









 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
Permalink
INTRODUCCIÓN
Los pastos de la diversidad de praderas nativas de las zonas altas en Bolivia, constituyen una importante fuente de alimento para el ganado de altura. Entre las gramíneas destacan la cebadilla (Bromus catharticus) y cola de ratón (Hordeum muticum) como alimentos energéticos y entre las pocas leguminosas del altiplano, el trébol silvestre (Trifolium sp.) y la añahuaya (Adesmia spinosisima), por el alto contenido de proteína, Finalmente el q’awchi (Suaeda foliosa) por su adaptación a suelos salinos y alto contenido de proteína cruda.
La semilla es la unidad de reproducción sexual de las plantas y tiene la función de multiplicar y perpetuar la especie, siendo este el medio más eficaz para que se disperse en el tiempo y espacio (Doria, 2010). Las semillas de estos pastos nativos, presentan sustancias químicas o estructuras físicas duras, que les impide iniciar el proceso de germinación, además sus dimensiones físicas de tamaño y morfología son desconocidas
Mancipe-Murillo et al. (2018), señalan que la viabilidad denota el potencial que tiene una semilla para permanecer viva, la cual está ligada al éxito o fracaso reproductivo de las poblaciones y de esta manera es una de las primeras variables de calidad a evaluar en las cinco especies de semillas.
Existen varios métodos para determinar la viabilidad de semillas de especies domésticas (Moreno-Álvarez et al., 2001) tales como el test de electro conductividad, test del índigo carmin, el método de Cloruro Ferrico (FeCl3) (Barone et al., 2016), la técnica de los rayos X descrita por Sosa-Luría et al. (2012). Sin embargo, la prueba bioquímica del tetrazolio resulta ser más rápida y confiable (Hernandez et al., 2009). En la presente investigación se determinaron el efecto de tres tratamientos de soluciones de tetrazolio: 0.5, 0.1 y 1.0
%, en la viabilidad de las cinco especies de semillas nativas.
Especies forrajeras nativas, con gran potencial de utilización en el contexto pastoril, son poco o deficientemente utilizadas en función de la carencia de informaciones técnico-científicas sobre sus características físicas, morfología y calidad de las semillas que producen.
MATERIALES Y MÉTODOS
Ubicación de la zona de estudio
El estudio se realizó en el laboratorio de semillas de la Estación Experimental Patacamaya (Ex - IBTA), dependiente de la Facultad de Agronomía, Universidad Mayor de San Andrés (Bolivia). Geográficamente ubicado entre los paralelos 17° 15′ 43.9″ de Latitud Sur y 67° 56′ 45.5″ de Longitud Oeste, a una altitud de 3 785 a 3 899 m s.n.m., a 105 km del centro de la ciudad de La Paz (PDM Patacamaya, 2012).
Metodología
Especies de semillas nativas
Las especies nativas provenientes del banco de semillas de la Estación Experimental Patacamaya fueron; añahuaya (Adesmia spinosissima), cola de ratón (Hordeum muticum J.), q′auchi (Suaeda foliosa Moq.), cebadilla (Bromus catharticus) y, layu layu (Trifolium amabile).
Estudio morfológico
La descripción morfológica de la estructura externa e interna se realizó en 10 semillas de cada especie. Además de observar el color con las tablas de Munsell, se identificaron la forma y el embrión de cada semilla, con un microscopio estereoscopio Leica EZ4 HD zoom 1x, 2x, 3x, y una navaja para los cortes de los tejidos duros.
Tamaño y peso de las semillas
La determinación de estas variables se realizó en 10 semillas de cada especie, midiendo la longitud, diámetro y grosor expresados en milímetros, con un calibrador digital Kamasa (0-150mm/0-6). El largo de la semilla desde la parte inferior hasta la parte superior de la misma, el diámetro en la región media de la semilla y el grosor volteando la semilla lateralmente. El peso fue determinado en una balanza digital de precisión de 0.001 g, considerando una muestra homogeneizada, con diez repeticiones de 100 semillas (ISTA, 2016).
Análisis de laboratorio
Para determinar la viabilidad se pesaron 0.100, 0.500 y 1 g. de sal de tetrazolio, en una balanza digital de precisión de 0.001 g para la preparación de soluciones de tetrazolio al 0.1, 0.5 y 1 %, con un pH de 6 a 8, posteriormente fueron resguardadas en un ambiente oscuro y fresco por 24 horas.
Se seleccionaron 100 semillas de cada una de las cinco especies nativas con cuatro repeticiones, haciendo un total de 2 000 semillas. Las mismas fueron remojadas por separado en agua destilada durante 48 horas. Posteriormente, fue removido el pericarpio o endospermo de cada semilla con hojas bisturí. En las semillas con testa se las eliminó y luego se realizaron cortes longitudinales, con la finalidad de facilitar la penetración del tetrazolio al embrión (corte dorsal) (Mancipe-Murillo et al., 2018).
A continuación, se prosiguió con la inmersión de las semillas del q′auchi y layu layu en un volumen de 2 ml de cada solución de tetrazolio (0.1, 0.5 y 1 %) y las semillas de cebadilla, cola de ratón y añahuaya en 4 ml de cada solución. Posteriormente las cajas Petri conteniendo las semillas de cada especie y tratamiento, se colocaron en un ambiente oscuro y cerrado a una temperatura de 35 °C durante dos horas. Los resultados se observaron en un microscopio objetivo 40x, basándose en el número de semillas teñidas de color rojizo, debido a la reacción del tetrazolio con el tejido vivo de la semilla. Se consideró como semillas viables a aquellas que presentaron el embrión totalmente teñido. Como semillas no viables, a aquellas que no reaccionaron con el tetrazolio, tenían mínimas zonas teñidas a lo largo del embrión o bien aquellos donde el eje embrionario y cotiledones presentaban tejidos deteriorados rojos, semillas enteras sin teñir, cotiledones y embrión no teñido. El porcentaje de semillas viables fue calculado con la siguiente fórmula (FAO y África Seeds, 2019):
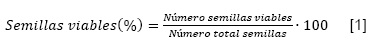
Análisis estadístico
Los datos de las variables viabilidad e inviabilidad de las semillas nativas fueron organizados en un arreglo bi-factorial, con factores fijos especie y tratamiento, analizados en un modelo lineal generalizado con el programa estadístico S.A.S. (versión 9.2. 2004), de acuerdo a la ecuación lineal aditiva (1). Los promedios fueron comparados mediante la prueba de Tukey.
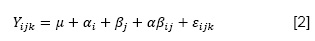
Dónde: Yijk = una observación cualquiera en la i-ésima especie, j-ésima semilla viable y en j-esima interacción; μ = promedio del experimento; α = efecto de i-ésima especie (i = 5: i1 = cebadilla; 2i = cola de ratón; 3i = añahuaya; 4i = layu layu y 5i = q’auchi); β = efecto de la j-ésima semilla viable (j =2: 1j= semillas viables y 2j= semillas no viables); αβij = efecto de la j-ésima interacción especie por viabilidad de las semillas; εijk = Error del experimento.
RESULTADOS Y DISCUSION
Características físicas
La calidad física de la semilla según ISTA (2016), involucra adicionalmente, tamaño y forma de la semilla, peso de mil semillas, color y daño por insectos y hongos. En la Tabla 1, se observa que la cebadilla y la cola de ratón obtuvieron las mayores dimensiones de largo y peso con 12.30 mm, 7.42 g por 1 000 semillas y 9.36 mm, 4.67 g por 1000 semillas, respectivamente. Sin embargo, en ancho y grosor fue la añahuaya la que destaco con 2.93 y 1.15 mm. El peso del q′auchi con 1.52 g fue el valor más bajo. Villca (2018) reporta un tamaño longitudinal de 0.08 a 0.14 mm, peso de 1 000 semillas de 0.75 g, valores menores al presente estudio. Para el Trifolium amabile varían con los pisos altitudinales y suelos, pero completamente diferentes a las halladas en esta investigación.
En trece genotipos de cebadilla, Abbott y Pistorale (2011) estimaron pesos de 1 000 semillas entre 9.94 a 10.43 g, ampliamente superiores a 2.49 g de nuestro estudio (Tabla 1). Estos valores coinciden con la variación tanto por efecto del material genético 8.78 a 11.58 g, como de la fecha de clausura entre 9.21 a 11.24 g en el peso de 1 000 semillas, reportados por Renata-Wolff et al., (2005). Lo mismo hallaron en tres especies y tres líneas de cebadilla criolla 4.27 a 10.75 g por 1 000 semillas (Covas y Ruiz, 2020). Los resultados de 1.2 a 1.7 mm de tamaño del q′auchi hallados por Pozo y Alzerreca (1990), son similares a nuestro trabajo.
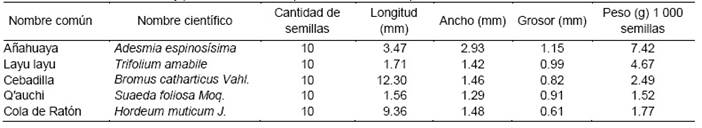
El peso de las semillas de la leguminosa Desmodium incanum DC, registrado de 4.1 g, (Gimena-Gillij et al., 2021) fue ligeramente inferior al peso de Medicago polymorpha L., de 4.5 g (Ovalle et al., 2005), y superior al de M. sativa (2.21-2.49 g), pero similar a 4.67g de la leguminosa nativa Trifolium amabile.
El éxito en el establecimiento de especies forrajeras introducidas como Lolium perenne dependerá en gran medida del tamaño de las semillas, sin embargo, depende de más factores. En Dactylis glomerata, el tamaño no sería un parámetro de apoyo para garantizar un establecimiento adecuado. Además, existen otros factores como humedad, cosecha, almacenamiento, condiciones climáticas, reserva de nutrientes que influyen en la calidad fisiológica de la semilla, en términos de germinación y emergencia (Mendoza-Pedroza et al., 2018). Los altos porcentajes de germinación y pureza hallados por Condori y Martínez (2020) y en el presente estudio, indican que la cebadilla y cola de ratón pueden ser cultivados en regiones similares al altiplano central.
Morfología
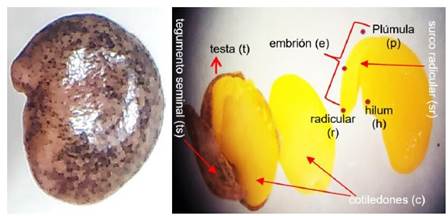
La semilla de añahuaya (Figura 1) visto de afuera es de color negro jaspeado, de forma elipsoidal redonda. La semilla interiormente tiene una forma alargada de color castaño claro, donde se puede observar el surco ventral (sv), el embrión (e) y el resto de la base radicular (r).
En la superficie externa se observa el tegumento seminal (ts, capa gruesa suberificada) que protege al embrión, y puede participar en la inhibicion de la germinación de las semillas por compuestos fenológicos (floridizim, ácido salicílico, ácido clorogénico y cumarina) (Bewley y Black, 1994) presentes en sus capas. Luego se ditingue una pequeña cicatriz llamada hilum (h), fisura que permite el intercambio de oxigeno, pero no de humedad en las semillas duras como la añahuaya. Al corte longitudinal en la semilla se puede observar el embrión, dos cotiledones (c) que darán origen a las hojas, la radícula a la raíz (r), la plúmula (p) al futuro tallo y el surco radicular (sr) (Figura 1).
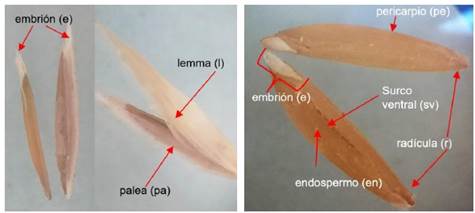
La semilla de cola de ratón es de color amarillo claro de forma ovoide alargado. La morfología de la semilla de cola de ratón (Figura 2) en su parte externa está compuesta de lemma (l), palea (pa) y una arista lateral color gris jaspeado. Se logra ver el surco ventral (sv), embrión (e) y el pericarpio (pe). Después de retirar los tegumentos el corte longitudinal mostro el endospermo (en), embrión (e), hilum (h), los pelos de pincel (pp). Mientras que, al extremo del embrión, con la lente de mayor aumento del microscopio, se observó, la radícula (r), la plúmula (p) y el micrópilo (m).
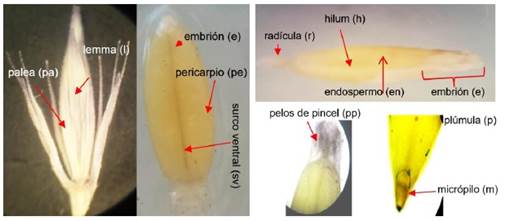
La semilla de cebadilla es de color castaño claro, de forma cariópside alargado. La Figura 3 muestra en la parte externa de la semilla: la palea (pa), lemma (l) y el pericarpio (pe). Al corte longitudinal de la semilla, se observan el embrión (e) con una lente de mayor aumento del microscopio, la radícula (r), surco ventral (sv) y endospermo (en), pero no se observa el escutelo, porque es el cotiledón único transformado en órgano absorbente adosado al endospermo (en). Según Perissé (2002), las estructuras responsables del mecanismo de la latencia frecuentemente no sólo incluyen a la testa, sino también otras partes de la semilla como, la palea (pa) y el lemma (l) en las gramíneas. Estas estructuras son muy delgadas en la cebadilla.
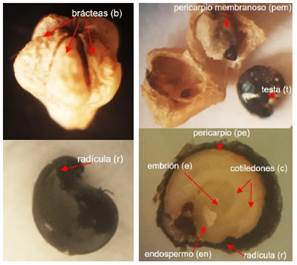
La semilla del q'awchi es de forma periforme achatada de color negro (Figura 4). Perissé (2002) menciona que el endospermo (en), pericarpio (pe) y la testa (t), imponen la latencia de varias maneras, ya sea interfiriendo la entrada de agua, el intercambio de gases o conteniendo inhibidores químicos que impiden el escape de los mismos presentes en el embrión, modifican la luz que llega al embrión o bien ejerciendo constricción mecánica, estructuras que en la semilla del q'awchi son muy notables.
En la parte externa de la semilla del q'awchi (Figura 4), es un fruto de forma de un aquenio de color gris con cinco carpelos y poroso, denominados brácteas por Pozo y Alzerreca (1990). La semilla verdadera está cubierta por un pericarpio membranoso casi transparente (pem), cuya superficie es lisa de color negro brilloso. Realizando un corte longitudinal se puede observar el embrión (e), endospermo (en) casi transparente, radícula (r), cotiledones (c) y el pericarpio (pe); además se observa la testa (t) o tegumento según Pozo y Alzerreca (1990).
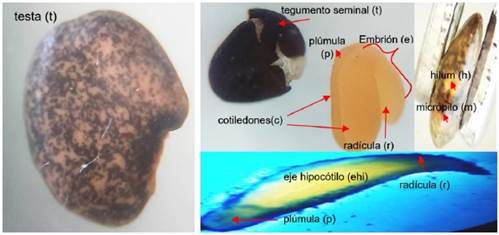
La semilla de layu layu es de color negro jaspeado, lisa de forma ovoide (Figura 5), lo que no coincide con la forma arriñonada, café oscuro que señala Villca (2018). En la parte externa presentan: el hilum (h) y el micrópilo (m), y una capa dura, el tegumento seminal o testa (t). En las leguminosas está cubierta protectora está formada por capas: la testa gruesa, suberificada, y una membrana transparente brillosa, que condicionan la dormición de la semilla, como consecuencia de que alguna o ambas son impermeables al agua y a los gases. En la parte interna se observan: el embrión (e) con dos cotiledones (c), un eje embrionario (e) definida, también podemos ver el eje hipocótilo (ehi) y la plúmula (p) que es una yerma opuesta al embrión (Figura 5).
Viabilidad e inviabilidad
En la Tabla 2, se muestra la influencia de la especie y el tratamiento de tetrazolio en la viabilidad e inviabilidad de semillas nativas. La especie y los tratamientos ensayados, al igual que su interacción, tuvieron un efecto significativo (p≤0.01) en la viabilidad e inviabilidad de las semillas nativas, es decir, estamos seguros con 99 % de probabilidad que existen diferencia entre especies nativas y tratamientos. El promedio general de semillas viables fue de 78.67 %, con un coeficiente de variación de 7.10 % y un rango amplio del 15 a 97 %. El porcentaje promedio de semillas no viables fue de 21.32 % con un desvío coeficiente de valor de 24.17 % en un rango de 3 a 85 % (Tabla 2).

Comparación de medias
En la Tabla 3, se muestra la comparación de promedios de viabilidad e inviabilidad de semillas nativas por la prueba de Tukey, según especies, tratamientos e interacción especie por tratamiento.
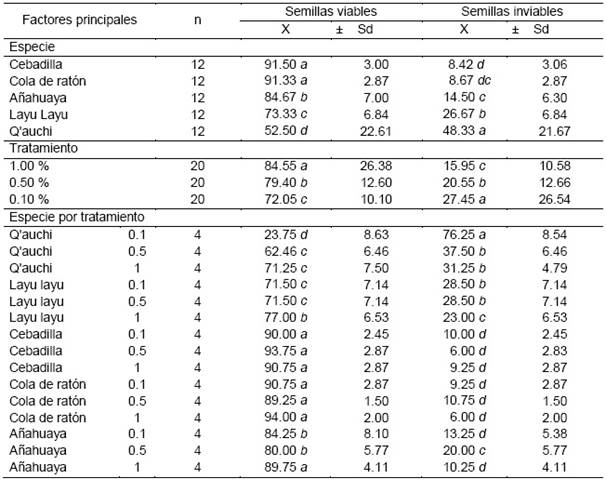
Especie
A la prueba de Tukey, estas diferencias son visibles únicamente entre tratamientos. La viabilidad de la cebadilla y cola de ratón fueron similares con 91.50 y 91.33 % respectivamente y superiores a la añahuaya (84.67 %), layu layu (73.33 %) y especialmente al q′auchi (52.50 %) (Figura 6).
Mancipe-Murillo et al. (2018), encontraron en semillas tropicales altoandinas diferencias en el valor de viabilidad, entre pruebas realizados sin ningún acondicionamiento de la semilla, con ruptura de la testa, mediante la prueba del embrión, y corte dorsal, similar a los valores altos obtenidos en tres semillas nativas: cebadilla, cola de ratón y añahuaya, con las tres concentraciones de tetrazolio del presente estudio.
Rojo et al. (2012) determinaron diferencias significativas (P<0.05) para la viabilidad entre tres especies de leguminosas, con valores promedios de 24.17 % para Pueraria phaseoloides (Roxb.) Benth, de 76.16 % para Lotus corniculatus L., y de 46.75 % para Gliricidia sepium Lam de Wit., estos valores fueron menores al de cebadilla, cola de ratón y añahuaya de la presente investigación. En cambio, las gramíneas limpias Brachiaria decumbens Stapf. y Panicun maximun Jacq. c.v. Tanzania, respondieron con mayores valores promedio de viabilidad; 78.5 y 64.0 % respectivamente, estadísticamente diferentes (P<0.05), con relación al layu layu y q′auchi, de nuestro estudio.
Hernández-Guzmán et al. (2021) en dos especies de pasto nativos del desierto Chihuahuense, banderita [Bouteloua curtipendula (Michx). Tor r.] y navajita [Bouteloua gracilis (Willd. Ex Kunth) Lag. ex Griffiths], así como de dos pastos introducidos de África a Estados Unidos y posteriormente a México: buffel [Pennisetum ciliare (L.) Link] variedad Común Americano y rhodes (Chloris gayana Kunth) variedad Bellm también obtuvieron diferencias significativas de: 70 a 90 %, 44 a 87 %, 44 a 94 % y 48 a 90 % en la viabilidad para semillas grandes y pequeñas respectivamente.
Por su parte Martin et al. (2016) para zámota (Coursetia glandulosa Gray) arbusto forrajero leguminosa sin espinas, reporta semilla pura viva (SPV) de 89.1 %, valor superior al layu layu, añahuaya y q′auchi, y 94.1 % para mezquite [Prosopis juliflora (SW) DC], árbol forrajero leguminosa con espinas, valor superior a todas las especies de semillas de nuestro trabajo. Ambas forrajeras tienen alto contenido de proteína, y son palatables para el ganado y fauna silvestre.
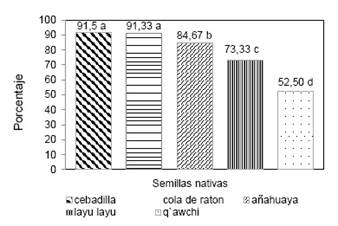
En relación a la inviabilidad de: layu layu, cola de ratón, cebadilla y añahuaya según la Figura 7, el q′auchi fue la especie con mayor valor promedio 48.33 % de semillas inviables, superior al promedio 26.67 % del layu layu, y al promedio 14.50 % de la añahuaya, y a las otras dos especies nativas con apenas 8.42 % para la cebadilla y 8.67 % para la cola de ratón.
Tratamientos
La viabilidad de las semillas nativas en estudio, fueron diferentes (p≤0.05) entre las concentraciones de tetrazolio ensayados. Siendo superior el tratamiento de la solución 1 % de tetrazolio con 84.55 % de semillas viables, respecto a 79.40 % de semillas viables obtenida con 0.5 % de concentración de tetrazolio y 72.05 % de semillas viables obtenidas con 0.1 %.
En semillas de Bombacopsis quinata, especie forestal valiosa para recuperación de bosques; Espitia et al. (2017), observaron el mayor porcentaje de semillas viables con 91 %, a la concentración de 0.5 % de tetrazolio y un tiempo de tinción de tres horas, pero sin diferencia a 84 % obtenida con la concentración de 1.5 % en igual tiempo de tinción, estos valores son superiores a las concentraciones de 0.1 y 0.5 %, del presente estudio, pero similares a 1 % de concentración de tetrazolio. Estos mismos autores; en semillas de Anacardium excelsum, obtuvieron mayores porcentajes de semillas viables en las concentraciones 0.5 y 1 % de tetrazolio, con 57 y 64 % de viabilidad respectivamente, siendo inferiores a los obtenidos en la presente investigación.
Barone et al. (2016), en semillas de Cedrela fissilis, hallaron porcentajes similares de viabilidad, a la concentración más baja de tetrazolio 0.1% del presente ensayo, pero superiores a las halladas en semillas de Myrocarpus frondosus.
Para la concentración de 1 %, la viabilidad máxima de 91 %, se alcanzó a las dos horas, con la concentración de 0.5 %, el máximo de 95 % de viabilidad se alcanzó a las cuatro horas, estos resultados coinciden con la metodología indicada para Festuca y Lolium (Leist, 1997). A las cuatro horas, no se encontraron diferencias significativas entre concentraciones de 0.5 y 1 %, la coloración era rojo carmín y los sectores teñidos eran similares para ambas. Resultados concuerdan con lo propuesto para Bromus spp. en ISTA (2016), sin embargo, presentan valores mayores a los nuestros.
A la prueba de Tukey (p≤0.05), con la menor concentración de tetrazolio 0.1 %, se detectó la mayor cantidad de semillas inviables (27.45 %). Entre tanto con la mayor concentración 1 % de tetrazolio, el porcentaje fue el menor (15.95 %), mientras que en la concentración intermedia de tetrazolio (0.5 %) la inviabilidad fue de 20.55 %.
En semillas de Bombacopsis quinata (Espitia et al., 2017) con una concentración de 1 % de tetrazolio hallaron valores de inviabilidad de 18 %, diferente estadísticamente al rango desde 0 a 5 % de semillas inviables, del resto de tratamientos (0.5 y 1.5 %). Estos mismos autores para semillas de Anacardium excelsum obtuvieron con una concentración de 0.5 % de tetrazolio, 72 % de semillas inviables, diferentes a 34 y 43 % de semillas inviables de los tratamientos de 1.5 y 1.0 %. En la presente investigación no se incluyeron tiempos de inmersión, por lo que los resultados se deben a la influencia de los niveles de las soluciones de tetrazolio experimentados.
Interacciones
A la prueba de Tukey entre los promedios de viabilidad de la cebadilla y cola de ratón (Tabla 3), resultado de la interacción especie por tratamiento, no existieron diferencias significativas (p≤0.05). Pero entre las especies: layu layu, q′auchi y añahuaya las diferencias (p≤0.05) fueron a favor de la mayor concentración (1 %). Lo mismo sucedió con los valores de inviabilidad, excepto que, para las últimas tres especies de semillas nombradas, las diferencias fueron a favor de la concentración más baja (0.1%).
La utilización de la solución de tetrazolio al 1 % en semillas de q’auchi permitió obtener mayor viabilidad (71.25 %), respecto al tratamiento con las soluciones de 0.5 % (62.46 %) y 0.1 % (23.75%) respectivamente. En tanto, en semillas de layu layu, la mayor viabilidad (77 %) se obtuvo con la concentración de 1 % de solución de tetrazolio.
Con respecto a las semillas de cebadilla, la viabilidad fue similar en las tres concentraciones de tetrazolio (0.1, 0.5 y 1 %). Las semillas de cola de ratón sometidas a las soluciones de tetrazolio con concentraciones de 0.1, 0.5 y 1% tuvieron similar viabilidad. Finalmente, las semillas de añahuaya obtuvieron mayor viabilidad en concentraciones de 1 % (89.75%), seguido de las concentraciones de 0.1 % (84.25 %) y 0.5 % (80 %) respectivamente.
El uso de soluciones de tetrazolio con mayor concentración (1 %) se convierte en opciones más adecuadas por permitir el mayor rendimiento de la sal de tetrazolio durante los análisis. Por otra parte, de acuerdo a la evaluación sobre la inviabilidad de semillas, en las especies q’auchi, layu layu y añahuaya presentaron mayor inviabilidad (76.25, 28.50 y 20 %) a una concentración de solución de 0.1 y 0.5 % de tetrazolio respectivamente.
CONCLUSIONES
Existen diferencias de tamaño y peso entre las cinco especies de semillas nativas, siendo las más grande y pesada la añahuaya; y la más pequeña y liviana la semilla de cola de ratón. Las semillas de añahuaya y layu layu poseen las mismas estructuras morfológicas, diferenciándose del q’awchi en su cubierta externa conformada por capas duras suberificadas llamadas brácteas.
La testa presente en las semillas de añahuaya, layu layu y q’awchi condicionan una dormancia química, además de una dormancia física y mecánica, además en el q’awchi por la dureza de la cubierta y el endospermo, cuyos tejidos podrían oponen resistencia mecánica al crecimiento del embrión.
La prueba de tetrazolio utilizando la concentración del 1 % permitió obtener mayor viabilidad en las semillas de q’auchi, layu layu y añahuaya, respecto al tratamiento con las soluciones de 0.5 y 0.1 %, respectivamente. Con respecto a las semillas de cebadilla y cola de ratón, las viabilidades fueron similares en las tres concentraciones de tetrazolio (0.1, 0.5 y 1 %). Las soluciones de tetrazolio más diluidas presentaron mayor inviabilidad de semillas, probablemente a la poca tinción de las semillas.