Vegetación, suelos y clima en los diferentes pisos altitudinales de un bosque montano de Yungas, Bolivia: Primeros resultados
Vegetation, soil and climate in different altitudinal belts in montane cloud forests of the Yungas, Bolivia: first results
Vegetation, Boden und Klima in unterschiedlichen Höhenstufen eines Bergregenwaldes in den Yungas Boliviens - erste Ergebnisse
Kerstin Bach1, Markus Schawe2, Stephan Beck3, G. Gerold2, S.R. Gradstein1 & Mónica Moraes R.4
]]>1Albrecht-von-Haller-Institut für Pflanzenwissenschaften, Abteilung Systematische Botanik, Untere Karspuele 2, 37073 Goettingen, Alemania, bach.kerstin@gmx.de Fax: ++49-551-392329.
2Georg-August-Universitat Göttingen, Abteilung Landschaftsökologie, Goldschmidtstr. 5, 37077 Göttingen, Alemania, mschawe@gwdg.de Fax ++49-551-3912139
3Herbario Nacional de Bolivia, Casilla 10077, La Paz, Bolivia
4Instituto de Ecología, Universidad Mayor de San Andrés, Casilla 10077, La Paz, Bolivia
Resumen
Desde junio de 2000 se ha iniciado un trabajo interdisciplinario en el Parque Nacional Cotapata (Yungas, Bolivia) a lo largo de un transecto altitudinal de 1.700 a 3.400 m. La meta común del proyecto es la investigación de las interacciones ecológicas entre factores bióticos y abióticos en base a la zonificación de la vegetación. Se reconocen tres tipos de vegetación: el subalpino tropical que se extiende hasta aproximadamente 3.150 m, luego el piso altimontano hasta los 2.100 m y en la base se encuentra el piso montano. Los transectos verticales corresponden a los pisos climáticos andinos de tierra templada y tierra fría con un gradiente de temperatura en intervalos de 12-17°C hasta 17-20° C.
El análisis de los suelos demuestra la siguiente diferenciación hipsométrica: En el piso montano dominan los suelos Humic Dystrudepts (según Soil Survey Staff 1998) A partir del piso altimontano y en relación con la inclinación, el microrelieve y la edad de los suelos se presentan en los siguientes tipos de suelo: Typic Placaquod, Spodic Dystrudept, Typic Durorthod y Histic Humaquept. Para la formación de suelos, la disminución de la evapotranspiración con el aumento de la altitud representa un factor decisivo. Cuando la evapotranspiración es baja, hay consecuencias directas para la dinámica del agua del suelo, ya que la infiltración aumenta. A 3.000 m de altitud, el contenido del agua del suelo está permanentemente en el rango de capacidad de campo. Estos resultados indican que la dinámica del agua del suelo podría influir significativamente en la distribución de las plantas terrestres y representan un factor importante para explicar la clasificación florística.
]]> Palabras clave: Bolivia, bosques montanos húmedos, clima, suelos, tipos de vegetación.Abstract
Since June 2000 an interdisciplinary research project is carried out in the Cotapata National Park (Yungas, Bolivia), which investigates climate, soils and vegetation covering an elevational gradient of 1.700 to 3.400 metres. The main objective is to determine correlations and interactions between biotic and abiotic factors along the elevational gradient. Four elevational vegetation types have been recognized: the montane forest (up to ca. 2.100 m), the high montane forest I and II (ca. 2.100-3.150 m) and the tropical subalpine forest (up to 3.400 m). Annual air temperature decrease from 17°-20°C to 12°-17°C with increasing elevation and 0.63°C per each 100 m elevation range.
The soil examinations show the following hypsometrical soil differentiation: in the montane forest Humic Dystrudepts (according to Soil Survey Staff 1998) are dominant. From the high montane forest the following soil types appear in relation to slope inclination, micro relief and soil age: Typic Placaquod, Spodic Dystrudept, Typic Durorthod and Histic Humaquept. Reduction of evapotranspiration with increasing elevation is a decisive factor for soil development. At 3.000 m the ground water almost permanently attains field capacity. It is proposed, that ground water balance is a keyfactor affecting the distribution of terrestrial plant species and the vegetation classification along the gradient.
Keywords: Bolivia, tropical montane cloud forests, climate, soil, vegetation types.
Zusammenfassung
Seit Juni 2000 finden im Cotapata - Nationalpark (Yungas, Bolivien) interdisziplinäre Untersuchungen zu Klima, Boden und Vegetation in einem Höhengradienten von 1700 m ü.M. bis 3400 m ü.M. statt. Gemeinsames Projektziel ist die Untersuchung der für die Vegetationszonierung entscheidenden ökologischen Wechselwirkungen zwischen biotischen und abiotischen Faktoren. Erste Ergebnisse dieser Forschung liegen vor. Vier Vegetationstypen lassen sich erkennen. An eine montane Bergstufe schliesst sich ab ca. 2100 m ein hochmontaner Wald an, der in 3150 m in die tropisch subalpine Stufe übergeht. Der hochmontane Wald lässt sich in eine untere und obere Stufe unterteilen. Bei ganzjährig humiden Klima umfassen die Vertikaltransekte einen Temperaturgradienten von 17-20°C bzw. 12-17°C Jahresmitteltemperatur, mit einer Differenz von 0.63°C per 100 Höhenmeter.
Die Bodenuntersuchungen zeigen folgende hypsometrische Bodendifferenzierung: In der montanen Stufe sind Humic Dystrudepts (gemaess Soil Survey Staff 1998) dominant. Ab der hochmontanen Stufe treten in Relation zu Hangneigung, Mikrorelief und Bodenalter folgende Bodentypen auf: Typic Placaquod, Spodic Dystrudept, Typic Durorthod und Histic Humaquept. Für die Bodenentwicklung entscheidend ist der Rückgang der Verdunstung mit zunehmender Höhe. Damit ist ganzjährig ein hoher Sickerwasserstrom mit zunehmender Höhe gegeben. Es ist zu vermuten, dass der Bodenwasserhaushalt sich vor allem auf die Verbreitung terrestrischer Pflanzenarten auswirkt und ein Faktor zur Erklärung der Vegetationsklassifikation ist.
Schlüsselworte: Bolivien, Bergregenwald, Klima, Boden, Vegetationstypen.
]]>
Introducción
A escala mundial, los bosques montanos húmedos son ecosistemas frágiles, los cuales tienen una importancia enorme en relación a los recursos de agua, energía y biodiversidad (Agenda 21, cap. 13). La FAO (Food and Agriculture Organisation) contribuye a la importante tarea de fomentar su protección y uso duradero mediante la campaña “International Year of Mountains 2002". En el año 2000, se inició un proyecto de investigación en el Parque Nacional de Cotapata destinado a estudios hipsométricos de factores bióticos y abióticos en el bosque montano húmedo. El proyecto está coordinado por la cooperación boliviano - alemana entre la Universidad de Göttingen (Departamento de Ecología Paisajística y de Botánica Sistemática) y el Instituto de Ecología de Universidad Mayor de San Andrés, en La Paz.
Se investiga por parte la ecología paisajística, los cambios causados por la altitud (cambios hipsométricos) del clima y del suelo en unidades de vegetación. Por el Instituto de Botánica Sistemática de la Universidad de Göttingen se determinan los tipos de vegetación característicos del área y se realizan los análisis ecoestadísticos.
Area de estudio
En la parte oeste de la Cordillera Oriental de Bolivia, se ha desarrollado un gradiente vertical característico para los trópicos interiores húmedos, en los cuales se ha realizado reducida investigación hasta ahora. Esa región se caracteriza por una gran biodiversidad florística, como también por un cambio a escala local pequeña de los parámetros ecológicos dependientes del relieve. Aquí se encuentra el Parque Nacional de Cotapata, localizado aproximadamente a 80 km al noreste de la ciudad de La Paz. Este Parque Nacional fue establecido en el año 1993 con una área de 580 km2. El gradiente vertical va desde los 1.100 hasta 5.600 m con pendientes escarpadas. El difícil acceso a esta región ha favorecido el hecho de que muchas regiones del Parque permanezcan intactas; solamente en los márgenes del área protegida, el manejo integrado está admitido. Al interior del Parque fue construida la Estación Biológica de Tunquini – dependiente del Instituto de Ecología y que funciona bajo un acuerdo con el Sistema Nacional de Areas Protegidas de Bolivia–, en que se cuenta con una adecuada infraestructura para realizar proyectos de investigación.
Metodología
Vegetación
]]> Los estudios florísticos fueron realizados en base a grupos indicadores. Este método permite el estudio de un alto número de relevamientos fitosociológicos a lo largo del transecto y en corto tiempo (Gentry 1982, Frahm & Gradstein 1991, Kessler & Bach 1999). Uno de los grandes impedimentos para estudios de la vegetación en bosques tropicales es el elevado número de especies. Otro problema frecuente radica en la dificultad de identificación de las especies encontradas. Además, para el estudio de árboles se requiere de áreas mucho mayores para encontrar un número representativo de especies. Por consecuencia, es difícil encontrar áreas ecológica y florísticamente homogéneas en lugares montañosos con un microrelieve muy diferenciado.Por estos motivos, se ha elegido según el método de Kessler & Bach (1999) los siguientes grupos de plantas: Pteridophyta, Araceae, Bromeliaceae, Melastomataceae, Palmae y Cactaceae. Estos grupos contienen menor número de plantas árboreas y mayormente consisten en plantas epífitas y hierbas. Su distribución geográfica o ecológica está bien delimitada y como grupo, tienen una distribución amplia en una variedad de hábitats.
La clasificación de la vegetación fue basada en el método Mueller-Dombois & Ellenberg (1974).
Clima
Las condiciones básicas de una investigación hipsométrica están dadas en un transecto del bosque intacto, manteniendo la misma exposición y el mismo sustrato geológico. Por este motivo, se eligió un transecto en el lado sureste del cerro Hornuni de 1.700 a 3.400 m. Este transecto vertical corresponde a los pisos climáticos andinos de tierra templada y tierra fría. Tanto la variación del clima y de la vegetación, como la diferencia hipsométrica de los suelos son condiciones climáticohidrológicas de los bosques montanos que hasta ahora han sido poco estudiadas (Bruijnzeel 2000). Desde el mes de julio 2000 hasta el mes de noviembre 2000, fueron realizadas investigaciones paralelas sobre la vegetación, los suelos y el clima a lo largo del transecto. De esa manera, los resultados obtenidos permiten una comparación directa de los factores abióticos y bióticos.
En la parte central de las unidades de vegetación, se instalaron tres estaciones complejas de clima, con exposición sureste. La estación 1 está ubicada a 1.840 m, la estación 2 a 2.550 m y la estación 3 se encuentra a 3.010 m. Con el fin de parametrizar la influencia del macroclima sobre la vegetación, se retiró la vegetación del área alrededor de las estaciones climáticas. Está prevista la medición continua de los siguientes parámetros climáticos: temperatura del aire, humedad del aip, precipitación vertical, aporte de agua de la niebla, radiación global, balance de radiación, radiación fotosintética activa (PAR), radiación UV-B, velocidad y dirección del viento, temperatura del suelo (4 profundidades), flujo de calor desde el suelo y contenido de agua del suelo (5 profundidades). Estos parámetros se registrarán cada 10 minutos con los dataloggers y será calculado el promedio de una media hora.
El cambio hipsométrico del clima dentro de la vegetación está siendo registrado en base a los parámetros principales de temperatura y humedad del aire. Para esto se han instalado a lo largo del transecto principal, entre los 1.800 y 3.200 m, un total de 23 dataloggers compactos. Cada 200 m de altitud, se instalaron en los árboles tres dataloggers compactos a 50 cm, 2 m y 15 m, sobre el nivel del suelo. Este método experimental garantiza una buena posibilidad de comparación entre las unidades de vegetación.
La diferenciación del microclima dentro de la vegetación para las diferentes unidades de relieve y las exposiciones diferentes se están investigando con los dataloggers compactos, en un transecto horizontal al nivel de la estación climática l (1.840 m),dispuestos también a 50 cm, 2 m y 15 m por encima del nivel del suelo en los árboles.
El análisis de la evapotranspiración potencial (pET) se realizó en base a los datos horarios según la fórmula modificada de Penman de Doorenbos & Pruitt (1988). Como ejemplo de la pET.
Suelos
]]> Los análisis pedomorfológicos tienen lugar en el mencionado transecto con puntos de muestreo cada 50-100 metros de altitud. Los suelos fueron analizados en el campo y en el laboratorio. El análisis contiene los parámetros pH, CICef, Ct, Nt, óxidos pedogénicos, el contenido total de los elementos, mirconutrientes, textura y densidad aparente.
Resultados
Vegetación
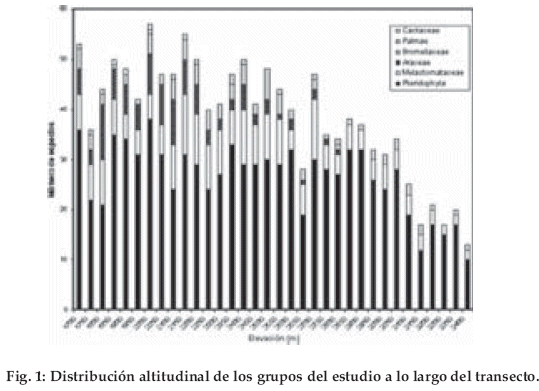
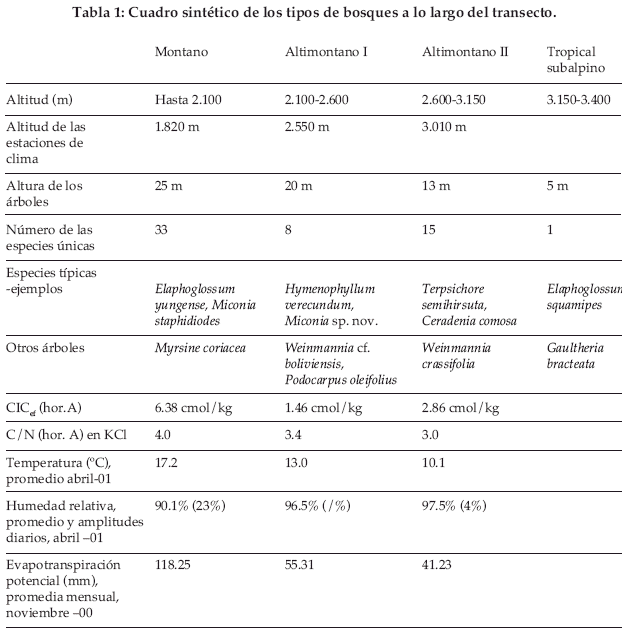
En el año 2000 se realizaron 65 relevamientos fitosociológicos, en base a los grupos indicadores con más de 2.400 registros. Encontramos estos grupos de plantas frecuentemente a lo largo del transecto y solamente las cactáceas fueron una excepción (Fig. 1). El porcentaje de las colecciones identificadas hasta julio 2001 alcanza el 75%, lo cual podría ascender hasta 88%, si descontamos a las Melastomataceae (Fig. 2). Considerando el número total de especies, el grupo más importante corresponde a las pteridófitas (70.5%), seguido por las melastomatáceas (14.5%), aráceas (7.6%) y las bromeliáceas (5.5%). La clasificación preliminar de la vegetación en el área de estudio muestra cuatro tipos diferentes de bosques: Bosque montano (hasta 2.100 m), bosque altimontano I (2.100-2.600 m, bosque altimontano II (2.600 –3.150 m) y el bosque tropical subalpino (3.150-3.400 m), los cuales son caracterizados a continuación.
Bosque montano (por debajo los 2.100 m)
]]> En la parte inferior del transecto se encuentra el bosque montano de 25 m de altura promedio con Myrsine coriacea. El dosel normalmente no está cerrado y en el sotobosque dominan dos especies hemiepífitas: Blechnum ensiforme y Philodendron ornatum. Las rubiáceas frecuentes en el sotobosque son Condaminea corymbosa, Faramea cf. multiflora, Ladenhergia cf . multiflora y Palicourea aff. semirasa.Bosque altimontano I (2.100-2600 m)
Por encima de los 2.100 m encontramos al bosque altimontano, que se caracteriza por la presencia de Podocarpus oleifolius y Weinmannia boliviana. Esta faja altitudinal se divide en dos tipos diferentes los cuales necesitan ser más investigados. Hasta los 2.600 m, Racinea schumaniana y Miconia sp. nov. son dominantes. El bosque crece hasta 20 m de altura en promedio y todavía el dosel está cerrado.
Bosque altimontano II (2.600-3.150 m)
En dirección al piso tropical subalpino, la altura de los árboles disminuye y alcanza en el tipo altimontano II una altura de 13 m promedio. Los musgos ganan más en importancia y entre los grupos indicadores son mayormente las pteridófitas epífitas que caracterizan este tipo de vegetación, como Terpsichore semihirsuta y Ceradenia comosa. El bosque está más iluminado pues a menudo el dosel se observa abierto. Las rubiáceas frecuentes en este tipo de bosque son Palicourea bachtienii, Coccocypselum lanceolatum, Nertera granadensis y Psychotria tristis.
Bosque tropical subalpino (3.150-3.400 m)
Por encima de los 3.150 m, empieza el piso tropical subalpino. Se encuentran especies claramente escleromorfas. Los árboles crecen aislados y alcanzan una altura máxima de 5 m apenas. Se encuentran muchos árbustos como Gaultheria bracteata o plantas epífitas como Elaphoglossum squamipes. Por encima de los 3.400 m encontramos al “páramo yungeño“.
A lo largo del transecto encontramos frecuentemente a Hymenophyllum plumieri. Además con una distribución amplia se tiene a Hymenophyllum fucoides (s.l.), H. ruizianum y Terpsichore laxa. Sin embargo, esas plantas no suben al bosque tropical subalpino.
Clima
El mesoclima del área de estudio se caracteriza por una humedad permanente a largo del año. La precipitación anual registrada en 1.800 m y 2.550 m es de 2.500 mm y aumenta a altitudes entre 3.000 - 3.000 mm.
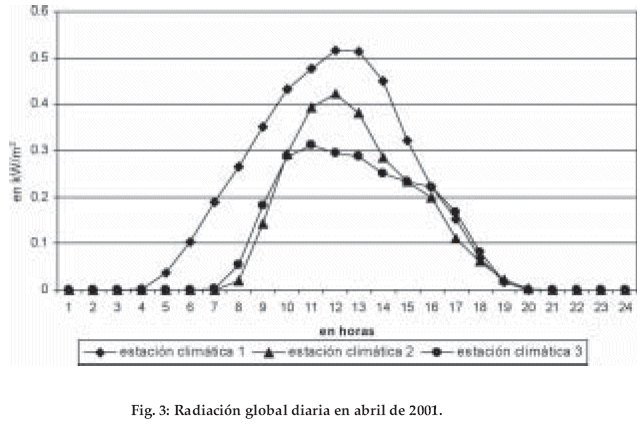
]]> Los datos del Valle del Zongo (región colindante con el área de estudio) son comparables con la zona estudiada, la precipitación anual registrada a partir del primer nivel de condensación (entre los 800 y 1.400 m) es mayor a 2.000 mm (Energoprojekt-Hidroinzenjering S.A. 1998).La radiación global, un factor importante para la determinación de la evapotranspiración, disminuye con la altitud. El mes de abril de 2001 es un ejemplo representativo de esta relación: a la altitud de 1.820 m, la mayor radiación global registrada presenta un máximo 0.5 kW/m2, a los 2.550 m un 0.4 kW/m2 y a los 3.010 m un máximo de 0.3 kW/m2 (Fig. 3). La disminución de la radiación ocurre debido al aumento de la nubosidad con el incremento de la altitud. Con la temperatura del aire se observa también un gradiente evidente. El promedio de la temperatura mensual en la estación 1 a los 1.820 m es de 17.2°C, en la siguiente estación 2 a los 2.550 m con 13.0°C y en la estación 3 a 3.010 m con 10.1°C. La amplitud diaria corresponde al orden de la temperatura media: Bosque altimontano I 6.1°C, bosque altimontano II 4.0°C, bosque tropical subalpino 4.0°C (Fig. 4, tabla 1). El gradiente de las temperaturas promedio (registradas por las estaciones climáticas) es de 0.63°C por cada 100 m de diferencia de altitud. Comparativamente, los mismos registros hechos en la vegetación a la altura de 2 m sobre el nivel del suelo, muestran una disminución de la temperatura en 0.51°C, debido a la mayor humedad presente en la vegetación.
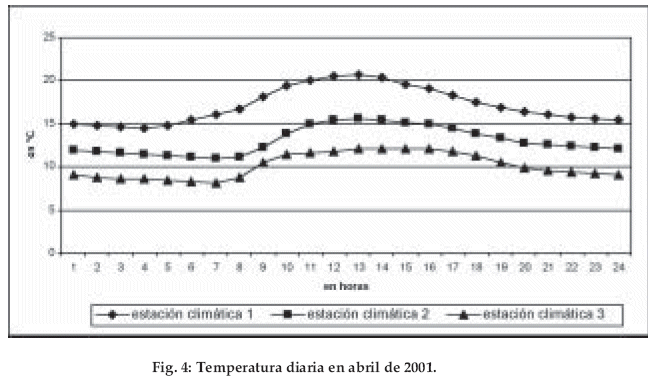
Con respecto a la humedad relativa, se presentan las siguientes correlaciones: Los valores máximos se encuentran en la estación 3 con un 97.5% de humedad relativa correspondiente a las amplitudes menores al 4%, mientras que los valores mínimos se registran en la estación 1 con un 90.1% y una amplitud del 23%. Cuando la humedad relativa es mayor, las amplitudes son menores. La humedad relativa máxima corresponde a las oscilaciones mínimas de la temperatura.
Se muestran las primeras relaciones de la evapotranspiración desde abril de 2001. La pET máxima se observa en la estación 1 a 1.820 m, mientras que la intensidad de pET a 2.550 m y a los 3.010 m es mucho menor. Los cálculos mensuales confirman estas grandes diferencias: a los 1.810 m se registran 118.25 mm, a 2.550 m es de 55.31 mm y a los 3.010 m con 41.23 mm (Fig. 5, Tabla 1).
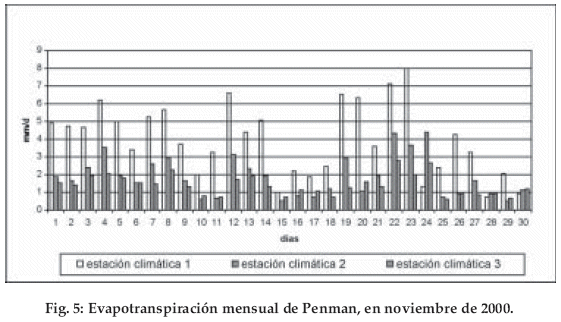
Las amplitudes de las sumas de los valores diarios se ven evidentemente influenciadas por el cambio hipsométrico y por los cambios en la nubosidad, las diferencias entre un día soleado y un día nublado. Cuando en la estación 1 hay una diferencia de 3 ó hasta 4 veces entre un día soleado y un día nublado, en la estación 3 la más alta, muestra solamente una diferencia de hasta 1.5 veces.
Suelos
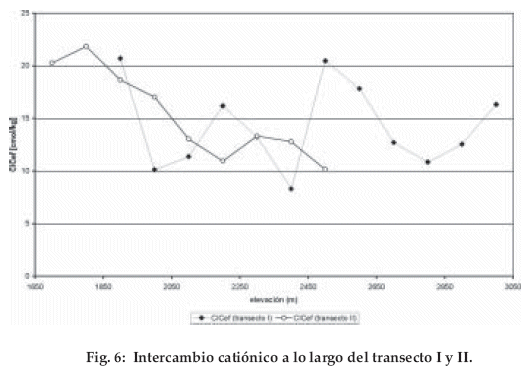
Como en otros bosques tropicales montanos en Suramérica y Centroamérica, los suelos están marcados por una acidificación muy fuerte (pH debajo de 4.5 en todos los horizontes y en el horizonte A y O, por debajo de 4), un CIC muy bajo (< 5 cmol/kg), una saturación fuerte de Al3+ (a una profundidad mayor de 50 cm superior al 70-90%), la proporción entre C y N mayor de 20, y una capa orgánica muy gruesa (30 cm) (Tab. 1).
Hacia la zona del bosque tropical subalpino, la génesis de los suelos está caracterizada por la presencia de Podzol y la acumulación de humedad (Typic Placaquod y Typic Durorthod) según Soil Survey Staff (1998). Los análisis de los parámetros pedofísicos y pedoquímicos, tanto en campo como en el laboratorio permiten apreciar la diferenciación de los suelos debido al cambio hipsométrico. En el bosque altimontano predominan Humic Dystrudepts. En el bosque altimontano II (21 00-3.100 m) varía el tipo de suelos y la profundidad de la materia orgánica. En relación a inclinación, microrelieve y edad de los suelos se presentan: Typic Placaquod, Spodic Dystrudept, Typic Durorthod y Histic Humaquept.
Discusión
Los límites de bosque encontrados en el presente trabajo son parecidos a los resultados obtenidos por Gradstein & Frahm (1987) en el norte del Perú, en la Cordillera oriental de los Andes. Estos tipos de vegetación corresponden al „bosque húmedo montañoso de Yungas“ (700m), al „bosque nublado de ceja“ (2.000–3600m) y al ecotono arbustivo-arbóreo de la „pradera parámica húmeda de alta montaña“ (Ribera 1992).
Cuando los diferentes tipos de la vegetación tienen una correlación con los tipos de suelo dominantes, no existe una disminución de la disposición de los nutrientes con la altitud (por ejemplo Tanner 1980) en los Yungas en Bolivia. E1 ecosistema de los bosques montanos está adaptado a los suelos pobres en nutrientes y a cubrir solamente sus requerimientos de materia orgánica. Las investigaciones planificadas analizarán los factores de estrés pedohidrológico y pedoquímico en relación a la diferenciación de la vegetación con el cambio hipsométrico.
]]> En la transición al bosque tropical subalpino se observa una dominancia de Typic Placaquod. El color de los suelos, el desarrollo de cintas de hierro, poco contenido de óxidos pedogénicos y traslado de sesquióxidos (Fe, Al) indican que en algunos perfiles la edad es muy avanzada con una podsolisación vertical y horizontal (Sommer et al. 2000).Hasta el bosque altimontano II, las bases intercambiables se concentran en la materia orgánica. A diferencia de las cordilleras de Venezuela (Hetsch 1976), Colombia (Hetsch & Folster 1979, van der Hammen & Ruiz 1984) y Ecuador (Schrumpf et al. 2001), los parámetros del suelo no tienen ningún gradiente vertical significativo (Fig 6). La estructura de los perfiles se diferencia por el relieve (inclinación, microrelieve con diferente drenaje de suelo), por la influencia de la composición de la vegetación, por la calidad y cantidad de materia orgánica y la edad de los suelos. Estas características también permiten diferenciar los tipos de suelo de bosques montanos de la zona alta.
Conclusiones
Para la descripción de las conexiones causales entre vegetación, clima y suelos podemos concluir solamente afirmaciones preliminares.
La clasificación preliminar de los suelos muestra una dominancia de Humic Dystrudept en la faja montana. El cambio de la vegetación en el bosque altimontano I se sincroniza con un cambio del tipo de suelo; este tipo de suelos Placaquods se ve influenciado por la retención de agua. En la faja altimontano II, el cambio de suelos no es significativo con un Typic Placaquod.
La evapotranspiración potencial según Penman muestra claramente una disminución con la altitud. Eso se encuentra también en la dinámica del agua del suelo: por un lado tenemos suelos en la faja montana suelos, los cuales carecen o solamente temporalmente tienen la influencia por retención de agua. Por otro lado, tenemos suelos en la faja altimontana II, los cuales están saturados de agua durante todo el año. Estos resultados indican que la dinámica del agua del suelo podría influir significativamente en la distribución de las plantas terrestres y representan un factor importante para explicar la clasificación florística.
Las influencias de la evapotranspiración sobre especies epífitas tiene que ser mejor investigada. Por encima de la faja altimontana I, se encuentra una capacidad de intercambio catiónico muy baja, pobre en bases y una alta saturación de aluminio. Si estos factores requieren una selección florística, deberían ser verificados en el futuro con otros muestreos y análisis de suelos.
]]>Agradecimientos
Para los datos suplementarios de flora agradecemos a E. García (Rubiaceae), C. Maldonado y N. Paniagua del Herbario Nacional de Bolivia. La determinación florística de los especímenes hubiera sido imposible sin la ayuda de T. Croat (Aráceas), M. Kessler (Cactaceae y Elaphoglossum), M. Moraes (Palmae), S. Renner (Melastomataceae), A. Smith (Pteridophyta) y T. Kromer (Bromeliaceae). Por su apoyo en el campo agradecemos a J.-K. Biener, J. Hanspach, I. Jimenez, L. Joachim, P. Just, O. Koch, H. Lange, J. Lendzion, A. Pareja, B. Steudel, P. Vidaurre, C. Wegner, M. Wunderlich y los habitantes de Chairo. El manuscrito fue mejorado por sugerencias de A. Pareja, J. Gonzales, A. Gorrin y los editores.
Referencias
Bruijnzeel,L.A. 2000. Hydrology of tropical montane cloud forests: a reassessment. En: Gladwell, J.S. (eds.): Proc. Sec. Int. Colloquium on Hydrology and Water Management in the Humid Tropics. 33 p. [ Links ]
Doorenbos, J. & W.O. Pruitt. 1988. Crop water requirements. FAO Irrigation and Drainage Paper, 24, Roma, 144 p. [ Links ]
Energoprojekt- Hidroinzenjering S.A. 1998. Estudio de factibilidad generación hidraúlica, Proyecto Coroico. Informe final. Base de Datos, Electricidad de La Paz S.A. La Paz. 51 p. [ Links ]
Frahm, J.P. & S.R. Gradstein. 1991. An altitudinal zonation of tropical rain forests using bryophytes. J. Biogeogr. 18:669-678.
Gentry, A.H. 1982. Patterns of neotropical plant species diversity. Evol. Biol. 15: 1-84.
]]> Gradstein, S.R. & J.P. Frahm. 1987. Die floristische Höhengliederung der Moose entlang des BRYOTROP - Transektes in NO-Peru. Beih. Nova Hedwigia 88: 105-113.Hetsch, W. 1976. Die Beziechung von Niederschlag und Bodenbildung in der Andenkordillere Venezuelas. Göttinger Bodenkdl. Berichte 41
Hetsch, W.& H. Foelster. 1979. Klimazonale Bodenbildung und Palaeobodenreste in den venezolanischen und kolumbianischen Anden. Z.f.Geomorphologie N.F. 33, 72-83
Kessler, M. & K. Bach. 1999. Using indicator families for vegetation classification in speciesrich Neotropical forests. Phytocoenologia 29(4).
Mueller-Dombois, D. & H. Ellenberg. 1974. Aims and methods of vegetation ecology. Wiley & Sons, Nueva York. [ Links ]
Ribera, M. 1992. Regiones Ecológicas. pp. 9-72. En: Marconi, M. (ed.): Conservación de la Diversidad Biológica en Bolivia. Centro de Datos para la Conservación. Bolivia, USAID/ Bolivia, La Paz, [ Links ]
Schrumpf, M., Guggenberger, G.,Valarezo, C.& W. Zech. 2001. Tropical montane rain forest soils. Development and nutrient status along an altitudinal gradient in the South Ecuadorian Andes. Die Erde 132: 43-59.
Soil Survey Staff. 1998. Keys to Soil Taxonomy (8th ed.) http://soils.usda.gov/ classification/keys/ RevKeysSoilTax8_02.pdf [ Links ]
Sommer, M. Halm, D. Weller, U. Zarei, M. & K. Stahr. 2000. Lateral podzolation in a granite landscape. Soll Sci.Soc.Am.J. 64: 1434-1442.
Tanner, E.V.J. 1980. Studies on the biomass and productivity in a series of montane rain forests in Jamaica. J.Ecology 68: 573-588.
]]>Van der Hammen, T. & P.M. Ruiz. 1984. La Sierra Nevada de Santa Marta (Colombia) Transecto Buritaca - La Cumbre. Studies on Tropical Andean Ecosystems 2. [ Links ]
Artículo manejado por: Renate Seidel
Recibido en: Agosto de 2002.
Aceptado en: Enero de 2003.
]]>