











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
INTRODUCCIÓN
Las especies del género de Masdevallia (subtribu Pleurothallidinae), se distribuyen desde sur de México al sur de Brasil, con mayor diversidad en Los Andes de sur América desde Venezuela, Colombia, Ecuador, Perú y Bolivia (Abele, 2007). El género Masdevallia, es uno de las más diversas (50 especies). En Bolivia existen aproximadamente 1 500 especies de orquídeas. El 80 % de las especies de Masdevallia son endémicas (Vásquez y Ibisch, 2000). La mayoría de las orquídeas bolivianas están bajo algún grado de amenaza, debido a que sus poblaciones son generalmente muy pequeñas, frágiles y de hábitat restringido. Sin embargo, la información científica sobre estas, así como del resto de las orquídeas bolivianas es casi inexistente.
La belleza de las orquídeas ha resultado, en que muchas son de valor comercial, como ornamental, tal el caso de M. solomonii. Esta especie y al parecer todas del grupo de orquídeas, se destacan por presentar complejas interacciones dentro del ciclo de su vida, como su relación obligada con hongos micorrízicos y que poseen sistemas de polinización muy especializados (Neiland y Wilcock, 1998). Una posible consecuencia en M. solomonii, es que su distribución es muy limitada, lo cual recaería en una mayor vulnerabilidad a la dependencia de sus interacciones bióticas con los polinizadores y las micorrizas. Esta hipótesis, se plantea debido a que, en su hábitat natural, la producción de frutos en orquídeas está por debajo del 20 %, y se estima que tan solo llegan a germinar entre 2 a 3 % en presencia del hongo micorrizíco (Singh, 1988).
El conocimiento de la ecología básica de las especies con potencial económico, como es M. solomonii es fundamental al momento de plantear estrategias de manejo, como la fenología que está relacionada a la biología reproductiva. La fenología no solo estudia la fase vegetativa y reproductiva, sino que incluye el clima, la estación, los cambios de un área particular (Leith y Radofor, 1971 citado por Lokho y Kumar, 2012). También, la fenología incluye la interacción entre plantas y animales (polinizadores) (Parra y Vargas, 2004).
En la naturaleza, las orquídeas son generalmente polinizadas por insectos, los cuales pueden llevarse casi todo el polen en una sola visita, ya que estos vienen agrupados en paquetes (polínios); aunque también se da la autopolinización en bajo porcentaje (menor al 5 %) (Neiland y Wilcock, 1998). Después de la fertilización ocurrirá el desarrollo y formación del embrión (embriogénesis) y de las otras estructuras de las semillas. El tiempo que requieren las semillas de orquídeas para madurar es muy variable. Por ejemplo, las especies del género Dendrobium y Phalenopsis necesitan de cuatro meses, mientras que Cattleya y Paphiopedilum requieren entre 10 a 12 meses y Vanda hasta 15 meses (Richter, 1969).
La simbiosis micorrízica entre la orquídea y el hongo depende de la distribución y densidad de los hongos en el ambiente, lo cual influye en la dinámica poblacional de las orquídeas, ya que es posible la existencia de una alta especificidad de asociación entre orquídeas y micorrizas (Rivas et al., 1998). En condiciones naturales la infección del hongo, que es uno de los factores determinantes para garantizar la germinación, está condicionada por otros factores como la disponibilidad de agua, temperatura, pH, sales minerales, vitaminas y reguladores de crecimiento (Pierik, 1990). El proceso de germinación se estima que tarda hasta tres años (Devesa, 1997), lo cual hace muy difícil su estudio en condiciones naturales (Bayman et al., 2002). Sin embargo, es posible disminuir este tiempo e incrementar las probabilidades de germinación de orquídeas a través del uso de herramientas biotecnológicas, suministrando a las semillas todos los nutrientes que necesitan para su crecimiento (Rodríguez et al., 2005), lo cual se logra mediante técnicas de biotecnología (Morales, 2011).
Los estudios de germinación in vitro en otras especies del género Masdevallia (M. chaparensis, M. exquisita, M. tovarensis, y M. auripurpurea) y distintos medios de cultivo, reportan porcentajes de germinación de 42, 53, 8.5 y 78 % respectivamente en los medios de cultivo de Murashige y Skoog; Knudson C, Hidro-Coljapp y medio básico (BA + ANA), respectivamente (Villegas, 2003; Sánchez, 2007; Michelangeli, 2010; Pedraza, 2011). Otros factores que se cree pueden afectar en la germinación son la calidad de luz recibida y el fotoperíodo, ya que algunas especies requieren de la luz para la germinación, mientras que otras pueden germinar en la oscuridad (Dutra et al., 2009).
El objetivo de la presente investigación fue caracterizar la fenología vegetativa y reproductiva de Masdevallia solomonii a lo largo de un ciclo anual en sus poblaciones naturales y determinar el efecto de la intensidad lumínica sobre la germinación in vitro.
MATERIALES Y MÉTODOS
Ubicación del área de estudio
El trabajo fue realizado en el camino precolombino de Chojllapata, se encuentra en el Parque Nacional y Área Natural y de Manejo Integrado Cotapata (PN- ANMI Cotapata) del departamento de La Paz en Bolivia (Figura 1). La zona de estudio corresponde, a un bosque montano nublado de Yungas representa la formación vegetal más compleja y diversa, se relaciona con los bosques húmedos de montaña (Kessler y Beck, 2001). Geográficamente ubicada a 67º 52' 35.947'' longitud oeste y 16º 14' 52.43'' latitud sur, a 2 859 m s.n.m.
Metodología
En el bosque montano superior, son abundantes las epífitas no vasculares como los musgos Campylopus, Grimmia, Anacolia, Leptodontium, Calyptohecium, Sphagnum y Racomitrium y los líquenes, Cladonia y Cora, así como las vasculares, que aumentan en los pisos inferiores, con varias especies de helechos y afines (licopodios y selaginelas), aráceas, bromeliáceas y orquidáceas. Los árboles dominantes son: Clethra, Clusia, Vismia, V. glabra y V. glaziovii, Hieronima, Alchornea, A. pearcei, Escallonia paniculata, Freziera, Gaultheria, Hedyosmum, Miconia, Oreopanax, Podocarpus, Symplocos y Weinmannia. Las palmeras son escasas y están representadas por individuos de los géneros Ceroxylon y Geonoma. En algunos sectores del bosque se encuentran los llamados “inciensales” con Clusia cf. lechleri. En laderas algo secas con vegetación pionera se tienen manchas de bosque semideciduo con Alnus acuminata (aliso) (Kessler y Beck, 2001).
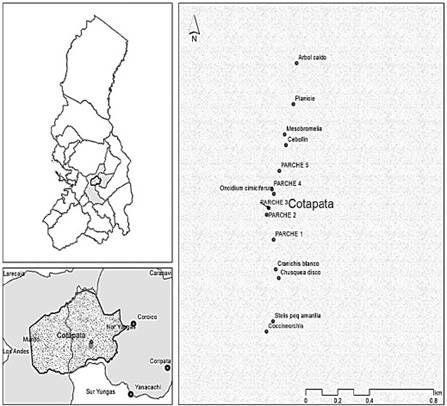
Figura 1 Localización del área de estudio, con demarcación de las subpoblaciones distribuidas en cinco parches en un trayecto del camino precolombino Chojllapata del PN-ANMI, Cotapata (Fuente: Mendoza, 2010).
A lo largo del camino precolombino Chojllapata, entre rangos altitudinales desde 2 914 a 2 878 m, se registró cinco parches de M. solomonii, se entenderá como parche, a una subpoblación, donde los individuos se encuentran agrupados y presentan una distribución restringida marcada a lo largo de un gradiente altitudinal. En el primer parche (Figura 2) se registró 47 individuos, a diferencia de los demás que se observó menor cantidad de individuos de 26, 7, 6 y 10 en segundo, tercer, cuarto y quinto parche. Además, en el primer parche se observó que M. solomonii compartía el hábitat con otras especies de orquídeas del mismo género, como M. scandens y M. yungasensis, y otras especies Epidendrum y Pleurothalis.

Figura 2 Especies de Masdevallia presentes en el parche 1. a, M. solomonii. b, M. yungasensis. c, M. scandens.
Seguimiento fenológico
Entre diciembre de 2008 y noviembre de 2009 se realizaron salidas mensuales por el camino precolombino Chojllapata. A lo largo de dicho camino se marcaron y georeferenciaron cinco parches con individuos de M. solomonii y se codificaron a todos los individuos fértiles de la especie en estudio. El seguimiento fenológico se realizó en base a fenofases reproductivas de individuos fértiles (con botones florales, flores y frutos) y estériles (que se encontraban en estado vegetativo). También se identificaron y se describieron fenofases del desarrollo y maduración de los frutos tal como sugieren Moya et al. (2007).
Colecta de frutos y siembra en condiciones controladas
Se colectaron cinco frutos de M. solomonii de diferentes plantas y fueron guardados en sobres de papel aluminio y llevados a la Unidad de Biotecnología Vegetal del Instituto de Biología Molecular y Biotecnología de la Universidad Mayor de San Andrés. Los frutos fueron secados bajo temperatura ambiente en sombra, hasta que liberen las semillas de manera natural, en un tiempo que duró entre 20 a 30 días. Estas semillas permanecieron almacenadas durante dos meses a 4 ºC y posteriormente, se realizó una mezcla de las semillas para obtener una muestra homogénea y representativa de la especie. Posteriormente, las semillas fueron desinfectadas con hipoclorito de sodio al 0.5 % durante 10 minutos con una gota de detergente líquido. Dentro la cámara de flujo laminar las semillas fueron vertidas en el aparato de filtración, donde se enjuagó con agua destilada estéril, seguidamente se colocaron en otro frasco en agua destilada estéril con la finalidad de obtener una solución con semillas. Con una jeringa se colocó 4 ml (la solución con semillas) en cada tubo de ensayo que contenía el medio de cultivo Knudson (1946) y se procedió al sellado del tubo de ensayo (Figura 3), primero con papel aluminio y después con plastifil. Finalmente, los tubos fueron llevados a la cámara de crecimiento, considerando a cada tubo como la unidad experimental.

Figura 3 Procedimiento de siembra de semillas. a, desinfección y enjuague de semillas de M. solomoni. b, filtrado de semillas en papel filtro. c, elaboración de una suspensión de agua con semillas. d, siembra en medios de cultivo.
Diseño experimental
Para el porcentaje de individuos en cada fenofases vegetativa y reproductivas en M. solomonii fue evaluado en función al número de registro en cada fenofase en relación al total de individuos registrados. Mientras que, las semillas que fueron sometidas a dos tratamientos de intensidad durante la germinación en la cámara de crecimiento que consistieron en: a) 1.965 µMol m-2s-1 (baja intensidad = T1) y b) 12 a 7.992 µMol m-2s-1 (alta intensidad = T2) para el análisis se empleó el T-Student (Calzada, 1970).
Variables de respuesta
Descripción de las fenofases reproductiva: entre diciembre de 2008 a noviembre de 2009 se realizó el seguimiento fenológico de manera mensual, donde se registró el número de individuos en estado vegetativo (estériles) en estado reproductivo (botones florales, flores y frutos).
Porcentaje de frutos formados: se cuantificó el número frutos que lograron madurar a las cuales se consideró procedentes de polinización natural.
Tamaño y diámetro del fruto: con un vernier se tomó medidas del diámetro y longitud de los frutos maduros al cabo de ocho meses.
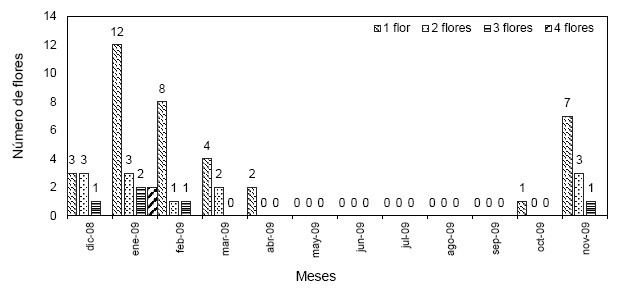
Porcentaje de germinación: se evaluó a las 18 semanas el total de semillas germinadas, y para ello se consideró, las semillas que pasaron las fases:
Fase 3, el embrión da lugar a una estructura protocormal la cual va adquiriendo diferentes formas la misma que presenta hinchamientos a manera de brotes apicales en todo el contorno de su superficie, en orquídeas es considerado que la semilla ha germinado; Fase 4, se caracteriza por la aparición de un brote meristemático, el cual se torna de color verde intenso, mientras que el protocormo se torna blanco; Fase 5, el surgimiento de hojas y en el extremo opuesto al brote meristemático se da el inicio de la emergencia de rizoides.
RESULTADOS Y DISCUSIÓN
Seguimiento fenológico
El seguimiento fenológico se realizó en el parche 1, donde se registró mayor cantidad de individuos de M. solomonii. El pico de floración se registró en enero y la proporción de individuos en flor fue disminuyendo gradualmente hasta decaer por completo entre mayo y septiembre. La producción de frutos aumentó a medida que disminuyó la floración y mostró su mayor pico en mayo. Los botones florales empezaron a aparecer en septiembre y el mayor pico se registró en noviembre. De la misma manera, se evidenció que desde mayo a octubre presentaron más del 40 % de la población en estado vegetativo (estéril), estos resultados concuerdan con el descenso de las fases reproductivas (botones florales, flores y frutos). Registrándose una mayor proporción de individuos estériles en octubre (Figura 4).
Generalmente la presencia de botones florales y flores en orquídeas se presenta en época lluviosa. En base a registros de datos climáticos del centro meteorológico “Chuspipata” que encuentra cercano al área de estudio, geográficamente localizado a 16° 18' 01'' Latitud S 67° 48' 50'' Longitud O a 3 000 m (Molina-Carpio, 2019). Contrastando, datos climatológicos con los patrones fenológicos, el mayor pico de floración de M. solomonii se presentó en enero, con mayor precipitación (650 mm) y con una temperatura que está por encima de 12 °C. Al respecto, Robbirt et al. (2012) indica que un incremento de 1 °C, en 12 de 15 especies de orquídeas favorece en la floración. En concordancia, a esta afirmación se observa en los registros de datos de climáticos de trabajo de Molina-Carpio (2019), desde noviembre a febrero la temperatura tiende hacer mayor a 12 ºC, y es en estos meses cuando se presentaron los botones florales y flores en M. solomonii. También, los resultados son corroborados por Rathcke y Lacey (1985) citados por Robbirt et al. (2012) quienes indican que los tres factores ambientales principales influyan en la floración son temperatura, fotoperíodo y precipitación.
La fructificación en M. solomonii se presentó con el descenso de la precipitación en los meses de mayo, junio, julio y agosto, de 171, 121, 89 y 144 mm respectivamente. Así mismo, se evidencia que la temperatura desciende por debajo de los 11 ºC (Molina-Carpio, 2019).
Además, es importe mencionar que el área de estudio, en el sotobosque donde se encontró los individuos de M. solomonii, hay especies arbóreas del género Clusia, cuyos forófitos están cubiertos de musgos, es donde están establecidos la especie en estudio y con poca frecuencia ingresa claros de luz, razón por la cual el ambiente casi siempre está húmedo. El hecho, que reduzca la precipitación en la época de maduración del fruto es propicio, de lo contrario podría generar la pudrición y no concluir la formación de semillas. Al respecto, Molina-Carpio (2019) señala que la evapotranspiración potencial es igual evapotranspiración real, lo que corresponde a un clima hiperhúmedo, ninguna deficiencia de agua y el suelo está saturado a lo largo del año.
Los patrones fenológicos en M. solomonii coinciden en otras especies de orquídeas, en un estudio realizado por Ordoñez y Rarrado (2017) con Cyrtochilum revolutum, Malaxis excavata, Ponthieva diptera y Stelis pulchella determinan que el incremento de las lluvias se asoció con una mayor floración, las cuales se encuentran en un bosque altoandino de Colombia. Al igual que Lokho y Yogendra (2012) en Dendrobium indican que el pico de floración comienza durante la primavera y a principios de verano, que coinciden con los meses de mayor precipitación (marzo a julio). Similares patrones estacionales se presentaron en Masdevallia coccinea con un pico de floración entre junio y julio (mayor precipitación) y no así presenta un efecto directo de la fructificación con la precipitación (Matallana-Puerto et al., 2022).
No obstante, en otra especie de Masdevallia ígnea Rchb no reporta una relación directa de la precipitación con el mayor pico de floración dentro del Jardín botánico José Celestino Mutis, esto probablemente se deba que esta área fue adecuada para la exhibición de flora nativa resguardada y no está en su hábitat natural. Al igual que, en Ansellia africana aprovecha las lluvias para potenciar su crecimiento vegetativo y posteriormente florece en septiembre cuando las temperaturas son altas y las precipitaciones escasas, con el correspondiente aumento de la humedad relativa (Essomo et al., 2016).
Además, la fenología depende del tipo de hábitat que proporciona el entorno, es así que en un bosque caducifolio (se encuentra a 35° 32' 34.92'' S y 71° 11' 11.16'' O, en orquídeas Gavilea venosa, G. odoratissima y Codonorchis lessonii de habito terrestre. La floración coincide con la estación cálida de primavera y termina posterior a la sequía de las hojas en verano (Bravo et al., 2012).
Número de flores por planta
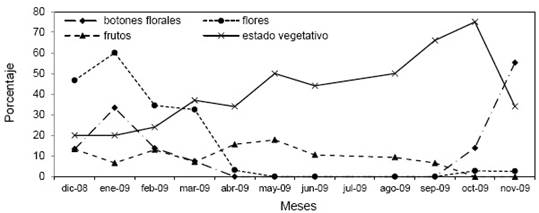
Durante los siete meses que se presentó la floración en M. solomonii (Figura 5) en la mayoría de los casos, por individuo llegaron a formar una flor y pocos tenían más de dos flores y a través de observaciones en campo se constató las plantas que tenían tres flores no tenían polinios viables, de los cuales se evidenció las flores no llegaron a formar frutos.
En la Figura 5 se observa que mayor número de flores por individuo se presentó en enero, coincidentemente fue de mayor precipitación. Inouye (2002) señala que una vez que se inicia la floración, la cantidad de precipitación durante la temporada de crecimiento puede afectar el número de flores y la duración de la floración de cualquier especie. Al respecto, en Masdevallia coccinea, la precipitación se relacionó de manera positiva con el número de flores (Matallana- Puerto et al., 2022). En M. solomonii se evidencia el número de flores es muy reducida en relación a M. ígnea, donde se registraron 12.32 flores por planta (Chavarro et al., 2005). Lo cual da indicios que M. solomonii es una especie de mucha preocupación, debido a que tiene un 13 % de producción de frutos, con reducido número de individuos en los cinco parches en el camino precolombino Chojllapata del PN-ANMI Cotapata. También mencionar que un proyecto de fenología en Masdevallia en Cotapata, los investigadores reportaron otro parche de Masdevallia en el camino precolombino Sillutinkara (Moya et al., 2007). Sin embargo, en el año 2010 se presentó torrenciales lluvias y se deslizó el sendero del camino, este camino es ampliamente usado con fines de transitabilidad de turistas, y se observó alteraciones en el entorno natural, y no se logró observar la misma cantidad de individuos de Masdevallia.
Descripción del desarrollo de los frutos
Durante el desarrollo y formación de los frutos de M. solomonii se llegó a distinguir las siguientes fases (Figura 6), el tiempo que toma en desarrollar y madurar los frutos fue de ocho meses.
a) Fase 1: se inicia cuando el ovario va aumentando de volumen y longitud como resultado de fecundación exitosa y otra de las particularidades es que aún la flor permanece abierta, de color característico de la especie y las caudas (sépalos modificados) presentan un ángulo de separación mayor a 90º.
b) Fase 2: el ovario va aumentando en grosor principalmente en la parte superior del fruto y así mismo, se evidencia los sépalos de la flor se van cerrando poco a poco; pero, aún se mantiene gran parte del resto floral y este va perdiendo color su característico.
c) Fase 3: durante este momento los sépalos y las caudas inferiores llegan a unirse mientras, la tercera cauda (superior) tiende a caerse y el resto floral presenta una consistencia gelatinosa con pérdida total de su coloración.
d) Fase 4: la cápsula se encuentra en un máximo desarrollo tanto en longitud y diámetro, presentando un 25 % de resto floral descolorido algo transparente y casi seco; durante esta fase el fruto tiende a aumentar en grosor, sobre todo en la parte terminal del fruto en relación a la base.
e) Fase 5: la cápsula se encuentra inmadura con poco resto floral seco y el fruto presenta una coloración de verde oscuro intenso y empieza a aumentar tanto en grosor como en longitud.
f) Fase 6: en la parte superior del fruto se distingue un tejido abultado verde oscuro de manera prominente, sin resto floral y fruto tiene una coloración verde oscura y de consistencia dura.
g) Fase 7: la cápsula se encuentra en proceso de maduración donde se evidencia que las tres suturas longitudinales se vuelven cada vez más prominentes y el fruto tiende a tener una coloración de verde claro.
h) Fase 8: el fruto ya se encuentra maduro debido a que de manera natural las dos suturas de la parte inferior comienzan a abrirse y liberar las semillas.

El tiempo que toma en madurar los frutos M. solomonii, desde los indicios de su formación fue ocho meses (224 días). No obstante, en otra especie de Masdevallia caudata requiere 151 días para la formación de frutos hasta la liberación de las semillas (Ordoñez, 2013). En Masdevallia coccinea el tiempo de maduración de frutos es de seis meses (180 días) (Matallana-Puerto et al., 2022).
Porcentaje de frutos formados
De la población estudiada se contabilizó un total de 45 flores de las cuales, en 12 flores el ovario empezó a aumentar de volumen, con lo cual demuestra que hay indicios de fecundación, sin embargo, al cabo de ocho meses, tres de estos frutos alcanzaron a madurar, que representa un 13 %. En observaciones de campo, los frutos que no llegaron a formar comenzaron a podrirse, esto probablemente se deba a que el ambiente donde están los individuos corresponde a un bosque montano nublado (Kessler y Beck, 2001). M. solomonii que alcanzó un 13 % de producción de frutos, lo cual está por debajo del parámetro esperado de un 20 %. En un trabajo realizado por Ordoñez (2013) reporta un 26.31 % de producción de frutos por geitonogamia y xenogamia en Masdevallia caudata.
Diámetro y longitud de los frutos
A las ocho semanas de evaluación el diámetro y longitud de los frutos de M. solomonii alcanzaron 7.35 ± 0.65 mm y 19.8 ± 3.8 mm. Los frutos de las diferentes especies de orquídeas silvestres muestran cierta variabilidad en la forma, el tamaño y la disposición. Respecto al tamaño, esta difiere en cuanto a la especie, género de una misma taxa (Benito, 2017).
Descripción morfológica de las semillas durante el proceso de germinación in vitro de M. solomonii
Las semillas de M. solomonii durante la germinación y desarrollo de las plántulas pasaron por las siguientes fases (Figura 7):
Figura 7 Esquema de las fases morfológicas de las semillas de M. solomonii durante el proceso de germinación. Fase 0: semillas sin germinar. Fase 1: imbibición de agua por la semilla. Fase 2: aumento de volumen y presión interna del embrión a la testa. Fase 3: desarrollo del protocormo o semilla germinada. Fase 4: aparición del brote meristemático apical y emergencia de rizoides. Fase 5: surgimiento de las hojas diferenciadas y desarrollo del plantin.
Germinación
En la Figura 8 a las 18 semanas de siembra la mayoría de semillas permanecieron en fase 0 en las dos intensidades lumínicas, no obstante, a una baja intensidad lumínica (1.965 µMol m-2s-1) las semillas presentaron mayor porcentaje en fases de 3, 4 y 5 con 2.69, 11.58 y 8.11 % respectivamente, y bajo estas condiciones las semillas germinaron en un 22.39 %. Mientras que a una alta intensidad (7.992 µMol m-2s-1) germinaron un 8.16 %. Además, mencionar que la prueba T de Student mostró diferencias significativas en ambos tratamientos.
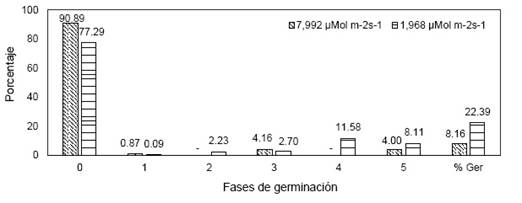
Figura 8 Fases de desarrollo de M. solomonii a diferentes intensidades lumínicas (7.992 µMol m-2s-1 y 1.965 µMol m-2s-1) después de 18 semanas de la siembra.
De hecho, la calidad o composición espectral de la luz ejerce diferentes efectos sobre los procesos fisiológicos en los que interviene, especialmente en la germinación. También afecta la morfogénesis y la organogénesis. Estos efectos no son producidos en igual medida por todos los tipos de luz (radiaciones de cualquier longitud de onda), ya que algunas radiaciones concretas tienen un efecto notable, mientras que otras tienen poco o ningún efecto (Mohr, 1995). En M. solomonii según los resultados obtenidos se puede corroborar que la intensidad lumínica juega un papel importante durante la germinación, al igual pasa en fases más avanzadas y llegan a formar vitroplantas. Este efecto posiblemente responde a que en la naturaleza las plantas de M. solomonii viven en las copas de los árboles o en dosel del bosque y está rodeado por la vegetación y esto influye en que la luz no llegue de manera directa. En un estudio realizado en Masdevallia auropurpurea Reich (Pedraza, 2011) en condiciones de oscuridad, los protocormos se desarrollaron con un mayor tamaño y diámetro que con iluminación.
Al respecto, algunos autores mencionan que las semillas de orquídeas tienen preferencia de germinación en la luz y otras en la oscuridad, entre estas podemos mencionar a Cypripediumrs, que germinan bien en la oscuridad y las plántulas jóvenes sobreviven mejor si se mantienen hasta 90 días en la oscuridad (Yamada, 1963 citado en Arditii, 1963), mientras que Cattleya no gemina en la oscuridad (Burgeff, 1909 citado en Arditii, 1963).
Comparando con otras especies del mismo género, M. solomonii tiene un bajo porcentaje de germinación, ya que en M. chaparensis y M. exquisita, reportaron un 42 y 53 % de germinación respectivamente (Villegas, 2003; Sánchez, 2007) y en M. caudata un 50 % de germinación (Ordoñez, 2013). Esta diferencia probablemente se deba a que los frutos colectados de su población natural pasaron dos meses en ambiente en el laboratorio previo a la siembra y posiblemente su viabilidad haya afectado en la germinación.
CONCLUSIONES
Los botones florales empezaron a aparecer en septiembre, y el mayor pico se registró en noviembre el pico de fructificación se presentó en mayo, estos están directamente relacionadas con la precipitación. de mayo a octubre se presentaron más del 40 % de la población en estado vegetativo (estéril). La producción de frutos fue un 13 % con un diámetro y longitud promedio 7.35 ± 0.65 mm y 19.8 ± 3.8 mm respectivamente y requiere ocho meses en madurar. A una baja intensidad lumínica (1.965 µMol m-2s-1) presentó mayor porcentaje de germinación (22.39 %), que con una alta intensidad (7.992 µMol m-2s-1) de 8.16 %.
Los resultados demuestran que, M. solomonii es una especie muy vulnerable por el escaso número de individuos y está restringida a factores intrínsecos (producción de frutos y germinación) y extrínsecos (condiciones ambientales y antropogénicos) y en base a esta información es referencial para la realización de programas de aprovechamiento.

