










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkInvestigación & Desarrollo
versión impresa ISSN 1814-6333versión On-line ISSN 2518-4431
Inv. y Des. vol.1 no.17 Cochabamba 2017
http://dx.doi.org/10.23881/idupbo.017.1-2i
ARTÍCULOS–INGENIERÍAS
MÓDULO DE ELASTICIDAD COMO CRITERIO DE REGULACIÓN HÍDRICA DEL FREJOL COMÚN (PHASEOLUS VULGARIS L.) BAJO REDUCCIONES CONTROLADAS DE RIEGO
ELASTICITY MODULUS AS A HYDRIC REGULATION CRITERION OF COMMON BEAN (PHASEOLUS VULGARIS L.) UNDER CONTROLLED IRRIGATION REDUCTIONS
Cristhian Vega Ponce y Jesús Abel Mejía Marcacuzco
Universidad Nacional Agraria La Molina, Lima-Perú
Facultad de Ingeniería Agrícola
(Recibido el 22 de febrero 2017, aceptado para publicación el 02 de mayo 2017)
RESUMEN
El propósito de esta investigación consistió en estudiar el efecto de reducciones controladas de riego sobre el módulo de elasticidad (εv) en plantas de frejol común (Phaseolusvulgaris L.) cultivadas en maceteros de respuesta hidrogravitrópica. Las plantas fueron sometidas a riego completo de raíces (RCR) y riego parcial de raíces (RPR), donde el agua asignada de acuerdo a la curva de retención agua-suelo permitió controlar y configurar cuatro tratamientos (RPR300, RPR500, RCR300 yRCR500 o control). Se monitoreó el potencial hídrico xilemático (Ψx) de las hojas, para luego construir la curva presión-volumen (P-V) y determinar εv. Los resultados mostraron que los diferentes volúmenes de agua aplicados generaron importantes variaciones en los niveles de εv; sin embargo,en los tratamientos configurados para llevar el suelo a capacidad de campo (RPR500 y RCR500) fue donde se obtuvieron los mejores desempeños de εv, efecto esperado principalmente antes de aplicar el riego a las plantas (15,63 y 15,34 MPA, respectivamente). Finalmente, aunque ambos tratamientos obtuvieron el mismo nivel de significancia de εv, RPR500 se destacó sobre el tratamiento control, porque los volúmenes de agua reducidos, combinados con el mantenimiento de diferentes valores de humedad en el suelo explorado por las raíces, pudieron ser claves en el favorecimiento de un ajuste elástico.
Palabras claves: Estrés Hídrico, Módulo de Elasticidad, Riego Parcial de Raíces.
ABSTRACT
The purpose of this research was to study the effect of controlled irrigation reductions on the elasticity modulus (εv) in common bean plants (Phaseolus vulgaris L.) cultivated in hydrogravitropic response pots. The plants were submitted to complete root irrigation (RCR) and partial root zone-drying irrigation (RPR), where the water assigned according to the water-soil retention curve allowed to control and configure four treatments (RPR300, RPR500, RCR300 and RCR500 or control). The xylem water potential (Ψx) of the leaves was monitored, and then the pressure-volume curve (P-V) was constructed to determine εv. The results showed that the different volumes of water applied generated significant variations in the levels of εv; however, in the treatments configured to bring the soil to field capacity (RPR500 and RCR500) it was where the best performance of εvwas obtained, effect expected mainly before applying the irrigation to the plants (15,63 and 15,34 MPa, respectively). Finally, although both treatments obtained the same level of significance of εv, RPR500 stood out about the control treatment, because the reduced water volumes, combined with the maintenance of different moisture values in the soil explored by the roots, could be a key in favor of an elastic fit.
Keywords: Water Stress, Elasticity Modulus, Partial Rootzone-Drying Irrigation.
1. INTRODUCCIÓN
El frejol común (Phaseolusvulgaris L.) es considerado el cultivo de mayor importancia económica y social en la agricultura familiar, debido a sus ventajosas propiedades nutricionales [1]. En el mismo sentido, es la leguminosa de mayor consumo humano; sin embargo, entre el 60 y 70 % de la producción mundial se obtiene bajo restringidas condiciones hídricas [2, 3], y aunque algunos autores han reportado que el desempeño productivo puede estar influenciado por el genotipo [1], también se señala que es una de las especies más sensibles a la falta de agua, con un potencial xilemático letal (-15 bares) similar al del tomate (Lycopersiconesculentum Mill), en comparación con otras leguminosas como el frejol caupí (Vignaunguiculata L., -25 bares) o el frejol de palo (Cajanuscajan L., -35 bares) [4].
En el caso de América Latina el 73 % de la producción proviene de regiones con niveles de déficit hídrico desde moderado hasta severo [5]; bajo este escenario el déficit de agua presente en las zonas de producción agrícola, genera fuertes necesidades que inducen a la actualización de estrategias orientadas a mejorar la aplicación del agua en las plantas [6], donde es necesario que las nuevas técnicas de riegos reducidos junto con ahorrar agua permitan mantener adecuados niveles hídricos en las plantas para lograr estabilidad de las cosechas [7, 8].
En este contexto, el riego deficitario controlado (RDC) y el riego parcial de raíces (RPR), son las metodologías que exitosamente pueden ayudar a conseguir este objetivo en una amplia variedad de cultivos de ciclo perenne [9, 10] y de ciclo corto [11, 12]. En este sentido, el principio de RPR se fundamenta en lograr mantener dos niveles de humedad diferentes en el suelo, efecto que busca conseguir en las raíces la activación de mecanismos que permitan conservar el estado hídrico de las plantas, con ahorros significativos de agua [13].
De esta forma, varias experiencias aseguran que el potencial hídrico xilemático (Yx) puede ser el mecanismo indicador de más fácil aplicación para determinar el estado y regulación hídrica en varios cultivos bajo riegos reducidos controlados[14]. Sin embargo, la elasticidad de la pared celular del tejido de las plantas(denominada módulo de elasticidad, εv),es también considerada como otro de los mecanismos fisiológicos importantes, debido a que una disminución del valor de este parámetro puede ayudar a mantener un régimen hídrico adecuado [15]; fenómeno que puede activarse como respuesta adaptativa de las plantas a las modificaciones naturales del contenido de humedad del suelo, causadas por las imperfecciones en la distribución espacial y temporal de las precipitaciones[16]; o tambiénpor modificaciones artificiales, cuando las reducciones de los niveles de agua en el suelo son controladas mediante avanzadas técnicas de riego [17].
En este sentido, algunos autores han expresado que levcorresponde a una de las interacciones que definen el modo en que funciona el sistema de regulación hídrica en las plantas, permitiendo que mantengan la hidratación celular y la turgencia, preservando el metabolismo, la expansión celular y el crecimiento [18, 19]; y aunque su determinación puede ser compleja, el resultado se compensa por la dinámica de las relaciones hídricas involucradas en su obtención [20].
En base a estos antecedentes el objetivo de este trabajo consistió en evaluar el impacto del riego parcial y el riego completo de raíces sobre el módulo de elasticidad (εv) en frejol común.
2. MATERIALES Y MÉTODOS
El trabajo se realizó entre octubre y diciembre de 2014 en el área experimental del Programa de Doctorado en Recursos Hídricos de la Universidad Nacional Agraria La Molina (UNALM) en Lima-Perú (12°0506 Latitud Sur, 76° 7500 Longitud Oeste y 230 msnm); durante el periodo de la investigación la temperatura promedio mensual fue de 18,06 ºC, humedad relativa 58,65%, evaporación 3,33 mm día-1, precipitación 0.95 mm y heliofanía 4,28 horas día-1 (Figura 1).

| Figura 1. | Ubicación del espacio donde se desarrolló el experimento en las instalaciones del PDRH-UNALM campus La Molina (Captura de Google Maps, septiembre 2016). |
Se utilizaron semillas de frejol común variedad Red Kloud, tipo determinado I; sembradas en sistemas de respuesta hidrogravitrópica, los mismos que corresponden a maceteros (Figura 2) que incorporan en su interior una mecanismo que encapsula a la raíz principal y ayuda a posicionar y orientar el crecimiento de las raíces laterales hacia las dos secciones diseñadas para aplicación del riego [7].
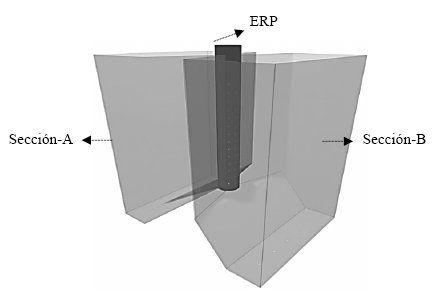
| Figura 2. | Perspectiva del macetero de respuesta hidrogravitrópica para cultivar plantas de frejol común bajo riego parcial de raíces (RPR). Se destacan sus dos secciones para el desarrollo de las raíces laterales y aplicación del riego de cada lado (A y B) y el encapsulador de la raíz principal (ERP). |
En el interior del ERP se colocó una turba comercial (Kekkilä Garden®, DMS-05) con una densidad de 0,3 g cm-3, que facilitó en primer lugar la germinación de la semilla en el segmento superior de este elemento (cama de germinación); y en segundo lugar direccionar el crecimiento de la raíz principal dentro del campo gravitacional, junto con permitir la salida de las raíces laterales por todo el eje vertical circunferencial del segmento bajo la cama de germinación, a través, de unos orificios que ayudan a orientar el crecimiento de estos órganos hacia las secciones A y B, que en su interior contenían un sustrato heterogéneo (pH: 7,67, CE: 1,35 dS m-1, materia orgánica: 5,93 %) colocado con una densidad de 1 g cm-3.
El día de la siembra se saturó con agua el sustrato contenido en ambas secciones del macetero, y una vez que aparecieron las primeras hojas verdaderas (12 días después de siembra, dds) se aplicaron 16 riegos de 250cm3 en ambas secciones (A y B) hasta después de la floración (30 dds), con la intención de estimular el crecimiento de las raíces laterales hacia estos dos partes del macetero.
Los maceteros se agruparon para conformar parcelas que se organizaron en el terreno de acuerdo a un diseño de bloques completos al azar con tres repeticiones. Cada parcela ocupó un área de 0,70 m2 y estuvo conformada por 12 maceteros de respuesta hidrogravitrópica con una semilla sembrada en cada uno; donde se evaluaron dos unidades por tratamiento. Las variables se sometieron a análisis de varianza y pruebas de comparación de medias de Tukey (p≤0,05), con ayuda del programa SAS® v9.1 [21].
● Monitorio y control de la aplicación del riego
Para monitorear la humedad del suelo durante la aplicación del riego, se instalaron en las secciones A y B de los maceteros, sensores FDR (Frequency Domain Reflectometry) EC-5 conectados a registradores de datos Em5b (DecagonDevice® USA), programados con ayuda del software ECH2O® Utility versión 1,74 con una configuración de registro horaria. Así mismo, se construyó la curva de retención agua-suelo con datos del potencial matricial del suelo (Ψms), que fueron capturados con el sensor MPS-6 (Decagon Device® USA), y datos del contenido hídrico volumétrico del suelo (Ψvs), capturados con el sensor EC-5 (Decagon Device® USA), ambos conectados al registrador de datos EM-50 (Decagon Device® USA); de esta manera se obtuvieron los parámetros del modelo de Campbell [22, 23], para finalmente poder definir los valores de Ψvsms y θvs del rango entre los que se mantuvo la humedad del suelo dentro de los límites de la fracción de agua transpirable, para que el factor controlador del estado hídrico de la planta dependa exclusivamente de las condiciones atmosféricas [24].
● Definición de los tratamientos
Los tratamientos correspondieron a riego parcial de raíces (RPR), el mismo que está diseñado para mantener la mitad del sistema radicular de las plantas sin riego (donde el suelo se puede encontrar con un nivel de humedad inferior a la capacidad de campo), mientras que al mismo tiempo la otra mitad se mantiene con riego (donde el suelo se puede encontrar a capacidad de campo), para luego intercambiar estas condiciones en la siguiente aplicación de agua; y riego completo de raíces (RCR), donde toda la sección del suelo que es explorada por las raíces se encuentra bajo el máximo régimen hídrico apropiado para las plantas. De esta forma RPR y RCR fueron combinados cada uno con dos volúmenes de agua (300 y 500 cm3), según la determinación de los límites de la fracción de agua transpirable del suelo. En este sentido, se obtuvieron cuatro tratamientos con la siguiente configuración: 1) RPR300, 2) RPR500, 3) RCR300 y 4) RCR500 (control).
Finalmente, se realizaron nueve riegos de acuerdo a la configuración de los tratamientos (con ayuda de un recipiente aforado en 300 y 500 cm3) considerando su inicio después de la floración, para evitar introducir alguna alteración en este órgano; de esta manera, el periodo de aplicación correspondió desde los 32 dds hasta los 52 dds, donde el volumen total de agua aplicada en esta etapa para cada planta/macetero fue de 900 cm3 (RCR500), 450 cm3 (RPR500), 540 cm3 (RCR300) y 270 cm3 (RPR300).
● Determinación del módulo de elasticidad y la dinámica de las relaciones hídricas en frejol común
La dinámica de las relaciones hídricas se determinó con la construcción de la curva presión-volumen (P-V) en las hojas de frejol común, siguiendo la metodología de Brown y Tanner [25]; en este sentido se monitoreo el Ψx en el momento de máxima radiación solar (entre las 11h y 14h), desde los 36 dds hasta los 43 dds en el instante anterior a la aplicación del riego y 24h después de este evento. Durante este periodo las mediciones se realizaron en la misma planta en hojas maduras no terminales (ramas del tercio medio) expuestas a radiación solar directa. Para evaluar este parámetro se utilizó una bomba Schölander (modelo 3005F01 de Soilmoisture Equipment Corp. Santa Barbara, CA), donde una hora antes de la medición las hojas fueron protegidas con bolsas plásticas aluminizadas para reflejar la luz del sol, evitar que se calienten, y conseguir el equilibrio entre el potencial hídrico de la hoja y el potencial hídrico del xilema [26] (Figura 3).

| Figura 3. | Preparación de las hojas para el monitoreo del potencial hídrico del xilema (Yx) y posterior determinación del módulo elasticidad. |
Una vez que se cortó la hoja (cubierta con la bolsa aluminizada) su peso fue registrado en una balanza analítica marca AND (modelo EJ-200, 0,01 g de precisión, Korea), luego se la introdujo en la cámara de la bomba Schӧlander donde fue presurizada a una tasa de 1 bar/2s. Con la ayuda de una lupa se observaba la parte del peciolo que queda expuesta y cuando comenzó a formarse un menisco de agua en el tejido, el ingreso de gas (nitrógeno) presurizado a la cámara fue detenido, este instante representaba el equilibro de la tensión interna del xilema con la presión del gas en el interior de la cámara; por lo tanto, el valor en el manómetro se registró como el potencial hídrico del xilema (Figura 4). Este ciclo se repitió siete veces en la misma muestra siguiendo el procedimiento de Jara y Celis [27].


| Figura 4. | Medición del potencial hídrico xilemático en la hoja de frejol común con la bomba Schlander (derecha) y formación del menisco en el peciolo de la hoja cuando se equilibran las presiones del xilema con la del gas en el interior de la cámara (izquierda). |
Después de realizada la última lectura, la hoja fue llevada al horno a 70ºC hasta que la muestra alcanzará un peso constante, de esta manera se obtuvo el peso seco (Pseco). El peso a turgor máximo (Ptm) se lo determinó por la extrapolación del potencial hídrico del xilema hacia cero (Ψx=0) en la curva que se describió entre el Ψx y Pfresco (peso fresco).
Después de la construcción de las curvas P-V para cada tratamiento, se siguió la metodología de Tyree y Hammel [28] para la determinación del potencial osmótico a turgor máximo (Ψo(CRA=1)), potencial osmótico a turgor cero (también denominado potencial hídrico xilemático letal, Ψo(turgor=0)), contenido relativo de agua a turgor cero (CRA(turgor=0)) y contenido de agua apoplástica (CAApoplástica); parámetros que también permitieron generar los diagramas de Hofler. Finalmente para determinar el módulo de elasticidad se empleó la ecuación desarrollada por Stadelmann [29], donde εv depende del potencial de turgor máximo de la hoja (Ψtm) y es inversamente proporcional al contenido relativo de agua(CRA) respectivo, junto con la inclusión de un coeficiente de regresión (β) como factor de proporción,tal como se muestra en la siguiente ecuación:
![]()
donde β está relacionado con la sensibilidad de los cambios de elasticidad y el grado de saturación de agua en las células, y puede ser calculado como el coeficiente de regresión a partir de una serie de cálculos individuales de pares de datos de potencial de turgor (Ψtn) y el contenido relativo de agua en la hoja (CRAn) con la siguiente ecuación:
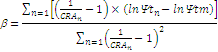
3. RESULTADOS Y DISCUSIÓN
● Control de la aplicación del agua en el riego completo y en el riego parcial
En la figura 5 se observan los patrones de humedad del suelo, generados por los nueve riegos aplicados en los secciones A y B de los maceteros, tanto con las técnicas RPR (derecha) y RCR (izquierda). En el caso de la aplicación de RPR, donde se realizaron cinco riegos en la sección A y cuatro en la sección B, se aprecia un desalineamiento entre los patrones hídricos de cada sección del macetero (Figura 5a y 5c), el mismo que si logra ser mantenido con las aplicaciones precisas y oportunas de agua de riego, se considera clave para el desencadenamiento de señales físicas y/o químicas en las raíces [30], logrando generar regulaciones que permiten mantener un adecuado régimen hídrico en toda la planta [31]. Mientras que en el caso de la aplicación de RCR, los nueve riegos se realizaron en ambas secciones al mismo tiempo (Figura 5b y 5d), efecto que permitía mantener el mismo nivel de humedad y la alineación de los patrones hídricos del suelo.
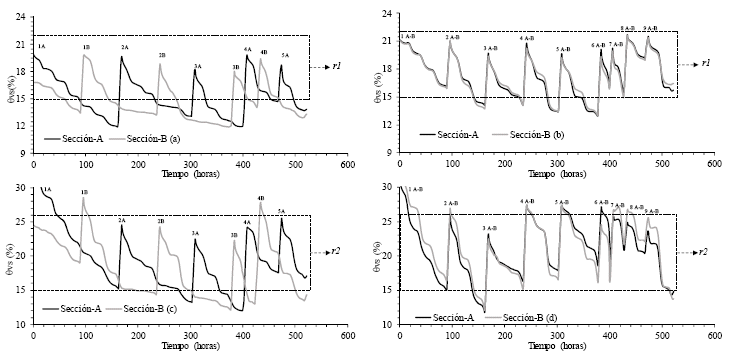
| Figura 5. | Patrones (medias móviles de seis horas) de la humedad volumétrica del suelo (θvs), originados por los nueve riegos aplicados en las secciones A y B de los maceteros de respuesta hidrogravitrópica, monitoreados con el sensor EC-5 en los tratamientos con riego parcial de raíces (RPR300(a) y RPR500(c)) y riego completo de raíces (RCR300(b) y RCR500(d)); donde r1 y r2 representan los rangos de la fracción transpirable del suelo. |
En la Figura 5 también se ha definido una región que representan los rangos de la fracción de agua transpirable del suelo; los mismos que fueron obtenidos en la curva de retención agua-suelo (Figura 6), donde se definió 26% a -41 kPa (θvs500, en los tratamientos donde se aplicó 500 cm3) y 22% a -76 kPa (θvs300, en los tratamientos donde se aplicó 300cm3) como los límites superiores para ambos rangos (siendo 28% a -33 kPa la capacidad de campo del sustrato), y 15 % a -252 kPa como el límite inferior o punto crítico (θvsc) para el caso del cultivo de frejol común [32].
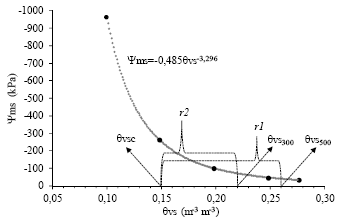
| Figura 6. | Curva de retención agua-suelo con los respectivos parámetros del modelo de Campbell (-0,485 y -3,296), generada a partir de datos (puntos negros) capturados con los sensores MPS-6 (monitoreo del Ψms) y EC-5 (monitoreo de la θvs); siendo θvsc el límite común de los rangos de la fracción de agua transpirable del suelo, definidos como r1 (θvs500-θvsc) y r2 (θvs300-θvsc), para los tratamientos donde se aplicó 500 y 300 cm3 respectivamente. |
● Dinámica de las relaciones hídricas y módulo de elasticidad en frejol común
Los diagramas de Hӧfler en los cuatro tratamientos de riego evaluados (Figura7a-7b), describieron la dinámica de las relaciones hídricas en todos los niveles de la hoja de frejol común, a medida que perdió o ganó turgencia (entre un momento antes de aplicar el riego y 24h después de su aplicación, respectivamente), entre los límites máximos de hidratación y deshidratación para cada momento (emulado en el proceso de construcción de las curvas P-V).
En este trabajo solo se muestran los diagramas de Hӧfler que fueron determinados con las mediciones realizadas antes del riego (ar, 20-11-2014) y 24h después del riego (24hdr, 21-11-2014) para ambas configuraciones de volúmenes de agua (300 cm3, figura 7a y 500cm3, figura 7b); donde se aprecia que la intercepción del eje de los potenciales (turgor, xilemático y osmótico) sobre el eje del contenido relativo de agua (CRA), se ubicó en el punto que define el inicio dela plasmólisis, también conocido comoel inicio de la pérdida de turgorfoliar(CRA(turgor=0)).
En este sentido, los diagramas de Hӧfler (Figuras 7a y 7b) permitieron observar como la variación y respuesta elástica del tejido es provocada por el mantenimiento de diferentes niveles de agua aplicados al suelo(según la curva de retención agua-suelo). Así mismo, autores que han observado el mismo fenómeno expresan que aunque no está demostrada la ventaja ecofisiológica de tener tejidos más elásticos o menos elásticos, indican que el déficit hídrico es el responsable de las restricciones elásticas en las paredes celulares [33, 34].
Del mismo modo, se dice que las variaciones elásticas que se producen en el corto tiempo podrían tener un mayor origen por cambios en el hidrogel de pectina de la pared celular, que por modificaciones en la red de fibras de celulosa [35]. Este efecto puede ser observado en las pequeñas amplitudes que el εv mantiene entre el momento antes del riego y 24h después del riego, principalmente, en los tratamientos donde se aplicó un volumen de agua que permitió mantener el contenido de humedad en el suelo cercano a capacidad de campo (Figura 7b). Por otro lado, la restricción de agua en los tratamientos con una configuración de 300 cm3 (para el momento antes de la aplicación del riego) pudo provocar rigidez en las paredes celulares del tejido, tal como lo revelan los altos valores del εv, principalmente en RPR300 (Figura 7a).
En general, si se compara gráficamente el desempeño elástico del tejido en el momento anterior a la aplicación del riego, se puede observar que el manejo adecuado de una técnica de riego reducido (tratamiento RPR500) puede lograr mantener similares o más bajos valores de εv(14,83 MPa), aun, cuando su CRA(turgor=0)es menor (0,87); en comparación a la utilización de una técnica convencional (tratamiento RCR500, εv=14,91 MPa y CRA(turgor=0)=0,91) tal como se aprecia en la figura 7b. Aun así, hay estudios indicando que un tejido más elástico puede mejorar muy poco su turgencia por aumentos de agua simplástica proporcionalmente grandes [36], pero sin embargo, estaría sujeto a perder turgencia solo con pérdidas apreciables de agua en el simplasto (caso RPR500ar); por otro lado, un tejido menos elástico puede mejorar mucho su turgencia con pequeños aumentos de agua en el simplasto, pero también puede empeorar mucho su situación con una pérdida equiparable de agua (caso RCR500ar).
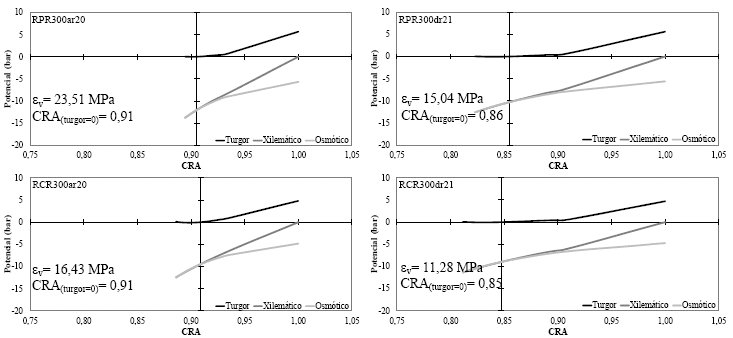
| Figura 7a. | Diagramas de Hӧfler con el respectivo valor del módulo de elasticidad (ev) y contenido relativo de agua a turgor cero (CRA(turgor=0)) de las hojas de plantas de frejol común cultivadas en maceteros de respuesta hidrogravitrópica donde se aplicó 300 cm3 de agua en las secciones respectivas, según los tratamientos RPR (superior) y RCR(inferior) antes del riego (izquierda, 20-11-2014) y 24h después del riego (derecha, 21-11-2014). |
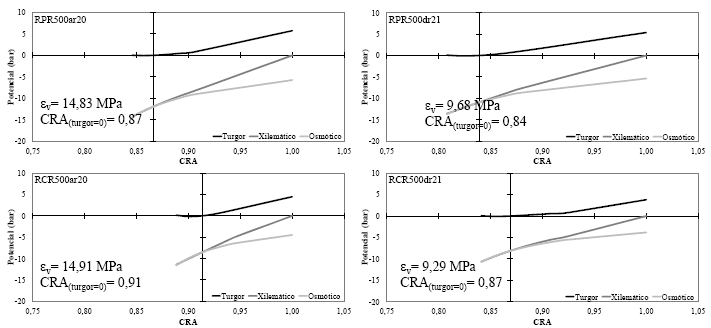
| Figura 7b. | Diagramas de Hӧfler con el respectivo valor del módulo de elasticidad (εv) y contenido relativo de agua a turgor cero (CRA(turgor=0)) de las hojas de plantas de frejol común cultivadas en maceteros de respuesta hidrogravitrópica donde se aplicó 500 cm3 de agua en las secciones respectivas, según los tratamientos RPR (superior) y RCR (inferior) antes del riego (izquierda, 20-11-2014) y 24h después del riego (derecha, 21-11-2014). |
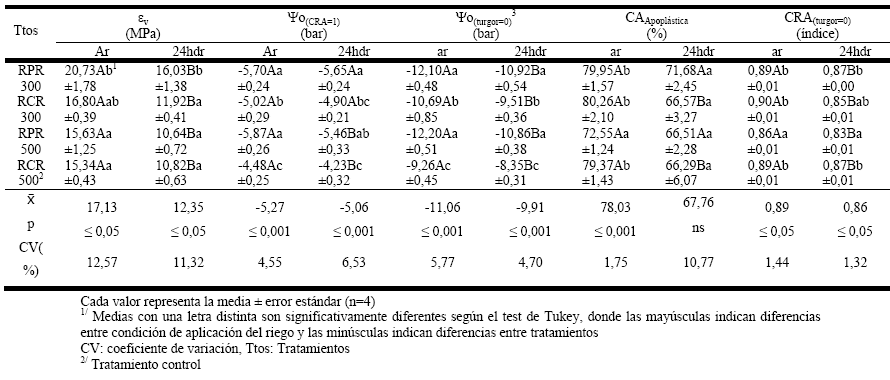
Por otro lado, en la Tabla 1 se aprecia que los valores de contenido de agua apoplástica (CAApoplástica) fueron mayores antes de la aplicación del riego que 24h después, esto es debido al momento de deshidratación que están atravesando las plantas; en este sentido, dentro de los períodos previos a la aplicación de riego, un bajo valor en el CAApoplástica indica que se está estimulando un mayor ingreso de agua desde el apoplasto (72,55 %) hacia el simplasto (27,45 %) en el tratamiento RPR500, cuando se compara con el tratamiento RCR500 (79,37 % en el apoplasto y 20,63 % en el simplasto); lo que evidencia un mecanismo de evasión a la deshidratación celular, motivado por la disminución del potencial osmótico (Ψo(CRA=1)). De esta forma, varios autores en diferentes cultivos también han indicado que por causas debidas al déficit hídrico y otras variables abióticas, las plantas disminuyen su potencial osmótico para impulsar la entrada de agua a las células y evitar la deshidratación [3, 34, 36, 37].
En el mismo sentido, se aprecia que 24h después de aplicar el riego el contenido de agua apoplástica tendió a disminuir (y por ende el contenido de agua en el simplasto tendió a incrementarse) lo que evidencia una tendencia de conseguir un equilibrio entre los dos espacios, por la hidratación a la que son expuestas las células cuando las plantas son irrigadas (principalmente en los tratamientos bajo RCR).
En el mismo contexto, también se aprecia que después del riego (24hdr) en todos los tratamientos aumentó la elasticidad del tejido (al disminuir el valor de εv); sin embrago, dentro de este momento en RPR300 el tejido de las hojas fue el más inelástico (valor de εv más alto). Así mismo, en los tratamientos bajo riego parcial de raíces (antes de la aplicación del riego) solo RPR500 llegó a tener el más bajo valor de εv y ser estadísticamente tan elástico como el control. Mientras que en el caso de Ψo(CRA=1) sus niveles pueden indicar la presencia de un ajuste osmótico en los tratamientos bajo RPR, porque sus valores fueron significativamente más negativos que en el tratamiento control (Tabla 1). En el mismo escenario, hay autores que no encontraron evidencias de ajuste osmótico ni elástico en el cultivo de pera (Pyruscommunis L.) cuando fue sometido a riego deficitario controlado [37]; sin embargo, las diferencias significativas encontradas en el contenido de agua apoplástica sugieren un mecanismo de adaptación al déficit de agua.
En este trabajo, se puede afirmar que en el tratamiento RPR500 el otro mecanismo de adaptación o evasión a la deshidratación (observado en el momento antes del riego), correspondió al mantenimiento de un potencial hídrico letal muy bajo (Ψo(turgor=0)), lo que permitió que un disminuido contenido relativo de agua (cuando su potencial de turgor es igual a cero, CRA(turgor=0)) mantenga en mejores condiciones hídricas a las plantas de frejol común en comparación con el tratamiento sin restricción de agua (RCR500).
| TABLA 1. | PARÁMETROS DE LA CURVA PRESIÓN-VOLUMEN EN LA HOJA DE FREJOL COMÚN ANTES DE LA APLICACIÓN DEL RIEGO (AR) Y 24H DESPUÉS DE SU APLICACIÓN (24HDR) DE OCHO EVALUACIONES REALIZADAS EN LOS CUATRO TRATAMIENTOS CON RIEGO PARCIAL (RPR) Y RIEGO COMPLETO (RC) DE RAÍCES, EN PLANTAS CULTIVADAS EN MACETEROS DE RESPUESTA HIDROGRAVITRÓPICA SELECTIVA. |
4. CONCLUSIONES
El control de la aplicación del riego con la curva de retención agua-suelo, permitió manejar adecuadamente los niveles de humedad en el suelo, efecto que pudo ser aprovechado por las plantas para desencadenar fluctuaciones significativas en los niveles del módulo de elasticidad (εv);tanto en las plantas bajo riego completo de raíces como en aquellas bajo riego parcial de raíces.
La aplicación de volúmenes de agua correspondiente a valores que llevaron al suelo a un nivel cercano a la capacidad de campo (tratamientos RPR500 y RCR500), incrementó significativamente la elasticidad del tejido de las plantas; sin embargo, solo bajo RPR500 se pudo observar un mayor desempeño elástico, debido a los ajustes que pudieron realizar las plantaspor la exposición de sus raíces cuando se combinan de forma controlada las reducciones de agua aplicadas al suelo junto con el mantenimiento de dos niveles diferentes de humedad en un mismo momento.
5. AGRADECIMIENTOS
El primer autor agradece a SENESCYT-Ecuador por la beca 2013-AR4P139 otorgada para los estudios en el PDRH-UNALM (Lima-Perú), y a la Universidad Técnica de Manabí (Ecuador) por el financiamiento de la bomba Schӧlander, a través del proyecto 2012-026IIC.
6. BIBLIOGRAFÍA
[1] G. Gomes, A. Moritz, G. Freiria, F. Furlan, e L. Assari, Desempenho produtivo de genótipos de feijão-vagem arbustivo em dois ambientes, Scientia Agropecuaria, vol. 7, No.2, pp 85-92, 2016.
[2] S. Cuéllar, y A. Covarrubias, Alternativas para enfrentar la sequía en el cultivo de frijol (Phaseolusvulgaris L.), Revista Claridades Agropecuarias, vol. 142, pp 32-41. 2005.
[3] J. Martínez, H. Silva, J. Ledent, and M. Pinto,Effect of drought stress on the osmotic adjustment, cell wall elasticity and cell volume of six cultivars of common beans (Phaseolus vulgaris L.), Eur. J. Agron., vol. 26, no. 1, pp 30-38.2007.
[4] C. Pimentel, Respostas das plantas à seca, enA relação da planta com a água. EDUR -Editora Universidade Federal Rural do Rio de Janeiro, Soropédica-RJ, 2004, 122p.
[5] J. Jiménez, y J. Acosta, Rendimiento de frijol común (Phaseolusvulgaris L.) yTépari (Phaseolusacutifolius A. Gray) bajo el método riego-sequía en Chihuahua, Revista Mexicana de Ciencias Agrícolas, vol. 4, no. 4, pp 557-567, 2013.
[6] M. Behboudian, and Z. Singh, Water relations and scheduling in grapevine, Horticultural Reviews, vol. 27, pp 189-225, 2001.
[7] E. Vega, Desempeño del frijol común bajo riego parcial de raíces en un sistema de respuesta hidrogravitrópica selectiva,. Tesis doctoral, Universidad NacionalAgraria La Molina, Lima-Perú, 2016. [ Links ]
[8] J. Zegbe, M. Behboudian, A. Lang, and B. Clothier, Responses of Petopride processing tomato to partial rootzone drying at different phenological stages, Irrigation Science, vol. 24, pp 203-210, 2006.
[9] J. Fernández, A. Díaz-Espejo, J. Infante, P. Durán, J. Palomo, V. Chamorro, I. Girón, and L. Villagarcía, Water relations and gas exchange in olive trees under regulated deficit irrigation and partial rootzone drying, Plant and Soil, vol. 284, no. 1, pp. 273-291, 2006.
[10] P. Romero, and A. Martinez-Cutillas, The effects of partial root-zone irrigation and regulated deficit irrigation on the vegetative and reproductive development of field-grown Monastrell grapevines, Irrigation Science, vol. 30, no. 5, pp.377–396, 2012.
[11] F. Yan, S. Yanq, F. Song, F. Liu, Differential responses of stomatal morphology to partial root-zone drying and deficit irrigation in potato leaves under varied nitrogen rates, ScientiaHorticulturae, vol. 145, pp. 76–83, 2012.
[12] R. Nunes, F. Mazzei, A. Oliveira, B. Corrêa da Silva, T. Massi, M. de Menezes, E. Fernandes, D. Michael, and E. Campostrini, Partial rootzone drying (PRD) and regulated deficit irrigation (RDI) effects on stomatal conductance, growth, photosynthetic capacity, and water-use efficiency of papaya, ScientiaHorticulturae, vol. 183, pp. 13-22, 2015.
[13] A. Sepaskhah, S. Ahmadi, A review on partial root-zone drying irrigation, International Journal of Plant Production, vol. 4, no. 4, pp. 241-258, 2015.
[14] I. Abrisqueta, W, Conejero, M Valdés-Vela, J. Vera, M. Ortuño, and M. Ruiz-Sánchez, Stem water potential estimation of drip-irrigated early-maturing peach trees under Mediterranean conditions, Computers and Electronics in Agriculture, vol. 114, pp. 7-13, 2015.
[15] B. Goldenbogen, G. Wolfgang, M.Hemmen, J.Uhlendorf, A. Herrmann, and E.Klipp Dynamics of cell wall elasticity pattern shapes the cell during yeast mating morphogenesis, Open Biol, vol. 6, no. 160136, pp 1-14, 2016.
[16] T. Du, S. Kang, J. Zhang, W. Davies, Deficit irrigation and sustainable water-resource strategies in agriculture for Chinas food security, J. Exp. Bot., vol. 66, no. 8, pp. 2253-2269, 2015.
[17] N. Arabzadeh and S. Emadian, Effect of water (drought) stress on water relations of Haloxylonaphyllum and H. persicum, Iranian Journal of Science & Technology, Trans.A, vol. 34, no.A3, pp 245-255, 2010.
[18] M. Gijón, C. Gimenez, D. Perez-López, J. Guerrero, J.Couceiro, and A.Moriana, Water relations of pistachio (Pistaciavera L.) as affected by phenological stages and water regimes, ScientiaHorticulturae, vol. 128, pp. 415-422, 2011.
[19] K. Hessini, J. Martínez, M. Gandour, A. Albouchi, A. Soltani, and C. Abdelly, Effect of water stress on growth, osmotic adjustment, cell wall elasticity and water-use efficiency in Spartinaalterniflora, Environ. Exp. Bot., vol. 67, no. 2, pp. 312-319, 2009.
[20] N. Saruhan, A. Sağlam, M. Demiralay, and A. Kadioğlu, Apoplastic and symplastic solute concentrations contribute to osmotic adjustment in bean genotypes during drought stress, Turk J. Biol., vol. 36, pp. 151-160, 2012.
[21] SAS Institute Inc. User´s Guide, SAS/STAT® v9.1, 2004. [ Links ]
[22] M. Bittelli and M. Flury, Errors in water retention curves determined with pressure plates, Soil Science Society of America Journal, vol. 73, no. 5, pp. 1453-1460, 2009.
[23] M. Khlosi, Performance evaluation of models that describe the soil water retention curve between saturation and oven dryness, Vadose Zone Journal, vol. 7, pp. 86-97, 2008.
[24] A. Pardossi, L. Incrocci, G. Incrocci, F. Malorgio, P. Battista, L. Bacci, B. Rapi, P. Marzialetti, J. Hemming, and J.Balendonck, Root zone sensors for irrigation management in intensive agriculture, Sensors, vol. 9, pp. 2809-2835, 2009.
[25] P. Brown and C. Tanner, Alfalfa osmotic potential: A comparison of the water-release curve and frozen-tissue methods, Agron. J., vol.75, pp. 91-93, 1983.
[26] P. Romero, J. Pérez-Pérez, F. del Amor, A. Martínez-Cutillas, I. Dodd, and P. Botía, Partial root zone drying exerts different physiological responses on field-grown grapevine (Vitisvinifera cv. Monastrell) in comparison to regulated deficit irrigation, Functional Plant Biology, vol. 41, no. 11, pp. 1087-1106, 2014.
[27] J. Jara y J. Celis J, Módulo de elasticidad y otros parámetros biofísicos en frijol negro, Agro-Ciencia, vol. 5, no. 1, pp. 49-56, 1989.
[28] M. Tyree and T. Hammel, The measurement of the turgor pressure and the water relations of plants by the pressure-bomb technique, J. Exp. Bot., vol. 23, no. 1, pp. 267-282, 1972.
[29] E. Stadelmann, The derivation of the cell wall elasticity function from the cell turgor potential, J. Exp. Bot., vol. 35, no. 6, pp. 859-868, 1984.
[30] J. Comstock, Hydraulic and chemical signaling in the control of stomatal conductance and transpiration, J. Exp. Bot., vol. 53, pp. 195-200, 2002.
[31] K. Prado, and C. Maurel, Regulation of leaf hydraulics: from molecular to whole plant levels, Frontiers in Plant Science, vol. 4, no. 255, pp. 1-14, 2013.
[32] F. Camarena, A. Huaringa, y E. Mostacero, Manejo agronómico en Tecnología para el incremento de la producción de frijol común (Phaseolusvulgaris L.). Primera edición. Universidad nacional Agraria La Molina – Consejo Nacional de Ciencia, Tecnología e Innovación Tecnológica. Lima-Perú, 153 p.
[33] B. Dichio, C. Xiloyannis, K. Angelopoulos, V. Nuzzo, S. Bufo, and G. Celano, Drought-induced variations of water relations parameters in Oleaeuropaea, Plant Soil, vol. 257, no. 2, pp. 381-389, 2003.
[34] A. Patakas and B. Noitsakis, Cell wall elasticity as a mechanism to maintain favorable water relations during leaf ontogeny in grapevines, Am. J. Enol.Vitic., vol. 48, pp. 352–356, 2002.
[35] H. Félix, Regulación de la hidratación y la turgencia foliares por mecanismos evitadores del estrés, y resistencia a déficit hídrico en vid: modelo vs. Experimentos, Tesis doctoral, Universidad Nacional de Cuyo, Mendoza-Argentina, 2011.
[36] M. Bolarin, M. Estañ, M. Caro, R. Romero-Aranda, and J. Cuartero, Relationship between tomato fruit growth and fruit osmotic potential under salinity, Plant Sci, vol. 160, no. 6, pp. 1153-1159, 2001.
[37] D. Díaz-Abril, J. Vélez-Sánchez, and P. Rodríguez, Irrigation reduction resistance mechanisms in the rapid fruit growth stage of pears (Pyruscommunis L.), Agron. colomb., vol. 24, no. 1, pp. 25-32, 2016.

