaptado a adversidades climáticas, limitantes edáficas y la influencia antrópica. Contribuye también la influencia de la abundancia de las plantas anuales, que colonizan rápidamente las parcelas en descanso, alcanzando un alto número de especies desde el inicio de la sucesión, e intervienen oportunamente en descansos antiguos. Además, muchas especies anuales y perennes presentan rangos de tolerancia amplios, permanecen mucho tiempo durante la sucesión. La estructura vertical muestra una predominancia de la vegetación de 0-5 cm en todas las parcelas. Fue posible identificar tendencias sucesionales como la presencia de especies anuales y oportunistas en los primeros años de descanso como Erodium cicutarium, Bromus catharticus, Tagetes multiflora y Tarasa tenella. El aumento de especies de hábito perenne con crecimiento lento y gradual como es la gramínea Stipa ichu, diversas especies de Nassella y arbustos Baccharis incarum y Parastrephia lepidophylla se observan en los descansos hasta los 6 años, después incrementan con una tendencia no lineal hasta los 10 y 20 años, dependiendo de la característica edáfica, humedad, ubicación e historia de la parcela. Según los análisis multivariados, los suelos alcanzan mayor cantidad de nutrientes (C, N, K, Mg y Ca) en 9-22 años de descanso comparado con las parcelas jóvenes.
Palabras claves: Altiplano, Descanso de suelo, Sucesión, Plantas indicadoras.
Abstract
Plant succesional dynamics in fallow plots in the Bolivian Altiplano has been studied, where peasants practice a long, sectorial fallow. The objective was to analyse the successional changes in species diversity, vertical structure of vegetation, abundance of different life forms. The results indicate that there have been little changes in richness and diversity, contrary to what has been reported for secondary successions, possibly due to the adaptation of selected species to the harsh conditions of climate, restricted soil conditions and the anthropogenic influence. This trend can also be related to the great abundance of annual plants, which have the capacity of colonizing rapidly the fallow plots, enabling the existence of a high number of species from the start of succession; these annuals occasionally and opportunistically also settle the older plots. Furthermore, many of the species present a very broad tolerance range, which indicates that after they have colonised a plot, they can stay there for a long time. The vertical structure of vegetation showed a dominance of a herbal layer of 0-5 cm in all plots. It was possible to identify serial trends with annuals like Erodium cicutarium, Tagetes multiflora, Tarasa tenella and Bromus catharticus in the first years of fallow. Till 6 years of fallow we recorded a constant increase of perennial grasses like Stipa ichu and Nassella spp. and some evergreen resinous shrubs; later on we noticed a non lineal increase till 10 to 20 years, depending on soil conditions, humidity, location and history of the plots. With increasing years of fallow the soil fertility (C, N, K, Mg, and Ca) recovered as shown by an analysis multivariable of 9 to 22 year/old plots in comparison to young fallows.
Keywords: Altiplano, Fallow, Succession, Indicator plants
]]>
Introducción
Por la gran extensión que ocupan los sistemas agrícolas con descanso largo son practicados por un gran número de comunidades tanto en el Altiplano boliviano como en otras zonas de los Andes, existiendo extensas superficies bajo este tipo de manejo (Hervé 1994a, Lara & Zapata 2001). El principal objetivo del descanso es la recuperación de la fertilidad de los suelos (Ellenberg 1981, Hervé 1994b, Pestalozzi 2000).
Sin embargo, no hay que ignorar los intensos y frecuentes disturbios representados por el pastoreo y la extracción de leña, un recurso escaso y muy valioso en el Altiplano con una profunda influencia sobre el desarrollo de la vegetación sucesional (Davidson 1993). Durante el periodo de descanso, la sucesión secundaria genera cambios en la estructura y composición de la vegetación, a medida que distintas especies colonizan los terrenos abandonados y van cambiando su abundancia y/o reemplazándose unas a otras (Queiroz et al. 1994).
Resulta prioritario conocer la dinámica de regeneración de la vegetación y de las principales especies involucradas. Este conocimiento no solo es importante para estimar la oferta y calidad del forraje y del recurso leña, sino también para evaluar el impacto del manejo sobre la diversidad de la vegetación. Por otra parte, desde un punto de vista de investigación básica, la sucesión vegetal ha sido escasamente estudiada en ambientes de alta montaña de los Andes, conociéndose relativamente poco sobre los mecanismos implicados y la velocidad de la regeneración con la que ocurre este proceso (Braun 1964, Hervé 1994b, Queiroz et al. 1994, Fontaine 2000, Sarps 2002). Por la gran extensión que ocupan estos sistemas en el Altiplano boliviano. La dinámica del proceso sucesional que genera patrones divergentes o múltiples rutas sucesionales es influenciada por la historia de uso previo de las parcelas, las características del suelo, la disponibilidad de propágulos de las plantas colonizadoras (Queiroz et al. 1994, Sarmiento et al. 2003) y la distribución de las precipitaciones a través del tiempo.
El estudio de la sucesión vegetal ha sido un tema clásico en ecología de comunidades y ha generado un debate sustancial, resultando difícil extraer una teoría general a partir de las tendencias encontradas en ambientes particulares (Odum 1963, Margalef 1968, Pickett & White 1985). Por ejemplo, se ha postulado que la diversidad de la comunidad debe aumentar progresivamente durante las primeras etapas de la sucesión, debido a la llegada progresiva de las especies y disminuir posteriormente en las etapas tardías, debido a la exclusión competitiva (Begon et al. 1995). Sin embargo, hay muchos casos en que la diversidad no sigue este patrón, como por ejemplo el caso del páramo venezolano (Sarmiento et al. 2003). También se ha postulado que uno de los principales mecanismos de la sucesión es el reemplazo de unas especies por otras, pero en ambientes con fuerte estrés, como los ecosistemas árticos y alpinos, se produce una autosucesión, en que son las mismas especies de la comunidad clímax las que progresivamente van colonizando (Svoboda & Henry 1987). Debido a la falta de una teoría unificada y a la poca información disponible sobre el proceso sucesional en ambientes de alta montaña andina, es necesario caracterizar y explicar los patrones sucesionales en estos ambientes.
El objetivo de este trabajo fue evaluar la sucesión secundaria que tiene lugar en el Altiplano central boliviano, en términos de riqueza de especies, abundancia de las formas de vida, estructura vertical de la vegetación y tendencias de las principales especies que intervienen en la sucesión.
Área de estudio
El trabajo fue realizado en la comunidad de Patarani (departamento La Paz, provincia Aroma) del Altiplano central boliviano, a 3.800 m de altitud, ubicada aproximadamente a 110 km hacia el sur de la ciudad de La Paz, cerca de Patacamaya (17°6’S y 68°W). Según datos metereológicos de la estación de Patacamaya obtenidos por SENAMHI (1981-1991) y procesados por Camacho (2001), la precipitación media anual es de 409 mm, con una estación seca entre abril y noviembre y una estación lluviosa de diciembre a marzo; la temperatura media anual es de 10º C. Los suelos son ligeramente ácidos, neutros o ligeramente alcalinos con un contenido muy bajo de materia orgánica y nitrógeno.
]]> El sistema predominante de manejo utilizado es el descanso en sectores, en el cual las tierras de la comunidad están divididas en sectores o aynuqas en que cada familia tiene su parcela. Las aynuqas son cultivadas y luego dejadas en descanso por decisión comunitaria. Cada sector es cultivado generalmente por tres años, iniciándose la rotación con tubérculos (generalmente papa, a veces oca) y continuando con cultivos de granos (quinoa y cebada para grano) y forrajeras (avena y cebada) o viceversa. Las aynuqas en descanso son de uso comunal para el pastoreo de bovinos y sobre todo ovinos. El 77% de las unidades productivas tiene alrededor de 40 ovinos, el 70% cerca de 4 bovinos y el 80% de 1 - 2 equinos (mulas y burros). La ganadería está estimulada por la creciente demanda de carne y leche de vaca, destinadas a ciudades cercanas (Camacho 2001). Además, las aynuqas de varios años de descanso se constituyen en proveedoras de leña de arbustos resinosos de tholas (Parastrephia lepidophylla y Baccharis incarum) que son recolectados por los pobladores.
Métodos
Selección de áreas de muestreo
Las áreas de cultivo y de descanso de la comunidad Patarani se encuentran en la llanura, en el pie de la ladera y en laderas de antiguos andenes. Los comunarios nos facilitaron campos de descanso en las cercanías de los ríos y terrenos de 50-500 m de distancia del pie de las serranías. Allá los comunarios nos destinaron áreas de descanso de 1-50 años, donde seleccionamos parcelas relativamente homogéneas fisionómicamente, desde el punto de vista de la cobertura vegetal y presencia de piedras. Estas parcelas no fueron alambradas y siguieron el ritmo de descanso con pastoreo y casual extracción de leña.
Toma de datos
A partir del año 1999 por cuatro años consecutivos durante la época húmeda se realizaron censos de vegetación en 15 parcelas con las siguientes edades iniciales de descanso: 1 parcela con un año de descanso, 1 de dos años, 2 de tres años, 1 de cuatro años, 1 de cinco años, 1 de seis años, 2 de siete años, 1 de 10 años, 1 de 15 años, 3 de 20 años y 1 de 50 años, obteniendo al final del estudio un total de 60 censos. Las parcelas campesinas tienen tamaños muy variables. Para poder comparar los datos de la vegetación de una parcela a otra definimos el mismo número de puntos de observación que serán repartidos según el mismo protocolo.
Para el muestreo de la vegetación, se utilizó el método del cuadrado puntual (Greig-Smith 1983). En cada parcela se definieron cuatro líneas de 30 m u ocho líneas de 15 m, según el tamaño de la parcela. Se ubicó la mitad de las líneas en un sentido y la otra mitad perpendicularmente, para cubrir lo mejor posible toda el área de la parcela y captar su variabilidad interna. En cada metro de estas líneas se registraron los contactos de las diferentes especies con una varilla graduada de 0.5 cm de diámetro, que era colocada verticalmente. En total se muestrearon 120 puntos por parcela, registrándose la ausencia o presencia de contacto de cada especie en los siguientes intervalos de altura, de acuerdo a las graduaciones de la varilla: 0-5 cm, 5-10 cm y posteriormente en intervalos cada 10 cm. Los muestreos se realizaron durante la estación de lluvias cuando la vegetación estuvo más desarrollada y la mayor parte de las especies se encontraron en floración.
]]> Riqueza y diversidad
La riqueza de especies en cada parcela se calculó como el número total de especies que fueron registradas en los 120 puntos muestreados. La frecuencia de cada especie, que es una medida de su abundancia, se calculó como la suma de todos los contactos de esa especie dividida entre el número total de puntos muestreados. El valor obtenido fue multiplicado por 100 para comodidad en el procesamiento de los datos, obteniéndose la frecuencia absoluta (Matteucci & Colma 1982, Greig-Smith, 1983). La diversidad de cada parcela se calculó utilizando los datos de frecuencia absoluta en el índice de Shannon - Wiener (Magurran 1987) y se los multiplicó por - 1 para obtener valores absolutos:
H' = Índice de diversidad de la parcela
i = Cada una de las especies
n= Número de especies
pi = Frecuencia absoluta total de la parcela representada por la especie i
yi = Frecuencia absoluta de la especie i
Análisis de medidas repetidas
]]> Para verificar si existían diferencias según la edad o año de observación, se llevó a cabo un análisis estadístico de medidas repetidas, agrupando los datos de las parcelas que tenían inicialmente las edades: de 1-4, 5-7, 10-15 y 20-22 años. Según Zar (1999), el análisis de medidas repetidas es adecuado cuando se realizan observaciones sobre un mismo sujeto como son en nuestro caso las parcelas, ya que el mismo conjunto de las mismas fue inventariado cada año. Los años correspondieron a lo que en este tipo de prueba se denomina factor intra-sujeto (within-subject factor). La edad de las parcelas representó el factor entre sujetos (betweensubjects factor).
Formas de vida
Se realizó también un análisis de la variación y distribución porcentual de la frecuencia absoluta total de tres formas de vida: gramíneas, hierbas y arbustos. Además, para cada una de ellas, se calculó el porcentaje del número de especies de hábito anual o perenne en la sucesión.
Estratificación de la vegetación
Con el método cuadrado puntual se analizó también la estructura vertical de la vegetación, determinando la distribución de los contactos registrados entre los diferentes intervalos de altura.
Distribución de las especies en parcelas de diferentes edades de descanso
Para visualizar la secuencia de las especies abundantes a lo largo de la sucesión y establecer la amplitud de su distribución durante los años de descanso, se calculó el promedio ponderado de cada especie y su tolerancia con respecto al tiempo (Jongman et al. 1996).
]]>PP = Promedio ponderado
T= Tolerancia de cada especie
i = Cada una de las parcelas
n= Número de especies
xi = Tiempo de descanso de la parcela i
yi = Frecuencia absoluta de la especie en la parcela i
Con los datos obtenidos se construirá una figura donde el promedio ponderado muestre la secuencia sucesional. Por otra parte, el rango de tolerancia nos indica la amplitud temporal de cada especie, estando las especies de gran tolerancia presentes en parcelas con un rango amplio de edades y las de menor tolerancia, restringidas a periodos determinados de la sucesión, pudiendo estas últimas ser utilizadas como especies indicadoras de edad.
Análisis de suelos
]]> Los análisis fisicoquímicos se realizaron en el Laboratorio de Calidad Ambiental del Instituto de Ecología (Universidad Mayor de San Andrés, en La Paz), según los procedimientos expuestos en Sivila & Angulo (en este número): nitrógeno (N), carbono (C), calcio (Ca), magnesio (Mg), sodio (Na), potasio (K) en el suelo, relación carbono nitrógeno (C/N), conductividad eléctrica (CE), pH y textura. Además, se estimó el porcentaje de pedregosidad en la superficie donde se realizaron los censos de vegetación.
Análisis multivariado de la vegetación y del suelo
Para el análisis multivariado se utilizaron solo los datos de aquellos censos donde también se tomaron datos fisicoquímicos del suelo, quedando en total 19 censos realizados entre 1999 y 2001: Cuatro parcelas de 1-4 años de descanso, cinco de 6-8 años de descanso, seis de 10-17 años de descanso, tres de 20-22 años de descanso y una de 50 años de descanso.
Se relacionaron los censos de vegetación con las características fisicoquímicas del suelo y la edad de descanso mediante el programa estadístico ADE-4 (Fossati et al. 2002). Se usó el análisis factorial de correspondencia (AFC) y el análisis de componentes principales normalizado (ACPn) para seleccionar las variables más explicativas de los censos de vegetación (Jongman et al. 1996, Kent & Coker 2000). De esta forma, se puede interpretar si existe influencia de algunas de estas variables sobre la estructura de la comunidad vegetal.
Con el fin de optimizar la lectura simultánea de los análisis de suelos y de la vegetación, se utilizaron métodos de acoplamiento y co-inercia (Fossati et al. 2002), siendo esto aconsejable cuando se tienen muchas variables y reducido número de muestras (Chessel & Mercier 1993).
El mapa factorial resultante muestra la relación existente entre la estructura de la vegetación y el suelo (Haase 1990, Fossati et al. 2002).
Resultados
Riqueza y diversidad de especies
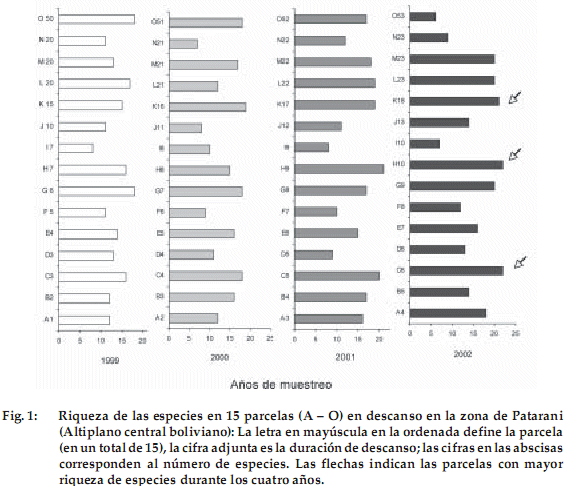
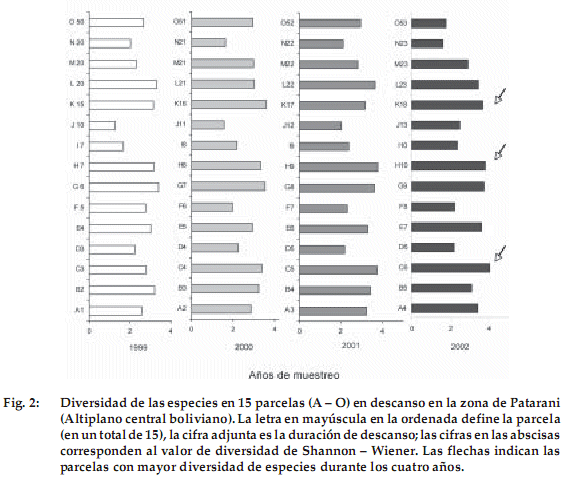
]]> En total fueron registradas 60 especies con el método de cuadrado puntual en los cuatro años de estudio, pertenecientes a 16 familias (Tabla 1). Las familias dominantes fueron las Poaceae con 19 especies, las Asteraceae con 11 y las Fabaceae (Leguminosae) con 6.Los valores de riqueza y diversidad se muestran en las Figuras 1 y 2. Los valores más elevados se presentan en las parcelas de 6 años de descanso (C6) con 22 especies (H’ = 4.0), de 10 años (H10) con 22 (H’ = 3.8) y de 18 (K18) con 21 (H’ = 3.7). El aumento de las especies en estas parcelas es gradual durante los cuatro años de estudio. Estos resultados indican pocas diferencias entre las parcelas de diferentes años de descanso. Sin embargo se observa que de las 15 parcelas estudiadas, 10 tienen tendencia a aumentar su riqueza y diversidad con relación al primer año de estudio (Figuras 1 y 2). De las cinco de las parcelas restantes, tres (con edad inicial de 2, 20 y 50 años de descanso) aumentaron su riqueza y diversidad hasta el año 2001. Dos de las parcelas con edad inicial de 3 y 5 años de descanso disminuyeron en el segundo año de estudio, después se mantuvieron casi constantes en los demás años.
Análisis de medidas repetidas
El análisis de medidas repetidas resume los resultados presentados, ya que no se detectaron diferencias en el número de especies de parcelas de distinta edad (F = 0.081, gl = 3, P = 0.969). Por el contrario, existieron diferencias significativas comparando el número registrado de especies en el primer año de muestreo con los demás años de observación (F = 5.710, gl = 3, P = 0.003). Los años de mayor precipitación promedio anual 2001 (539 mm ) y 2002 (422 mm) presentaron mayor número de especies (15-16 contra 13 en 1999 y 2000). Datos tomados con el método fitosociológico de Braun Blanquet (Ellenberg 1956) en áreas más extensas (19 parcelas) apoyan esta diferencia pequeña con un registro en el año 2001 de un promedio de 25 especies. Esto sugiere que hay germinación de un mayor número de especies en años más lluviosos, como ocurre en otros ecosistemas secos (López 1999).
Formas de vida
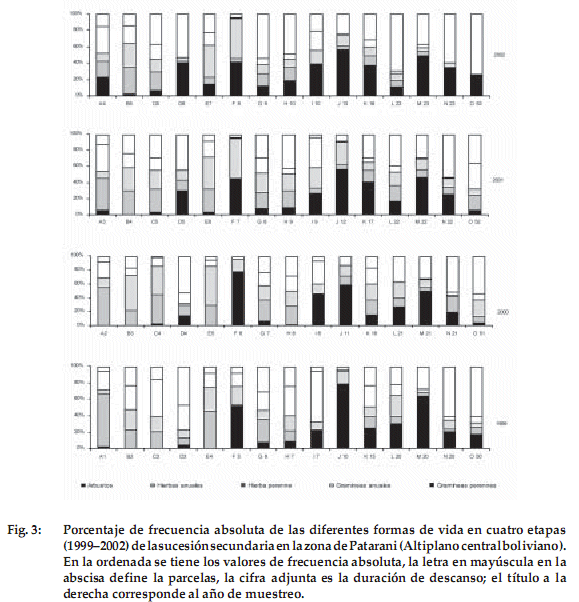
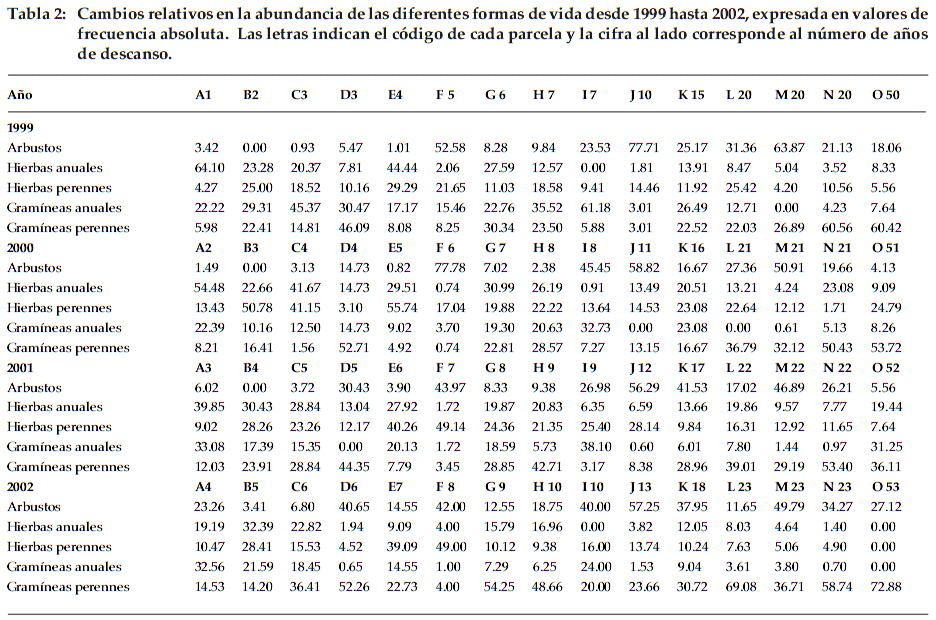
]]> Se presentan los cambios relativos en la abundancia de las diferentes formas de vida y hábitos a lo largo de cuatro años, expresada con valores de frecuencia absoluta (Figura 3 y Tabla 2).A continuación se detallan las tendencias encontradas para cada forma de vida encontrada en la zona de estudio:
Arbustos: En 11 de las 15 parcelas estudiadas, el porcentaje de arbustos aumentó con relación al primer año de estudio. Los valores máximos en todas las parcelas se registraron en los años 1999 en una parcela de 10 años de descanso (J10 con 78%), en otra de 20 años (M20 con 64%) y en el año 2000 en la parcela de 6 años (F6 con 78%). En los siguientes años de descanso, no se alcanzaron valores por encima de 60%. Si observamos los datos sincrónicos de desarrollo arbustivo, se nota que solamente existe en las parcelas A1 hasta E4 un aumento gradual en los cuatro años de estudio, de acuerdo al potencial base de plántulas. Al inicio de nuestro estudio, por ejemplo D3-D6 aumentó de 5-41%. En las parcelas de mayor edad por encima de 5-6 años no hay una tendencia homogénea, algunas suben por un año y en el otro bajan. Las parcelas M20-M23, J10-J13 y F5-F7 muestran porcentajes por encima de 40%, en cambio otras parcelas como H7-H10 y G6-G9 están por debajo de 20%. Probablemente hayan algunos factores edáficos, hídricos y posiblemente antropogénicos -no medidos en este estudio - que expliquen estos resultados.
Hierbas anuales: En 10 de las 15 parcelas estudiadas, las hierbas anuales disminuyen su porcentaje con relación al primer año de la toma de datos. Las hierbas anuales presentan su máximo al inicio de la sucesión, en la parcela A1 con un porcentaje de 64% y disminuyen en general en las parcelas mayores de cuatro años de descanso, alcanzando en algunas parcelas de N20 y O53 valores con 1.4% a 0%, respectivamente. Las especies anuales como Erodium cicutarium, Tagetes multiflora y Tarasa tenella no muestran un ritmo anual, sino se portan como efemérofitos, ya que aparecen después de unas lluvias y desaparecen frente a condiciones secas; otras especies a veces actúan como bianuales.
Hierbas perennes: Las hierbas perennes no muestran una tendencia clara, alcanzan sus valores máximos en las parcelas de 5 años (E5 con 56%) y 8 años (F8 con 49%), disminuyendo, aumentando o manteniéndose a lo largo de la sucesión con valores alrededor del 20%. Solamente la parcela de 53 años de descanso carece de registros.
Gramíneas anuales: Las gramíneas anuales predominan por lo general en parcelas con pocos años de descanso. Además en 11 de las 15 parcelas estudiadas, el porcentaje de gramíneas anuales disminuye con los años de descanso. Es notorio el valor excepcionalmente alto del antiguo barbecho de 52 años (O52) con 31%; esto posiblemente se explique por la elevada precipitación en el año 2001. En el próximo año, no encontramos ninguna planta anual, ni gramínea ni hierba en esta parcela.
Gramíneas perennes: Las gramíneas perennes tienden a aumentar con los años de descanso, por ejemplo en las parcela de 20 años aumenta de 22% hasta 69% (L20-L23). En 13 de las 15 parcelas estudiadas su porcentaje aumenta alcanzando en la parcela de 53 años el valor de 73% por la dominancia de Aristida asplundii, especie poco consumida por el ganado. En el año 2002 se nota un aumento de las gramíneas perennes en todas las parcelas (menos B5) respecto al anterior año, posiblemente por la acumulación de humedad en el suelo derivada de las precipitaciones excepcionalmente altas en el año 2001.
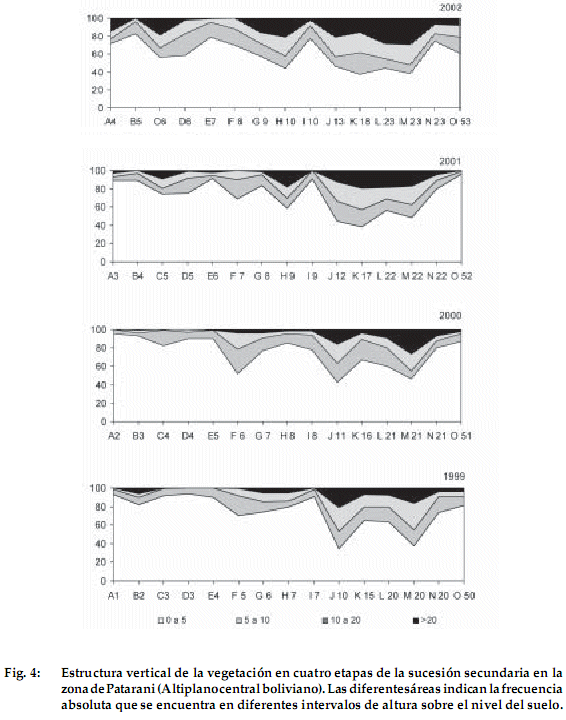
]]> Estructura vertical de la vegetación
La Figura 4 muestra los cambios sucesionales en la estructura vertical de la vegetación en los cuatro años de estudio. Independientemente de la etapa sucesional, la mayor parte de la vegetación se encuentra en un rango de 0 a 5 cm de altura y solo una parte muy reducida se encuentra por encima de los 10 cm. Es notorio un aumento del tamaño de la vegetación a medida que transcurre el tiempo de descanso, para llegar a un porcentaje mayor de vegetación por encima de los 20 cm en barbechos de 9-23 años de descanso. La parcela de 50 años de descanso muestra una vegetación predominante con especies de 0-5 cm con pocos ejemplares arriba de 20 cm. Por lo general, las especies que alcanzan mayor altura son gramíneas y arbustos.
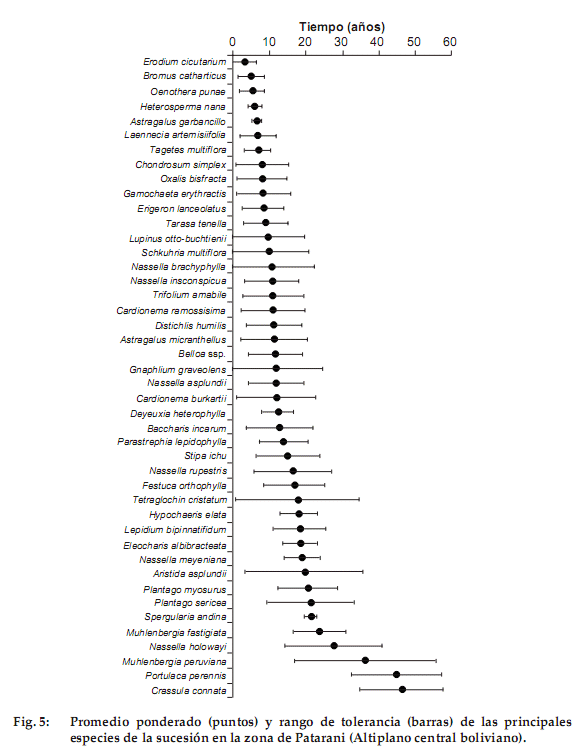
Tendencias sucesionales de las especies
La Figura 5 indica el promedio ponderado y la tolerancia para las 44 especies más abundantes. Las especies que se encuentran en parcelas con menor edad de descanso sonErodium cicutarium (3.3 ± 3 años) y la gramínea bianual Bromus catharticus (5 ± 3.5 años). Estas especies presentan además un rango de tolerancia menor, que nos indica que son especies casi exclusivas de parcelas con edades de descanso recientes. Entre 5 y 10 años de descanso aparecen la leguminosa perenne Astragalus garbancillo (6.6 ± 8 años), Laennecia artemisiifolia (6.9 ± 5.1 años), Tagetes multiflora (7 ± 3.6 años), Oxalis bisfracta (8.1 ± 6.8 años), Chondrosum simplex, sinónimo de Bouteloua simplex (8.1 ± 7.2 años) y Erigeron lanceolatus (8.4 ± 5.8 años). De estas especies las que presentan un rango de tolerancia menor son Tagetes multiflora y Astragalus garbancillo. Algunas especies que tienen su máxima abundancia en etapas posteriores son Nassella inconspicua (10.8 ± 7.3 años), Trifolium amabile (10.8 ± 8.3 años), Distichlis humilis (11.2 ± 7.4 años), Deyeuxia heterophylla (12.4 ± 4.4 años) y Parastrephia lepidophylla (13.8 ± 6 años). De todo este grupo, las especies que presentan una tolerancia menor son Deyeuxia heterophylla y Parastrephia lepidophylla. De 15 a 20 años encontramos a Stipa ichu (15.2 ± 8.6 años), Festuca orthophylla (16.9 ± 8.5 años) y la ciperácea Eleocharis albibracteata (18.5 ±4.7), que es la especie con menor rango de tolerancia en este grupo. Finalmente, las especies que aparecen en los últimos años de descanso sonNassella meyeniana (19.1 ± 4.8 años), Plantago myosurus (20.4 ± 8.5 años), P. sericea (21.4 ± 11.9 años), Muhlenbergia fastigiata (23.7 ± 7.1 años), Portulaca perennis (44.7 ± 12.4 años) y Crassula connata (46.3 ± 11.2 años). Las especies con menor tolerancia de este grupo sonNassella meyenianayMuhlenbergia fastigiata. El análisis de Figura 5 nos muestra la sucesión como un continuo, donde las especies tienen rangos de tolerancia amplios, lo que indica que están presentes durante gran parte de la sucesión y dificulta la definición de etapas seriales claras.
Análisis multivariado del suelo y de la vegetación
Parámetros fisicoquímicos del suelo
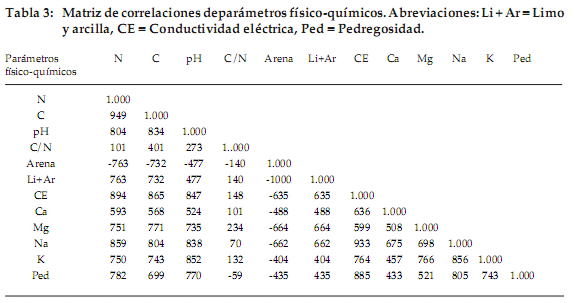
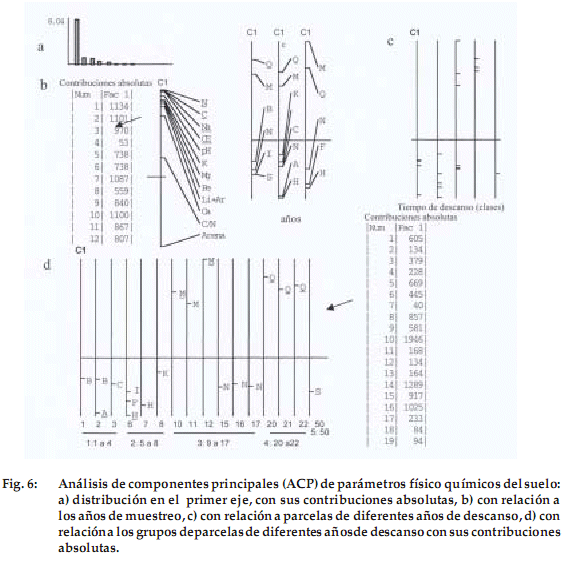
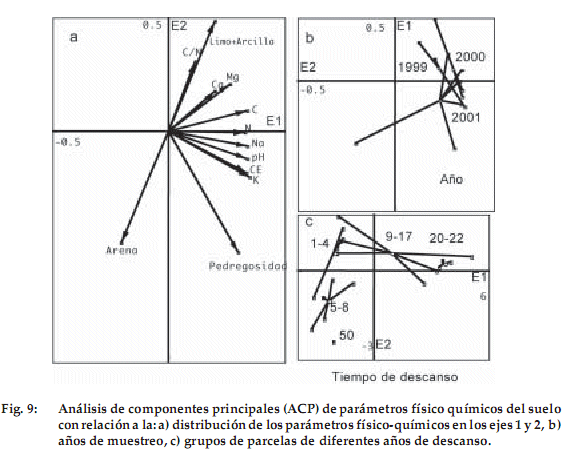
]]> En los parámetros del suelo (Tabla 3), las variables que mejor se correlacionan con el nitrógeno son el carbono (r = 0.949), el pH (r = 0.804) y la conductibilidad eléctrica (r = 0.894). Por lo tanto, en suelos con alto contenido de nitrógeno existe elevado contenido de carbono, un pH adecuado y una fuerte conductibilidad eléctrica. Las variables que mejor se correlacionan con la pedregosidad son la conductibilidad eléctrica (r = 0.885), el sodio (r = 0.805) y el nitrógeno (r = 0.782). La variable limo–arcilla tiene correlación positiva con el nitrógeno (r = 0.763) y carbono (r = 0.732). La arena tiene correlación negativa con nitrógeno (r = -0.763) y carbono (r = -0.732), por que los suelos donde la tasa de limo y arcilla es mejor son más ricos en nitrógeno y carbono que los suelos arenosos.El primer eje del ACPn es nítidamente el más importante (Fig. 6a). Las variables que contribuyen a este eje son el nitrógeno, carbono, calcio y magnesio (contribuciones absolutas entre 0.1134 y 0.1087). Opuestamente a estas variables, el porcentaje de arena no tiene una contribución significativa (Fig. 6b). No existe una relación notable entre los años de muestreo y los parámetros de suelo durante los tres años de estudio (Fig. 6c).
Los análisis físico-químicos de suelo se diferencian claramente en cuatro grupos, según los años de descanso: (1): 1-4, (2): 5-8, (3): 9-17, (4): 20-22 (Fig. 6c y 6d). Las parcelas con mayores valores de contribución son las que se encuentran en el tercer grupo de edad, seguidas por el cuarto grupo. El primer y segundo grupo y por otro lado las parcelas con código «N» se separan por presentar textura gruesa al igual que la parcela aislada de cincuenta años.
Existe entonces un cambio en los suelos de las parcelas con más de nueve años de descanso, presentando mayor cantidad de nutrientes y sales en relación a las más jóvenes.
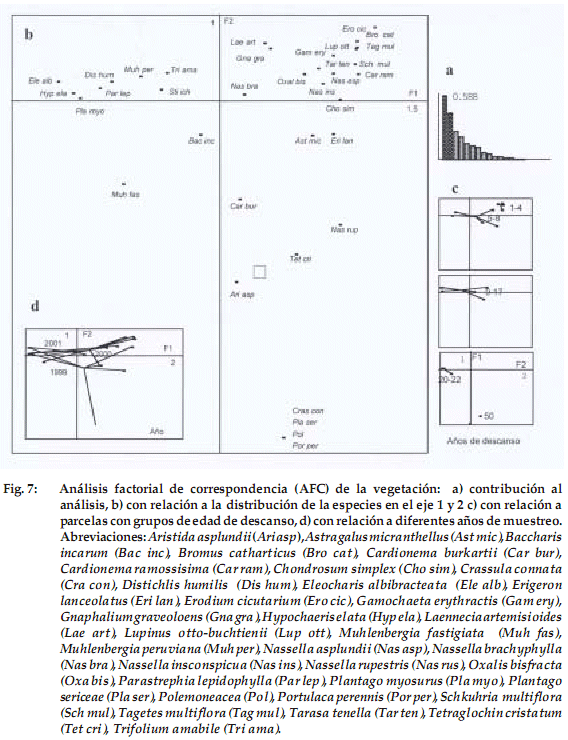
Vegetación y flora
La distribución de los censos de la vegetación en el AFC deja ver una tipología de la vegetación según el tiempo de descanso (Fig. 7 a). Las parcelas de los cuatro grupos de edad (1-22 años) se encuentran distribuidas en el eje 1, separándose de la parcela de 50 años que ésta más asociada al eje 2. Además el grupo de parcelas de 1-4 años de edad de descanso se distancia de los grupos de 5-8 y 9-17 (las mismas que entre ellas no presentan diferencia) y a su vez estas últimas se alejan de las del grupo de edad de 20 a 22 (Fig. 7c).
]]> Si relacionamos los grupos de edades de descanso con las especies (Fig. 7b y 7c), basándonos en las contribuciones absolutas más fuertes, vemos que las especies predominantes en el primer grupo de edad, que en su mayoría contribuyen al eje 1 son: Erodium cicutarium,Bromus catharticus, Schkuhria multiflora, Oxalis bisfracta, Nassella insconspicua y Lupinus otto-buchtienii. En el segundo y tercer grupo de edad tenemos a Parastrephia lepidophylla, Trifolium amabile y Baccharis incarum. En el cuarto grupo tenemos a Eleocharis albibracteata, Hypochaeris elata y Plantago myosurus. Finalmente en la parcela de 50 años, que contribuye mayormente al eje 2, tenemos a Aristida asplundii, Tetraglochin cristatum, Portulaca perennis y Crassula connata. En la Figura 7d notamos cómo la vegetación se agrupa con relación a los diferentes años de estudio, por un lado más cerca del eje 1 los años 2000 y 2001 y el año 1999 que contribuye más al eje 2.
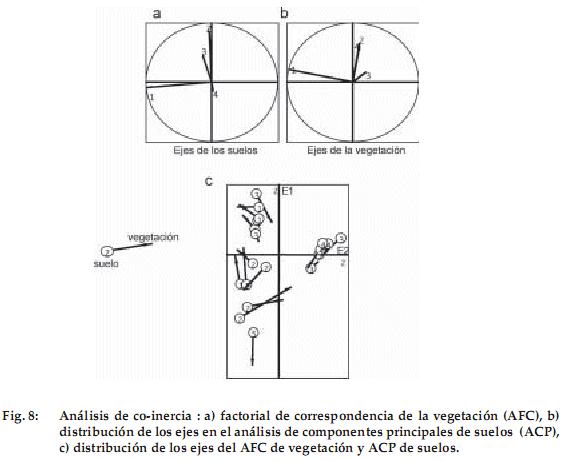
Análisis de co-inercia
En las Figuras 8a y 8b los cuatro ejes de los ACPn (suelos) y el AFC (vegetación) muestran una relación entre los ejes 1 y 2 de co-inercia. La co-inercia de los diferentes grupos de edades tanto en vegetación (representada por la flecha) como en suelos (representados por un círculo con el año de grupo) se nota en la Figura 8c. La longitud de la flecha indica el nivel de relación entre los parámetros físico-químicos del suelo y la vegetación: mientras más larga sea, menor relación se da. Los grupos 3 y 4 presentan una buena relación entre la vegetación y los factores físico–químicos del suelo y el grupo 2 tiene una menor relación.
Las variables de textura se encuentran distribuidas a lo largo del eje 2 (Fig. 9a). Las texturas finas (limo–arcilla) se sitúan en la parte positiva del eje y las texturas más gruesas (arena y pedregosa) en la parte negativa del eje. Comparando esta relación con la Figura 9c, vemos que las parcelas con edad de descanso de 1-8 años (primer y segundo grupos) son de textura arenosa, mientras que las parcelas de 9-22 años son de textura fina limo arcillosa y la parcela de 50 años de descanso es pedregosa.
Con relación a las variables químicas del suelo (Figura 9a) se distribuyen en la parte positiva del eje 1. Sobreponiendo esta gráfica con la Figura 9c, podemos interpretar que las parcelas de 1-8 años de descanso no están siendo influidas por la disponibilidad de nutrientes, mientras que la vegetación predominante en las parcelas de 9-22 años de descanso tiene una relación opuesta. En la parte de inferior de la gráfica se encuentra la parcela de 50 años de edad de descanso, pero al estar aislada no resulta muy claro interpretar su relación con las características químicas.
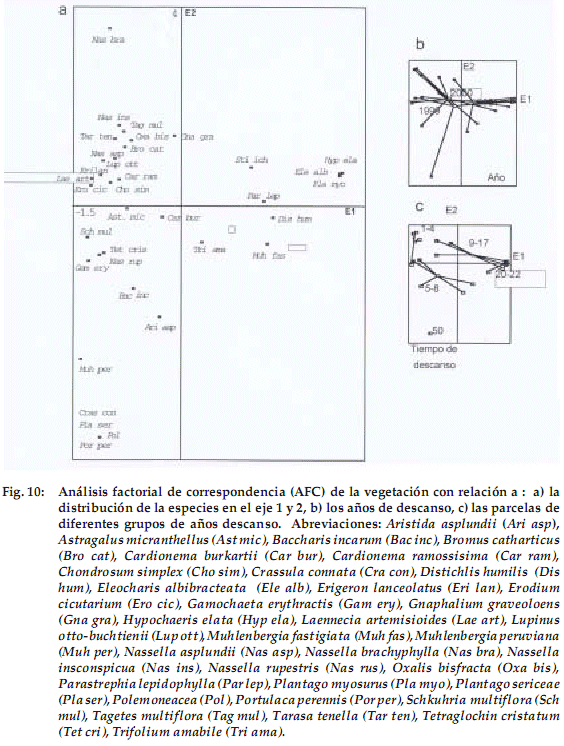
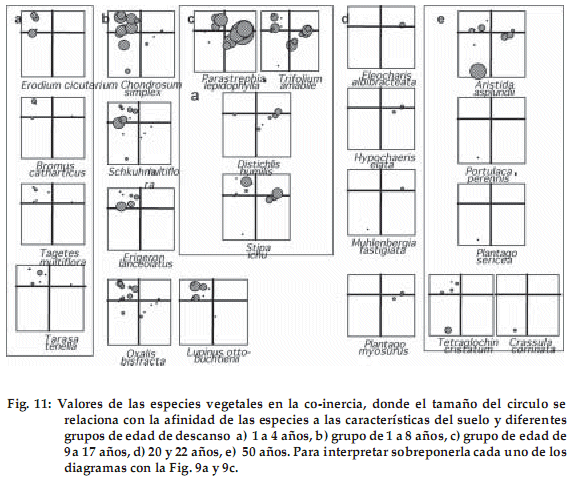
Al relacionar los parámetros físico-químicos del suelo con las especies identificadas en los diferentes grupos de edad (sobreponiendo las Fig. 9a y 10a), podemos observar que las especies que se desarrollan en suelos con textura gruesa (arenosa) y con menor disponibilidad de nutrientes son: Chondrosum simplex, Erodium cicutarium, Tarasa tenella, Tagetes multiflora, Erigeron lanceolatus, Bromus catharticus, Oxalis bisfracta,Nassella asplundii,Schkhuhria multiflora, Baccharis incarum, y Astragalus micranthellus.
]]> Por último, el grupo de parcelas de mayor edad de descanso, en que influye además la pedregosidad, presenta a: Aristida asplundii, Tetraglochin cristatum, Portulaca perennis y Crassula connata.Por el contrario, aquellas especies que crecen en suelos con textura fina asociada con una mayor concentración de nutrientes y una elevada concentración de sales alcalinas son: Parastrephia lepidophylla, Distichlis humilis, Trifolium amabile, Stipa ichu, Muhlenbergia fastigiata, Hypochaeris elata, Eleocharis albibracteata y Plantago myosurus.
La Figura 11 respalda la relación entre la afinidad de las especies vegetales en la coinercia, con los diferentes grupos de edad y las características del suelo (sobreponiendo cada uno de los diagramas de la figura 11 con las figuras 9a y 9c).
Discusión
La dinámica de la vegetación del ecosistema semi-árido de altura va depender en primer lugar de factores limitantes climáticos con régimen de precipitación unimodal con baja precipitación estacional, temperaturas diurnas muy variables con alta radiación durante el día, heladas nocturnas, fuertes vientos; en segundo lugar de factores limitantes edafológicos, y en tercer lugar de la historia de la influencia antrópica (Ruthsatz 1983). En el caso del Altiplano boliviano, esta influencia se remonta a 4000 ó 3000 a.c. según datos palinológicos (Markgraf 1985, Baied et al. 1993) y según Kessler & Driesch (1993) desde hace 10.000 años, intensificándose durante la Colonia de España, utilizando plantas para combustible, además de la introducción de vacas, ovejas, entre otros y aplicando prácticas agrícolas que causaron la degradación de vastas regiones de los Andes (Posnansky 1983, Ruthsatz 1983, Beck 1985).
Por las características de los factores que están influenciando en la dinámica de vegetación en este ecosistema de altura según los resultados obtenidos en el estudio realizado, corresponderían al modelo denominado por Grime (2001) como proclimax. El proclimax es el resultado de sucesivas interrupciones del proceso sucesional normal, donde las especies que dominan compiten bajo constantes presiones de disturbio como el pastoreo y extracción de leña y bajo factores climáticos adversos, donde la representación de algunas especies que juegan un determinado rol puede variar del caso de una sucesión normal (Grime 2001) y donde solo dominan las especies con mayor adaptabilidad (Lorini 1994, Garcia 1997).
Otro aspecto que dificulta el análisis de la sucesión en los ecosistemas de altura es el no disponer de un paisaje natural prístino o poco alterado con que se puedan comparar los resultados de las parcelas en descanso. Esta vegetación natural constituiría un punto de referencia o patrón que nos permitiría discutir si todas las especies logran colonizar las parcelas en descanso, diferenciar las especies que solo están presentes en la sucesión de aquellas que son exclusivas de la vegetación natural y evaluar el tiempo requerido para una regeneración total del ecosistema. Estudios anteriores en las comunidades altiplánicas de Huaraco y Ulla Ulla no lograron este fin por la destrucción del alambrado antes de llegar a un estado de vegetación semi-natural.
]]> Riqueza y diversidad
La presión a la que se ve sometida la vegetación en el área de influencia altoandina se refleja en el reducido número de especies (60), registrado por el método del cuadrado puntual en los 60 censos de vegetación. Esta reducida diversidad se explica por varias razones, mencionadas arriba, además de las características geomorfológicas. Las parcelas se encuentran ubicadas en lugares planos, alejados de las laderas donde existe mayor diversidad de especies, donde son lugares más resguardados de los vientos fríos, además al ser suelos más rocosos existe menor influencia de pastoreo y de cultivos. Si las parcelas de estudio estuviesen en las laderas, probablemente la flora circundante tendría mayor incidencia en la composición florística de parcelas en descanso, ya que las plantas que crecen en las laderas podrían actuar como un sumidero de semillas lo cual podría influir para que aumente la diversidad (Shmida & Wilson 1985, citado por Clark 2002). La diferencia en la riqueza de especies atribuida a la ubicación geomorfológica también se manifiesta en dos trabajos en la comunidad de Titicani-Tacaca realizados en parcelas de laderas en descanso con un total de 81 especies (Chumacero 2003); y el otro en las colinas onduladas y parte de la serranía de vegetación semi-natural con 196 especies (Villavicencio 1990). Si comparamos nuestros resultados con los obtenidos en los anteriores estudios mencionados, vemos que el número de especies en la comunidad de Patarani es más reducido. Esta diferencia también se debe a la diferencia de humedad que existe entre ambas comunidades, así en el Altiplano central (Patarani) el promedio de precipitación es de 409 mm (1981-1991), mientras que en el Altiplano norte (Titicani - Tacaca) es 719 mm (1957-1988).
Al realizar un análisis independiente de cada una de las parcelas en el transcurso de cuatro años de estudio, la mayoría de las parcelas presentan un aumento en su riqueza y diversidad con relación al primer año de estudio. Sin embargo, no son evidentes las diferencias entre edades de descanso, es decir, que una parcela con más años de descanso, no tiene mayor número de especies que parcelas con menos años de descanso. A diferencia de un estudio similar sobre los cambios en la vegetación en parcelas de descanso y realizado en el Páramo de Gavidia en Venezuela (3.200-3.800 m, 1.200 mm de precipitaciones), donde se aplicó el mismo método del cuadrado puntual en 18 parcelas en descanso (de 10 meses a 20 años de descanso) sus resultados muestran un crecimiento lineal de la riqueza y la diversidad de especies con relación a los años de descanso (Pérez & Arriaga 2000). Estos resultados están dentro de lo que se esperaría en el curso de una sucesión donde la diversidad aumenta en el tiempo, a medida que más especies colonizan o bien aumenta hasta un máximo en etapas intermedias y luego disminuye (Huston 1994). La diferencia entre ambos resultados puede darse por un lado a las características del ecosistema de altura sometido a uso continuo, también dentro de la época de descanso sigue el pastoreo y la extracción parcial de leña. La poca variabilidad en el número de especies y diversidad en el análisis de las parcelas en diferentes años de descanso, pudiera estar relacionada con la presencia de muchas especies con buenas características de colonizadoras, con una producción abundante de semillas y dispersión eficiente. Esto ayudaría a que el número de especies sea alto desde los primeros años de descanso.
Cabe resaltar que una gran parte de las especies presentes en esta sucesión son anuales, lo que obedece a una estrategia de adaptación a la escasa y muy estacional precipitación. Al ser anuales de zonas semi-áridas, estas especies invierten gran parte de su energía en estructuras reproductivas y bancos de semillas que les permiten recolonizar el espacio año tras año y esto explicaría, al menos en parte, por qué la riqueza de especies permanece constante en la sucesión. Aparte de la estrategia del banco persistente de semillas, las plantas anuales suelen tener dos estrategias de respuesta al agua: 1) La producción de una fracción de germinación constante aunque pequeña de año a año; 2) una respuesta más bien de tipo predictivo, que les permite regular la proporción de su banco de semillas que germinaría en un año, en función a las condiciones más o menos buenas que la planta adulta podría encontrar (Cohen 1966, 1967; Venable & Lawler 1980). Sin embargo, la respuesta a un nivel más fino suele ser más específica por cada especie.
Formas de vida y estructura de la vegetación
El análisis de datos de las formas de vida y estructura de la vegetación llama la atención en el comportamiento de los arbustos, que aumentan en los primeros cuatro años de la sucesión, llegando a su máximo en parcelas de seis años de descanso y en otras en 10 ó 20 años. Teóricamente, la parcela de 50 años debería tener más arbustos, que no es el caso. Esta reducción de los arbustos se observa también en algunas parcelas a partir de seis años de descanso (Fig. 3). Explicaciones generales y reales siempre indican el saqueo de leña, pero aparte existe además un fuerte factor climático y edáfico. Claramente los arbustos y gramíneas perennes han podido aprovechar de las precipitaciones elevadas en el año 2001, que se reflejan en un aumento general de su frecuencia absoluta y además en un aumento general de la estructura vertical en 2002 (Figuras 3 y 4). Los suelos limo-arcillosos mantienen mejor la humedad y presentan mayor fertilidad, que se manifiesta en el mejor desarrollo de las gramíneas perennes y arbustos.
Estas tendencias de aumento de los arbustos también se encontraron en la comunidad de San José de Llanga –Altiplano central - donde Ramos (1995) citado en Queiroz et al. (2001) encontró que los cambios que ocurren en plantas perennes en campos en descanso tienden a estabilizarse después de cinco años en descanso, pero que la cobertura de arbustos deParastrephia lepidophylla continúa aumentando hasta llegar a su pico entre los 10-20 años de descanso. Además en otro estudio de Queiroz et al. (1994) exclusivo sobre P. lepidophylla se conoció que su cobertura y densidad aumenta hasta los 8-12 años, decreciendo en periodos más largos de descanso. Las causas de esta disminución no se estudiaron a fondo, en que seguramente influyen la mortalidad debida a la avanzada edad de los arbustos y la extracción de leña de Parastrephia lepidophylla por la gente (Barrera 1994, citado en Queiroz et al. 2001, Camacho 2001).
Con respecto a la abundancia de especies de hábito anual y perenne, los resultados muestran (aunque no de manera lineal en todas las parcelas) que existe una tendencia a que exista mayor número de hierbas y gramíneas anuales en los primeros años, que luego van disminuyendo a lo largo del tiempo. Lo contrario ocurre con respecto a hierbas y gramíneas perennes que aumentan conforme se incrementa la edad del barbecho, no dándose esta tendencia en forma lineal (Fig. 4), lo cual se explica por las diferencias específicas que existen entre las parcelas. Similar tendencia presenta Ramos (1995 citado en Queiroz et al. 2001) que indica que en parcelas de descanso de San José de Llanga con seis o más años existen mayores pastos perennes como Festuca orthophylla, mientras que declina la cobertura de especies anuales de hierbas y gramíneas.
Vale tomar en cuenta que el uso de las palabras anual y perenne proviene de lugares templados del hemisferio norte, con un ritmo marcadamente estacional, con relativa humedad durante todo el año. Bajo las condiciones semiáridas de las montañas tropicales no existe un ritmo fijo, ni para plantas herbáceas ni para leñosas. Su desarrollo sigue los principios del oportunismo: también en plena época de invierno aparecen hierbas anuales como Erodium cicutarium y florecen los arbustos siempreverdes de Baccharis y Parastrephia.
]]> Según Grime (2001) en hábitats improductivos, las hierbas perennes y arbustos están sometidos a un importante estrés de competitividad con especies anuales y ruderales en etapas tempranas de la sucesión. El lento crecimiento de plantas perennes parece estar relacionado con la limitación de nutrientes. En regiones semiáridas con deficiencia de nutrientes en el suelo, el clímax de la vegetación a menudo está compuesto de arbustos escleromorfos y pequeños árboles con crecimiento lento; esto no es inusual y proporciona una cobertura en que especies leñosas discontinuas permiten la presencia de hierbas bajas tolerantes al estrés (Siccama et al. 1970, Arno et al. 1972, Inouye et al. 1987 citados en Grime 2001).En el análisis de la forma de vida y estructura de la vegetación observamos que la sucesión secundaria es relativamente lenta y tiende a moverse directamente desde la competencia de cobertura por especies anuales hasta incluir el lento crecimiento de arbustos leñosos como Parastrephia lepidophylla, Baccharis incarum, Tetraglochin cristatum y/o matas de gramíneas de Nassella brachyphylla, N. inconspicua y Stipa ichu, cuyos individuos están distribuidos en forma dispersa y bajo su protección se encuentran hierbas anuales y/o perennes, dependiendo de la edad de descanso, por ejemplo Tagetes multiflora y Oxalis bisfracta.
Nuestros resultados concuerdan con la dominancia de hierbas y gramíneas con un rango de altura de 0-5 cm, dominando el tipo de crecimiento en roseta y en cojines que es una adaptación al clima frío y al estrés hídrico (Grime 2001, Sarmiento et al. 2003). Unos ejemplos son Oxalis bisfracta cuyas raíces son muy ramificadas y alcanzan profundidades en el suelo mayor a 80 cm y las gruesas y tuberosas raíces de Trifolium amabile. Estas adaptaciones de la vegetación también responden al estrés causado por el pastoreo, ya que se puede observar que esta acción ejerce una presión selectiva de las especies, permaneciendo las oportunistas y aquellas resistentes al ramoneo y pisoteo del ganado (Connell 1978, Garcia 1997, Chumacero 2003).
Estos resultados también pueden ser analizados desde el punto de vista de la oferta y calidad del forraje. Un primer análisis nos revela una disminución progresiva de gramíneas y herbáceas a medida que aumentan los arbustos (hasta una edad media), lo que incidiría en una oferta menor de forraje a medida que avanza la sucesión (Genin & Fernández 1994). Esto indica que en las parcelas de descanso jóvenes hay abundante oferta de forraje (Bromus catharticus, Nassella spp., Trifolium amabile, Lupinus otto-buchtienii), especialmente en la época húmeda, que es cuando se observa ganado ovino. Si bien en las parcelas de descanso mayor existen muchas gramíneas, no predominan las especies palatables sino las especies poco palatables como Aristida asplundii, Muhlenbergia peruviana y Stipa ichu.
Tendencias sucesionales de las especies
En el análisis de promedio ponderado y la tolerancia de las especies (Fig. 5), los rangos de tolerancia de la mayoría de las especies son amplios, lo que indica que se mantienen durante mucho tiempo una vez que colonizan, pudiendo aumentar o disminuir en abundancia pero sin desaparecer completamente. Esta es otra característica de esta sucesión, donde el reemplazo de las especies a lo largo del tiempo parece ser menos importante que sus cambios en abundancia. Un patrón similar fue observado en sucesiones secundarias en el páramo venezolano e interpretado como una consecuencia de una exclusión competitiva incompleta en un ambiente donde la adaptación al estrés es más importante que la competencia entre especies (Sarmiento et al. 2003).
Sin embargo, haciendo un análisis más fino por especie, se ha podido identificar tendencias sucesionales definidas lo cual proporciona información valiosa sobre la amplitud ecológica de cada una de ellas durante el tiempo de descanso. Las especies Erodium cicutarium y Bromus catharticus son casi exclusivas de edades recientes de descanso (3-5 años). Después aparecen especies que se mantienen entre 5 y 10 años de descanso con un rango más amplio de tolerancia en barbechos como Tagetes multiflora, Oxalis bisfracta yChondrosum simplex. Estos resultados coinciden con los encontrados por Chumacero (2003), quien realizó análisis similar en parcelas con un rango de edad de 1-8 años de descanso, además también con los trabajos realizados por Genin et al. (1994). Entre los 12-13 años predominan Deyeuxia heterophylla y Parastrephia lepidophylla, esta ultima especie coincide con resultados encontrados por Queiroz et al. (1994) y Ramos (1995). En tanto el promedio ponderado con menor tolerancia en las edades de descanso entre 15 y 20 años se encuentran Stipa ichu, Festuca orthophylla y Eleocharis albibracteata. Finalmente, las especies que se encuentran con menor rango de tolerancia en la parcela de 50 años son Portulaca perennis, Crassula connata, Muhlenbergia peruviana y M. fastigiata.
Sin embargo, en estos resultados nos llama la atención que Muhlenbergia fastigiata se encuentre dentro del último grupo de edad, ya que en el estudio realizado por Chumacero (2003), esta especie se encuentra en años de descanso de 3-5 y 7-5 años. Esta diferencia puede deberse a que en la comunidad Titicani-Tacaca los suelos presentan mayor humedad que en Patarani, ya que según datos de Pestalozzi (1998) y Navarro (2002) generalmente se encuentra creciendo en lugares húmedos y levemente salinos. Igualmente, Eleocharis albibracteata que en nuestros resultados aparece en parcelas de 15 a 20 años de descanso, fue encontrada por Pestalozzi (1998) creciendo en laderas húmedas y levemente salinas.
También debemos tomar en cuenta que la agrupación de estas especies está basada en el seguimiento de una sola parcela, que en el primer año de estudio contaba con 50 años de descanso. Esta parcela tiene la particularidad de tener un suelo altamente pedregoso, que quizás sea la razón por la cual no ha vuelto a ser cultivada en tan largo periodo de descanso. Esta característica del suelo puede ser relacionada con la presencia de pequeñas especies suculentas como Crassula connata y Portulaca perennis, que buscan suelos cálidos y una protección a lado de piedras, igualmente como la gramínea anualMuhlenbergia peruviana.
]]>Análisis multivariado de la vegetación y de suelo
El análisis multivariado nos ayudó a comprender mejor los patrones de la distribución vegetacional en la sucesión, confirmando en la mayoría de los casos los resultados obtenidos en análisis parciales y esclareciendo los resultados obtenidos al añadir la variable suelo para la interpretación de la vegetación (Grime 2001, Clark 2002). Con este aporte se explica mejor que algunas de las tendencias encontradas no solamente obedecen a la edad de descanso, sino a las características del suelo, como es el caso de Aristida asplundii que domina y se adapta mejor en suelos pedregosos (Fig. 10).
Los resultados muestran (Fig. 9a y 9c) que las parcelas con menor edad de descanso se agrupan en suelos arenosos, por lo tanto, con menos disponibilidad. Obviamente no todas las parcelas jóvenes en descanso presentan suelos arenosos, ya que puede ser solo una coincidencia de la distribución de las parcelas analizadas, pero si es evidente que parcelas jóvenes generalmente muestren suelos con pocos nutrientes. Esta es la razón por la cual van a predominar especies oportunistas como Erodium cicutarium, Bromus catharticus, Tarasa tenella, Tagetes multiflora y Chondrosum simplex (Fig. 9 y 10) que por lo general tienen hábito anual, cuya estrategia de competencia requiere poca cantidad de nutrientes (Grime 2001, Clark 2002).
Es también deducible que en estas parcelas con menor disponibilidad de nutrientes, el proceso de colonización de arbustos y especies perennes sea más lento, como lo registrado en las planicies arenosas en Cedar Creek, Minnesota (Inouye et al. 1987, citado en Grime 2001). Esta lenta colonización de especies perennes también se observa en llanuras aluviales arenosas del Altiplano boliviano, en resultados obtenidos en el estudio del matorral deParastrephia lepidophylla en San José de Llanga (Queiros 1994). A pesar de su lento desarrollo, esta especie llega a dominar suelos fluviolacustres, ya sea en largos periodos de descanso o en vegetación semi-natural (Ruthsatz 1977, Seibert 1983, Navarro 2002).
En el caso de parcelas de descanso de 9 a 22 años de descanso nuestros resultados del análisis multivariado (Fig. 9 y 10) muestran una tendencia de aumento de los elementos como el C, N, K, Mg y Ca con relación a las parcelas más jóvenes, observaciones que coinciden con Hervé & Sivila (1997), Pestalozzi (1998), Queiroz et al. (2001) y Chumacero (2003).
Se observa un aumento de carbono en parcelas de 9 a 12 años de descanso. Su acumulación requiere de largos periodos de descanso, debido a la escasa acumulación de materia orgánica a causa del pastoreo extensivo y la baja cubierta vegetal de la puna seca (Hervé & Sivila 1997). También Chumacero (2003) registra un aumento de carbono de 0.55-1.10 % en sus parcelas en descanso a lo largo de siete años de descanso. Hervé (1994b) señala que este elemento aumenta hacia los 5-8 años para luego estabilizarse en largos periodos de descanso en la comunidad de Pumani (Altiplano central). El nitrógeno y el potasio también indican un aumento en nuestras parcelas; Chumacero (2003) registra un incremento del 13% de nitrógeno después de siete años de descanso.
Uno de los resultados que llama la atención (Fig. 9a, 9c) es el incremento en la concentración de minerales de Ca, Mg y Na en las parcelas agrupadas de 9-22 años de descanso en suelos arcillo-limosos alcalinos algo salinos, lo cual se refleja en un incremento de pH y de la conductibilidad eléctrica. Suelos con zonación química similar son descritos por Navarro (1993) sobre relieves del abanico fluvial de piedemonte o en llanuras fluvio-lacustres semiendorreicas del Altiplano. Probablemente el aumento de sales alcalinas en suelos puede explicarse por la cercanía del río Qhora que cruza Patarani, saliendo de su cauce y ocasionalmente aportando sedimentos. En el invierno se seca completamente, mostrando en su lecho afloramientos de sales. Beck (1985) y Navarro (1993) describen a las especies que crecen en este tipo de suelos alcalinos Muhlenbergia fastigiata, Parastrephia lepidophylla y/o Festuca orthophylla; con excepción a la última, estas especies también están representadas en nuestro análisis multivariado (Fig 10a), donde encontramos además a Distichlis humilis. Esta gramínea indicadora de las praderas salobres con suelos alcalinos, cálcico-salinos, del tipo solonchak ocupa grandes extensiones en playas fluvio-lacustres altiplánicas estacionalmente anegadas de forma somera (Navarro 2002), aspecto encontrado también por Ruthsatz (1977), Seibert (1983) y Garcia (1997).
Zeballos et al. (2003) describen la amplia distribución de Parastrephia lepidophylla en su contribución a la flora de Oruro, donde este arbusto siempreverde se desarrolla en planicies y laderas secas, típicas de suelos con capa freática poco profunda. Esto se corrobora en el estudio de Loza (1998), que además explica que los suelos con capa freática poco profunda presentan por lo general textura arcillosa, debido a que se compactan fuertemente, reteniendo la percolación del agua. También nuestras parcelas de 9-22 años presentan una textura arcillo-limosa, desarrollándose especies que requieren mayor humedad como Eleocharis albibracteata y Muhlenbergia fastigiata.
]]> Conclusiones
El tipo de dinámica sucesional en parcelas de descanso en la comunidad de Patarani Altiplano central correspondería al modelo de ecosistema de proclimax (Grime 2001), donde las especies que dominan se han adaptado a las constantes presiones de disturbio histórico del pastoreo y extracción de leña, además de las adversidades del clima. Esto se observa en el reducido número de especies presentes y en la heterogeneidad entre parcelas de un mismo grupo de edad, que nos estaría indicando la ocurrencia de rutas divergentes en la sucesión, en que las especies que colonizan y dominan son comunidades que están condicionadas por las características edáficas (Navarro 1993) y ambientales de cada sitio. Algunos otros factores, que determinan rutas divergentes, son la distancia a las fuentes de semillas (parcelas vecinas) y la presencia y características de un banco de semillas permanente. Mediante el análisis de los resultados - corroborado por datos bibliográficos - podemos identificar algunos patrones del cambio de la vegetación durante la sucesión y las especies que aparecen en las diferentes etapas de la sucesión y en diferentes condiciones edáficas.
En barbechos recientes se nota una mayor presencia de especies anuales y/o oportunistas como: Erodium cicutarium, Bromus catharticus, Tagetes multiflora y Tarasa tenella, que se mantienen durante varios años de la sucesión, lo que representa la poca diferencia de riqueza y diversidad durante los años de descanso. El aumento de especies de hábito perenne con crecimiento lento y gradual como es la gramínea Stipa ichu y las diversas especies de Nassella y los arbustos Baccharis incarum y Parastrephia lepidophylla se observan en descansos hasta de seis años, después suben con una tendencia no lineal hasta los 10 y 20 años, dependiendo de la característica edáfica, humedad, ubicación e historia de la parcela. Hay que esperar a esta edad para contar con abundantes arbustos de suficiente desarrollo vertical para ser usados como recurso leña. La reducción de los años de descanso no sería favorable para el mantenimiento de la diversidad y la riqueza florística. Aparte de los arbustos y gramíneas nombrados arriba, existen especies que solo entran después de varios años de descanso como Spergularia andina, Plantago sericea, Crassula connata y Portulaca perennis.
Con base al análisis multivariado, podemos concluir que el tipo de vegetación en este tipo de ecosistemas está relacionado con las características físico-químicas del suelo. Además, se observa que las condiciones del suelo permiten alcanzar mayor acumulación de nutrientes con los largos años de descanso (Fig. 9), corroborando las conclusiones de Hervé & Sivila (1997), Pestalozzi (1998) y Queiroz et al. (2001), aunque nuestros datos de suelos no están estadísticamente sustentados.
Agradecimientos
Este trabajo forma parte del proyecto Fertility Management in the Tropical Andean Mountain: Agroecological Bases for a Sustainable Fallow Agriculture (TROPANDES), financiado por la Comunidad Europea (INCO-DC) y realizado en Bolivia por el Instituto de Ecología en cooperación con el Institut de Recherche pour le Développement (IRD) y otras instituciones nacionales y extranjeras. Agradecemos a la Comunidad Europea por brindarnos el financiamiento, a nuestra coordinadora Prof. Tarsy Carballas de la Universidad Santiago de Compostela (España), a la red andina MOSANDES por brindar la beca a Teresa Ortuño para realizar el análisis de datos con Lina Sarmiento en la Universidad de los Andes (ULA) de Mérida (Venezuela). A la comunidad de Patarani, les agradecemos por permitirnos realizar el trabajo en sus parcelas y en especial al Sr. Ignacio Patzi y familia por acogernos en su hogar y brindarnos toda su colaboración. A Mónica Zeballos, Nelson Loza, Heidi Muñoz y Freddy Lipa, quienes sin su ayuda no hubiese sido posible realizar el trabajo de campo. A Odile Fossati por su valiosa colaboración en el análisis de multivariados. A Peter Feinsinger, Ramiro López, Dominique Hervé, Geovana Gallardo, Nelson Loza y Claudia Chumacero por sus comentarios constructivos sobre los análisis en versiones anteriores de este trabajo.
Referencias
Arno, S., E. & J. R. Habeck. 1972. Ecology of Alpine larch (Larix lyalli Parl.) in the Pacific Northwest. Ecol. Monogr. 2: 417-450.
]]> Baied, C. & J. Wheeler. 1993. Evolution of high Andean puna ecosystems, environmental, climate, and cultura change over the last 12.000 years in the central Andes. Mountain Res. Developm. 3 (2): 145-156.Barrera, C. 1994. Uso de la tierra y caracterización de poblaciones de thola (Parastrephia lepidophylla), en el cantón San José de Llanga. BS Tesis de Ingeniería Agronómica, Universidad Mayor Real y Pontificia de San Francisco Xavier, Sucre. 88 p.
[ Links ]Beck, S. G. 1985. Flórula ecológica de Bolivia. Puna semi-árida en el Altiplano boliviano. Ecología en Bolivia 6: 1-41.
[ Links ]Begon, M., J. Harper & C. Townsend. 1995. Ecología, individuos, poblaciones y comunidades. Ediciones Omega, Barcelona. 889 p.
[ Links ]Braun, O. 1964. Forrajes del Altiplano. Ministerio de Agricultura, Servicio Agrícola Interamericano. Boletín Experimental (La Paz) (30): 1-13.
Camacho, M. 2001. La gestión del espacio y las prácticas de manejo del suelo en la región altiplánica de Bolivia: el caso de la comunidad de Patarani en la provincia Aroma. Tesis de Magister Scientiae en Ecología y Conservación, Instituto de Ecología, Universidad Mayor de San Andrés, La Paz. 82 p.
[ Links ]Chessel, D. & P. Mercier.1993. Couplage de triples statistiques et liaison d’ espèces environnement. Pp. 5–44. En: D. D. Lebreton & B. Asselain (eds.). Biométrie et Environnement. Masson, Paris.
Chumacero, C. 2003. Dinámica del descanso de tierra en un sistema de cultivo tradicional, en relación a la suceción vegetal y a la fertilidad de suelos en la comunidad Titicani- Tacaca, La Paz-Bolivia. Tesis de licenciatura en Biología, Universidad Mayor de San Andrés, La Paz. 110 p.
[ Links ]Clark, D. B. 2002. Los factores edáficos y la distribución de las plantas. Pp.193-221. En: M. Guriguata & Kattan (eds.). Ecología y Conservación de Bosques Neotropicales. Libro Universitario Regional, Cartago.
[ Links ]Cohen, D. 1966. Optimizing reproduction in randomly varying environment. J. Theor. Biol. 12: 119-129.
[ Links ]Cohen, D. 1967. Optimizing reproduction in a randomly varying environment when a correlation may exist between the conditions at the time a choice has to be made and the subsequent outcome. J. Theor. Biol. 16: 1-14.
[ Links ]Connell, J. 1978. Diversity in tropical rain forests and coral reefs. Science 199: 1302-1310.
[ Links ]Davidson, D.W. 1993. The effects of herbivory and granivory on terrestrial plant succession. Oikos 68: 23-35.
[ Links ]Ellenberg, H. 1956. Aufgaben und Methoden der Vegetationskunde. Ulmer, Stuttgart. 136 p.
[ Links ]Ellenberg, H. 1981. Desarrollar sin destruir. Respuestas de un ecólogo a 15 preguntas de agrónomos y planificadores bolivianos. Instituto de Ecología, Universidad Mayor de San Andrés, La Paz. 55 p.
[ Links ]Fontaine, M. 2000. Caractéristiques écophysiologiques de quelques espèces d’une successión végétale dans les Andes vénézuéliennes. DEA d’Ecologie. Laboratoire d’Ecophysiologie végétale. Université de Paris XI. Orsay- France. Instituto de Ciencias Ambientales y Ecológicas (ICAE). Universidad de los Andes, Mérida. 39 p.
Fossati, O., M. Simier, L. Blanc & R. Marín. 2002. Tratamientos multivariables de datos ecológicos. I: Presentación y análisis simples. Informe Institut de Recherche pour le Développement Universidad Mayor de San Andrés, La Paz. 73 p.
[ Links ]García, E. 1997. Composición florística y ecología de las comunidades ruderales de las calles de la ciudad de la Paz. Ecología en Bolivia 29: 1-18.
[ Links ]Genin, D. & J. Fernández. 1994. Uso pastoril de las tierras en descanso en una comunidad agropastoril del Altiplano boliviano. Pp.199-201. En: D. Hervé, D. Genin & G. Rivière (eds.). Dinámicas del Descanso de la Tierra en los Andes. Instituto Boliviano de Tecnologia Agropecuaria (IBTA) - Institut Français de Recherche Scientifique pour le Développement en Coopération (ORSTOM), La Paz.
[ Links ]Grime, J., P. 2001. Plant strategies vegetation processes, and ecosystem properties. Ed. John Willey & Sons, Chichester. 416 p.
[ Links ]Greig-Smith, P. 1983. Quantitative plant ecology. University of California Press, Berkeley. 359 p.
[ Links ]Haase, R. 1990. Community composition and soil properties in north Bolivian savanna vegetation. Journal of Vegetation Science 1: 345-362.
[ Links ]Hammer, R. D. 1988. Space and time in the soil landscape: ill defined universe. Pp.105-140. En: D. L. Pederson & V. T. Paker (eds.). Ecological Scale. Theory and Applications. Columbia University Press, Nueva York.
[ Links ]Hervé, D. 1994 a. Desarrollo sostenible en los Andes Altos: los sistemas de cultivo con descanso largo pastoreado. Pp. 15-36. En: D. Hervé, D. Genin & G. Rivière (eds.). Dinámicas del Descanso de la Tierra en los Andes. IBTA-ORSTOM, La Paz.
[ Links ]Hervé, D. 1994 b. Respuesta de los componentes de la fertilidad del suelo a la duración del descanso Pp. 155 - 170. En: D. Herve, D. Genin & G. Rivière (eds). Dinámicas del Descanso de la Tierra en los Andes. IBTA-ORSTOM, La Paz.
[ Links ]Hervé, D. & R. Sivila. 1997. Efecto de la duración del descanso sobre la capacidad de producir en las tierras altas de Bolivia. Pp. 189-199. En: Liberman, M. & C. Baied (eds.) Desarrollo Sostenible de Ecosistemas de Montaña: Manejo de Áreas Frágiles en los Andes. The United Nations University, Liga de Defensa del Medio Ambiente, Instituto de Ecología, Secretaria ejecutiva PL 480. Artes Gráficas Latina, La Paz. 383 p.
[ Links ]Huston, M.A. 1994. Biological diversity. Cambridge University Press, Cambridge. 686 p.
[ Links ]Inouye, D., W. 1987. Old field succesion on Minnesota sand plain. Ecology 68: 12-26.
[ Links ]Jongman, R., C. Ter Braak & O. Van Tongeren. 1996. Data analysis in community and landscape ecology. Cambridge University Press, Cambridge. 299 p.
[ Links ]Kessler, M. & P. Driesch. 1993. Causas e historia de la destrucción de bosques altoandinos en Bolivia. Ecología en Bolivia 21: 1-18.
]]>Kent, M. & P. Coker. 2000. Vegetation description and analysis, a practical approach. John Wiley & Sons, Nueva York. 363 p.
[ Links ]Lara, R. & C. Zapata. 2001. Mapa de biomasa vegetal potencial de Bolivia (preliminar). Proyecto de coordinación e implementación del plan de acción Forestal para Bolivia. FAO-PAFBOL, La Paz. 16 p.
[ Links ]López, R. 1999. Cantidad mínima de lluvia que induce la germinación en un semi-desierto andino, la Prepuna (Potosí, Bolivia). Ecología en Bolivia 33: 63-73.
[ Links ]Lorini, J. 1994. La agroecología y el desarrollo altiplánico. El Modelo La Paz - Bolivia. Instituto de Ecología, Universidad Mayor de San Andrés, La Paz. 82 p.
[ Links ]Loza, F. 1998. Flora y vegetación arvense en dos comunidades campesinas del Altiplano norte (Prov. Omasuyos, La Paz) Tesis de licenciatura en Biología, Universidad Mayor de San Andrés, La Paz. 100 p.
[ Links ]Magurran, A. 1987. Ecological diversity and its measurement. Croom Helm, Londres. 179 p.
[ Links ]Margalef, R. 1968. Perspectives in ecological theory. Chicago Press, Chicago. 110 p.
[ Links ]Markgraf, V. 1985. Paleoambiental history of last 10.000 years in norwest Argentina. Zentralbl. Geol. Paläontol., Teil 2, Hist. Geol Paläontol. 11 (12): 1739-1749.
[ Links ]Matteucci, S & A. Colma. 1982. Metodología para el estudio de la vegetación. Secretaria General de la Organización de Estados Americanos, Programa Regional de Desarrollo Científico y Tecnológico. Washington, DC. 166 p.
[ Links ]Navarro, G. 1993. Vegetación de Bolivia: Altiplano meridional. Rivasgodaya 7: 69-98.
[ Links ]Navarro, G. 2002. Vegetación y unidades biogeográficas. Pp. 1-500. En: G. Navarro & M. Maldonado (eds).Geografía Ecológica de Bolivia. Vegetación y Ambientes Acuáticos. Centro Ecología Simón I. Patiño – Departamento de Difusión, Cochabamba.
Odum, E. P. 1963. Ecology. Holt, Rinehart and Winston, Nueva York. 201 p.
[ Links ]Pérez, R. & M. Arriaga. 2000. Interpretación ecológica de la ganadería extensiva y sus interrelaciones con la agricultura en el piso agrícola del páramo de Gaviria, Andes venezolanos. Trabajo para Magíster Scientiae en Ecología Tropical, Universidad de los Andes, Mérida. 161 p.
[ Links ]Pestalozzi, H. 2000. Sectoral fallow systems and the management of soil fertility: the rationality of indigenous knowledge in the high Andes of Bolivia. Mountain Research and Development 20 (1): 64-71.
[ Links ]Pickett, S., T., A. & P.S. White (eds.). 1985. The ecology of natural disturbance and patch dynamic. Academic Press, Nueva York. 470 p.
[ Links ]Posnansky, M. 1983. Los efectos sobre la ecología del Altiplano de la introducción de animales y cultivos por los españoles. Pp. 13-22. En: E. Geyger & C. Arce (eds.) Ecología y Recursos Naturales en Bolivia. Centro Portales, Cochabamba.
[ Links ]Queiroz, J., C. Barrera & J. Valdivia. 1994. La estructura y composición botánica de la vegetación de los campos agrícolas de la comunidad de San José Llanga, Altiplano central, en función de los suelos y periodo de descanso. Pp. 215-224. En: D. Hervé, D. Genin & G. Rivière (eds). Dinámicas del Descanso de la Tierra en los Andes. IBTA-ORSTOM, La Paz.
[ Links ]Queiroz, J. C. 2001. Ecology and natural resources of San José de Llanga. Pp. 59-112. En: Coppock, L. & C. Valdivia. 2001. Agropastoralismo Sostenible en el Altiplano Boliviano: El caso de San José Llanga. USAID, La Paz.
[ Links ]Ramos, A. 1995. Relaciones de presión de pastoreo, productividad animal y disponibilidad de forraje en áreas de pastoreo del Cantón San José de Llanga, Provincia Aroma, Departamento de La Paz. Tesis de licenciatura en Ingeniería Agronómica, Universidad Mayor Autónoma Tomás Frías, Potosí. 67 p.
[ Links ]Ruthsatz, B. 1977. Pflanzengesellschaften und ihre Lebensbedingungen in den Andinen Halbwüsten Nordwest – Argentiniens. Dissertationes Botanicae 39: 1-168.
]]>Ruthsatz, B. 1983. Der Einfluss des Menschen auf die Vegetation semiarider tropischer Gebirge am Beispiel der Hochanden. Ber. Dt. Bot. Ges. 96: 535-576.
[ Links ]Sarmiento, L., L. D. Llambí, A. Escalona & J. Marquez. 2003. Vegetation patterns, regeneration rates and divergence in an old-field succession of the high tropical Andes. Plant Ecology 166 (1): 145-156.
Sarps. A. 2001. Caractérisation fonctionnelle des principales espèces d’une succession secondaire. Diplôme d’Ingénieur de techniques agricoles, ENITA de Clermont-Ferrand, Université Paris XI. 43 p.
Seibert, P. 1983. Human impact on landscape and vegetation in the central High Andes. Pp. 261-276. En: W. Holzner, M. J. A. Werger & I. Kusima (eds.). Man’s Impact on Vegetation. Dr. W. Junk Publishers, La Haya.
Shmida, A. & M. V. Wilson. 1985. Biological determinant of species diversity. J. Biogeogr. 12: 1-20.
Siccama, T.G., F.H. Bormann & G.E. Likens. 1970. The Hubbard Brook ecosystem study: productivity, nutrients and phytosociology of the herbaceus layer. Ecol. Monogr. 40: 389-402.
Svoboda, J. & G. Henry. 1987. Succession in marginal artic environments. Artic and Alpine Research 19: 373-384.
Venable, D. L.& L. Lawler. 1980. Delayed germination and dispersal annual in desert: escape in space and time. Oecologia 46: 272-282.
Villavicencio, X. 1990. Floristischvegetationskundliche Untersuchungen im nördlichen bolivianischen Altiplano, Gemeinde Titicani-Tacaca. Tesis de diploma Universidad Berlin. 84 p.
[ Links ]Zar J. H. 1999. Bioestadistical analysis. 4ta Ed. Prentice may, Upper Saddle River, Nueva Jersey. 663 p.
[ Links ]Zeballos M., E. García & S. G. Beck. 2003. Contribución al conocimiento de la flora departamental de Oruro. Liga de Defensa del Medio Ambiente, Herbario Nacional de Bolivia, Museo Nacional de Historia Natural. Artes Gráficas Latinas, La Paz. 84 p.
[ Links ] ]]>