










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de Investigación e Innovación Agropecuaria y de Recursos Naturales
versión impresa ISSN 2409-1618
RIIARn vol.5 no.Especial La Paz 2018
ARTÍCULOS
Determinación de la edad óptima de faeneo, calidad y características productivas de la carne de llama
Condori G.16; Ayala C.17; Renieri C.18; Gerken M.19; Antonini M.20; Quispe J.16
16 Investigador, Proyecto DECAMA, Facultad de Agronomía, Universidad Mayor de San Andrés, Bolivia.
17 Docente Investigador, Facultad de Agronomía, Universidad Mayor de San Andrés, Bolivia.
18 Docente, Universidad de Camerino, Italia.
19 Docente, Universidad de Gottingüen, Alemania.
20 Investigador, ENEA, Italia.
Introducción
La producción y la comercialización de carne de llama en Bolivia, tiene características peculiares dentro su propia cadena productiva, desde luego existen aún algunas restricciones sobre todo por el desconocimiento de las bondades que tiene esta carne. De forma general y desde el punto de vista de higiene y las formas de comercialización, es evidente que no hubo grandes cambios y la oferta de la carne se mantiene como un producto tradicional, pese a los esfuerzos realizados por instituciones de investigación y de cooperación, para mejorar la calidad de la oferta de carne de llamas y alpacas, lo cual se logró medianamente mejorar la oferta de los subproductos transformados como el charqui y con ello consecuente mejorar de los ingresos de los productores.
En el marco del Proyecto SUPREME - Facultad de Agronomía de la Universidad Mayor de San Andrés, se han realizado investigaciones que ayudan a determinar la edad óptima de faeneo en llamas, en el cual se ha considerado un carácter principal como el peso vivo del animal, la cual varía de acuerdo con la edad del animal, así como también la composición bromatológica de la carne que cambia según la edad del animal, y el desarrollo alométrico de las partes de mayor importancia económica y es que a través de estos parámetros que se llegó a determinar un rango de edad óptima para el faenamiento de las llamas. En principio se considera un estudio de las características de la carne en llamas machos no castrados y castrados, la composición química de la carne y sus características físico-químicas.
Metodología
El trabajo se realiza en la Estación Experimental de Choquenaira, dependiente de la Facultad de Agronomía -UMSA, ubicada a 35 Km. de la ciudad de La Paz, a una altura de 3750 m.s.n.m., con una precipitación anual entre 400 a 600 mm concentrada en tres meses del año, una temperatura promedio de 14°C, la temperatura mínima registrada en el mes de junio alcanza a -3°C y la máxima durante los meses de enero y febrero alcanza a 20 °C. Geográficamente se encuentra situada a 16° 40' de latitud sur y 68° 10' longitud oeste.
Para el estudio se utilizan 50 llamas machos de una misma edad, nacidos entre la tercera semana de enero y la primera semana de febrero de 1997, los animales pertenecen al tipo "intermedio" y fueron colectados en la región de Turco del Departamento de Oruro. Los animales desde su arribo a la Estación Experimental de Choquenaira, se mantienen por las noches en un corral dormidero y la alimentación diaria se realiza bajo el sistema de pastoreo común en praderas nativas del tipo pajonal con predominancia de Stipa ichu y Festuca dolicophylla El destete se realizó a los 11 meses de edad y a los 12 meses se efectúo el castrado del 42% de los animales; posteriormente se formaron 7 grupos de 7 animales (4 enteros y 3 castrados). La matanza de animales de cada subgrupo se efectúa cada 3 meses a partir de los 13 meses de edad.
El control de peso corporal se efectúa con una balanza de 100 kg de capacidad y precisión de 250 g. Las medidas biométricas se realizan de acuerdo a la metodología empleada para ovinos por ASPA (Asociación Científica de Producción Animal, 1991), y son: a) Altura a la cruz.- se mide con el bastón de Lydken en un plano horizontal al punto más alto del tronco, b) Longitud de tronco.- se mide desde la articulación escapo-humeral a la tuberosidad del isquion, en forma diagonal, c) Ancho de Ancas.- se mide con el bastón de Lydken entre las articulaciones coxo-femorales y d) Perímetro torácico.- se mide con una cinta métrica alrededor del tórax detrás de la espalda al mismo nivel de la altura a la cruz. Tanto el control de peso como las medidas biométricas se realizan cada 31 días.
Antes del sacrificio se realizaron los siguientes controles de peso: 1) Control de peso corporal antes de someter a ayuno alimentario no hídrico, 2) Control de peso corporal después del ayuno de 24 horas. El sacrificio se realizó de acuerdo a la metodología seguida para bovinos y comprende los siguientes pasos: a) Aturdimiento, b) Colgado, c) Degüello, d) Desangrado, e) Desolle y f) Eviscerado.
Posterior al sacrificio se efectuó el control de peso de la carcasa en caliente y 24 horas. después el peso de carcasa en frío, en ambos casos a temperatura ambiente. A continuación se realiza el pesaje de la cabeza (incluyendo la lengua), piel más fibra y, patas; aparato digestivo, que comprende: esófago, estómago, intestinos delgado y grueso, con el contenido alimenticio; posteriormente el contenido alimenticio es vaciado y pesado para determinar el peso vivo neto (PVN); finalmente, se pesaron los órganos como el corazón, hígado y pulmón. Todos los pesos se efectúan en una balanza de 10 kg de capacidad y 5 g de precisión.
La carne está constituida por las fibrillas musculares que se reconoce por bandas, según Mora et al. (1994) indican que la banda o línea Z, es la unión de una sarcómera con otra, dentro del límite de la sarcómera se encuentra la banda A que constituyen los filamentos gruesos y delgados sobrepuestos entre sí. La banda H que forman los filamentos gruesos libres de filamentos delgados y la banda I formada solo por filamentos delgados. Un filamento grueso se constituye por muchas moléculas de miosina y tiene la forma de un bastón. El filamento delgado está formado por moléculas de actina los cuales tienen forma esférica y se unen como un collar. Charley (1995) describe la estructura anatómica del musculo como sigue:

Las enzimas necesitan convertir el glicógeno en ácido láctico y otras substancias que están en solución en el sarcoplasma. Esto incluye al musculo pigmentado mioglobina que es distinta a la hemoglobina en la sangre, donde aparece un término corto que se denomina almacenamiento de oxígeno en el musculo.
Determinación de características físicas de la carne
Inmediatamente después del sacrificio se procede a medir el pH con un equipo provisto de electrodo de superficie para carne, el cual se inserta a una profundidad aproximada de 2 cm en forma perpendicular con respecto a la superficie del músculo y a temperatura ambiente, se considera como dato el promedio de dos mediciones efectuadas casi en un mismo lugar del músculo, siguiendo la metodología recomendada por ASPA (1996), estas mediciones se realizan a 1, 6, 12, 24, 48 y 72 horas después de la muerte del animal.
Simultáneamente a la medición del pH, se determina el tiempo de duración de la rigidez cadavérica por el método físico, que consiste en comprobar la rigidez ejerciendo una presión sobre los músculos de las carcasas colgadas, tomando con la mano la articulación del carpo y tratando de flexionar los músculos del brazo y de la espalda, de forma tal que cuando la rigidez cadavérica esta instaurada no es posible flexionar el músculo (Aldana, 1984).
Las enzimas necesitan convertir el glicógeno en ácido láctico y otras substancias que están en solución en el sarcoplasma. Esto incluye al musculo pigmentado mioglobina que es distinta a la hemoglobina en la sangre, donde aparece un término corto que se denomina almacenamiento de oxígeno en el musculo.
Determinación de características físicas de la carne
Inmediatamente después del sacrificio se procede a medir el pH con un equipo provisto de electrodo de superficie para carne, el cual se inserta a una profundidad aproximada de 2 cm en forma perpendicular con respecto a la superficie del músculo y a temperatura ambiente, se considera como dato el promedio de dos mediciones efectuadas casi en un mismo lugar del músculo, siguiendo la metodología recomendada por ASPA (1996), estas mediciones se realizan a 1, 6, 12, 24, 48 y 72 horas después de la muerte del animal.
Simultáneamente a la medición del pH, se determina el tiempo de duración de la rigidez cadavérica por el método físico, que consiste en comprobar la rigidez ejerciendo una presión sobre los músculos de las carcasas colgadas, tomando con la mano la articulación del carpo y tratando de flexionar los músculos del brazo y de la espalda, de forma tal que cuando la rigidez cadavérica esta instaurada no es posible flexionar el músculo (Aldana, 1984).
La capacidad de retención de agua se determina por el método de compresión, el cual consiste en extraer del músculo largo dorsal una muestra de carne de 300 mg que es colocada entre una superficie plana de vidrio plexiglass y otra superficie cubierta con papel filtro S&S 2040b y posteriormente, comprimido en una prensa a una presión de 50 Kg/cm2. El dato a obtener es el área marcada en el papel filtro por la salida del agua y, el área de expansión de la carne producida por la fuerza de presión, Grau y Hamm (1953) modificado. La capacidad de retención hídrica se determina a 1, 6, 12, 24, 48 y 72 hrs. a temperatura ambiente.
El análisis bromatológico se realizó en el Laboratorio de Diagnóstico y Salud (SELADIS), en muestras de aproximadamente 350 g extraídas del músculo largo dorsal de cada carcasa entre la 8va y la 9na vértebra torácica. En el análisis se determina la proporción de: a) Proteína (Método de Kjeldahl), b) Humedad (método de desecación en estufa), c) Grasa (método de Barshall), d) Contenido de ceniza (método de Calcinación), e) Colesterol (método de Lieberman - Bouchardat).
Resultados
Aptitud de la llama para producción de carne
Es evidente que de las dos especies, llamas y alpacas distribuidas ampliamente en toda la región andina, contribuyen a la seguridad alimentaria de las familias de los Altos Andes, donde la llama ofrece una vocación productiva hacia la producción de carne y no solo por el mayor volumen corporal, sino porque el otro subproducto como es la fibra, para su aprovechamiento requiere del proceso adicional de descerdado, lo cual la hace menos eficiente en comparación con la alpaca, aunque actualmente se realiza el aprovechamiento de la fibra de llama para obtener el hilado artesanal.
Bajo el contexto anterior, podemos iniciar, que Fernández (1977) citado por Rodríguez (1983), señala que el peso al nacer en llamas es superior al de alpacas en 1.69 Kg, y el peso corporal anual de las llamas es superior en 19.1 kg al de alpacas y dentro la especie según Tichit (1991) la Q'ara es utilizada para la producción de carne y transporte y la T'amphulli para la producción de fibra, aunque se indica que de las dos especies domésticas se obtiene como producto o subproducto la carne.
Las llamas tienen una gran cualidad para la producción de carne pues convierten el forraje tosco de las alturas en un producto de alta calidad nutritiva, en comparación a las especies introducidas que no tienen ninguna posibilidades de producir en ecosistemas adversos donde si la llama o la alpaca quienes lejos de subsistir pueden producir.
Rendimiento de la canal
El rendimiento de la canal, es la relación entre el peso de la canal con el peso vivo del animal, la calidad de la canal depende de la composición de grasa, hueso y carne magra por tanto esta relación tiene una directa dependencia con la raza, es así que en bovinos especializados para la producción de carne el rendimiento a la canal será superior al de una raza no productora de carne. Por otro lado la robustez del animal influye de modo decisivo sobre el rendimiento y volumen de carne aprovechable (Pinto, 1975). El porcentaje de carne magra de la carcasa varía mucho y es inversamente proporcional al contenido de grasa.
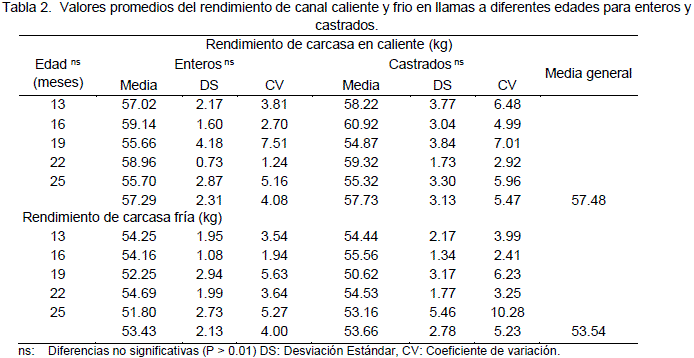
El rendimiento de la canal en llamas según Pinto (1975) es de 52.12% en carcasa caliente y 46.64%, después de 24 horas. Martínez y Alzerreca (1983) determinaron que para alpacas un rendimiento de 48.74% en carcasa caliente y en frío 46.36%, Bravo et al. (1981) reportan rendimientos de 58.1 y 59.6%, para llamas machos sin castrar y castrados respectivamente; a 1 año de edad. Al respecto Condori (2000) reporta valores promedios de 57.48 y 53.54 % en canal caliente y en frío (Tabla 2) respectivamente superiores a los reportados por Pinto (1975) y similar al reporte para carcasa caliente de Bravo (1981).
La merma que existe después de 24 horas (3.94%) es inferior al reportado por Pinto (1975) 5.48%, la merma es debido principalmente a la perdida de agua por exudación de la carne la cual está directamente relacionada a la capacidad de retención de agua.
Relación de parámetros de productividad y calidad de carne que determinan la edad optima faeneo en llamas
Considerando estos tres aspectos de importancia, como el balance de peso vivo y contenido proteico en la carne, y la evaluación alométrica de cortes comerciales, se sugiere una edad óptima de faeneo a partir de los 18 meses de edad, sin embargo considerando el acabado o engorde de los animales, que según González (1996) señala un tiempo que varía desde los 45 a 60 días obteniéndose un incremento de 10 kg de peso como promedio en alpacas y llamas y según Di Marco (1993) señala que para la terminación o acabado de bovinos es necesario cierta acumulación de grasa subcutánea, para evitar la deshidratación de la carcasa durante la maduración obteniendo una buena composición corporal para su comercialización. Por tanto se estima un rango de edad óptimo para la faena que tiene su inicio a partir de los 20 hasta los 25 meses.
Este rango de edad considera el balance óptimo de proteína y peso vivo y las consideraciones alométricas, por otro lado es pertinente señalar que según un estudio realizado por Ayala (1990) animales mayores de 3 años presentan en la carcasa macro quistes de sarcocystis con un alto índice de infestación que conlleva a una baja calidad de la carcasa. Este estudio coincide con Rivera etal. (1996) citado porCondori (2000), quienes indican que la carne de llama es ideal cuando el animal no pasa de los tres años de edad. Por que presenta mayor suavidad y jugosidad. Al respecto CISA (1996), señalan que las carcasas de llamas y alpacas machos y hembras de 12 a 24 meses son comercializadas como carne de primera y de buena calidad. Al respecto Rodríguez (1995) indica que las llamas alcanzan pesos substanciales solo hasta los dos años de edad.
Considerando el crecimiento diferencial de cortes comerciales Condori (2000) encontró que los cortes de pierna y brazuelo tienen un desarrollo relativo más precoz con relación a otros cortes, en animales comprendidos entre 19 y 25 meses de edad. Se establece que desde el punto de vista comercial la pierna y el brazuelo son cortes de mayor importancia por la mayor masa muscular que presentan. Significa que estos cortes a medida que pasa la edad del animal ya no aumentan substancialmente de peso, en cambio el cuello al contrario de la pierna y brazuelo aun continua en crecimiento en el periodo de desarrollo de 19 a 25 meses de edad. Esto demuestra que si el animal continua con vida aumentará en peso aquellos cortes de menor calidad como el cuello. Lo cual insinúa que una llama macho no deberá sobrepasar los 25 meses para ser beneficiado.
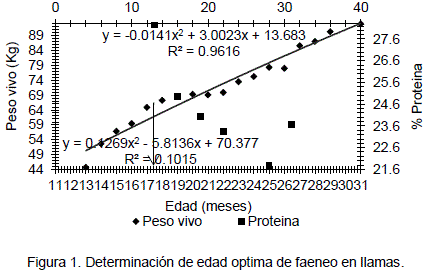
En la Figura 1, se presenta la relación inversa entre la evolución del peso vivo (kg) y la curva de decrecimiento del contenido proteico en función de la edad (meses) en machos castrados. Se observa que ambas curvas ajustadas se interceptan a una edad de 18 meses con 64 kg y 24.2% de proteína.
Características físicas que determinan la calidad de la carne Color
El color de la carne es un factor importante que está íntimamente relacionada con la mioglobina. La cual constituye las tres cuartas partes del pigmento total de la carne roja. La molécula de la mioglobina está formada por una proteína, la globina y un grupo hem que incluye un anillo planar de protoporfirina con un átomo de hierro. El contenido de mioglobina en los músculos, varía según las especies, la raza, el sexo y la edad. La función de la mioglobina en el animal vivo es almacenar oxígeno y por lo tanto los niveles son más altos en los músculos con mayor carga de trabajo. Estos músculos presentan una coloración más oscura como el caso de los músculos de la pierna que contienen más mioglobina (Varnam y Sutherland 1995 citado por Condori, 2000). A propósito según Charley (1995) un animal muere y el aporte de oxigeno cesa y los tejidos no se oxigenan. La mioglobina en la forma no oxigenada junto al hierro en su estado ferroso le proporciona a la carne recién cortada un color rojo púrpura. Sin embargo, cuando se expone al aire el pigmento de la superficie de la carne recién cortada se oxigena y forma la oximioglobina que tiene un color rojo cereza brillante. Cuando es expuesto a bajos niveles de oxígeno el complejo oxigeno - mioglobina se disocia y el hierro se oxida al estado férrico dando lugar a la metamioglobina de color rojo café.
Por otro lado el color de la carne, también está influenciada por las condiciones en que se ha sacrificado al animal. Condiciones inadecuadas pueden afectar el cambio del pH del músculo del animal después de la muerte. Este cambio está relacionada con el color de la carne puesto que un pH anormal se presenta en carnes tipo exudativa blanda y pálida (PSE) y carne seca, firme y oscura (DFD) (Varnam y Sutherland 1995 y Charley 1995 citados por Condori, 2000)
pH en la carne
Los organismos animales necesitan de energía continuamente y la mayoría exceden en el consumo de sus requerimientos, este exceso es almacenado como reserva en forma de glucógeno principalmente en el hígado pero también se puede almacenar en los músculos, el cual tiene una estrecha relación con el descenso del pH después de la muerte del animal. El almacenamiento del glucógeno es temporal porque en el ayuno de 24 horas bajan los niveles de azúcar, en la sangre casi a cero a pesar de la reserva de glucógeno (Maynard et al. 1981 citados por Condori 2000). Al respecto Limonta (1987), señala que el pH de la carne es de 7.3 a 7.5 después del sacrificio esta tiende a bajar dependiendo de la cantidad de ácido láctico producido durante la glucólisis del glucógeno. El ácido láctico producido está en función de la cantidad de glucógeno presente en el animal al momento del sacrificio.
Cuando una animal muere se inicia los procesos metabólicos en el músculo que altera su naturaleza de cuando era vivo. La circulación cesa y los músculos no obtienen energía ya que la actividad mitocondrial cesa ante la ausencia de oxigeno interno. Consecuentemente el glucógeno, se convierte en ácido láctico en anaerobiosis por glucólisis post mortem. El descenso del pH puede estar relacionado con la producción de lactato, aunque los iones hidrogeno generados provienen de la hidrólisis del ATP y no de la producción de lactato (Varnam y Sutherland 1995 citados porCondori, 2000).
En las carnes que presentan condición PSE (pálidas suaves y exudativas) según Mora et al. (1994) el pH baja bruscamente después del sacrificio, debido a que el animal no tuvo las condiciones adecuadas de matanza. Cuando existe algún tipo de manipuleo excesivo antes del sacrificio, el animal libera la hormona adrenalina que acelera el desdoblamiento del ATP, por ello ocurre una rápida glucólisis, produciendo mucho ácido láctico que causa el anormal descenso del pH. En las carnes DFD (oscuras firmes y secas) el glucógeno ha sido consumido totalmente lo que ocasiona un pH final por encima de lo normal.
Aldana (1984) indica que el proceso de acidificación se debe a la transformación enzimática del glucógeno muscular en ácido láctico. El proceso de transformación continua incluso a temperaturas comprendidas entre los 0 °C y a los 6 °C aunque sufriendo una cierta atenuación a temperaturas inferiores a 0 °C. El pH, desde el punto de vista higiénico y sanitario, tiene importancia porque si las carnes que tienen pH final, neutro o alcalino es un estado favorable para el desarrollo y multiplicación de los microorganismos sobre todo los de putrefacción, por otro lado constituyen un peligro para el consumo y perjuicios económicos para la industria cárnica.
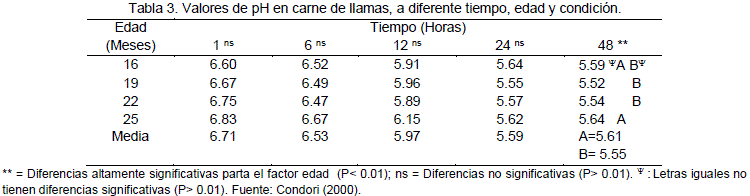
En la carne de camélidos, Condori (2000), demostró que la covariable temperatura y el peso inicial no influyen en el pH de la carne. La condición de castrado no influyen en el pH final de la carne, sin embargo existen diferencias altamente significativas (P<0.01) para el factor edad a las 48 horas después de la muerte del animal.
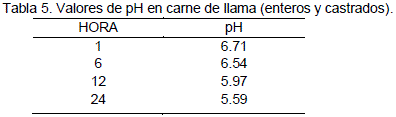
En la Tabla 3 se presentan los valores promedios de pH a diferentes tiempos después de la muerte. Los valores promedio obtenidos son: a 1 hora el PH = 6.71 y a las 24 horas pH = 5.59, valores similares a los reportados por Penny (1984) y Visier (1980), que coinciden en establecer un valor de pH = 5.5 a 24 horas para carnes rojas en general.
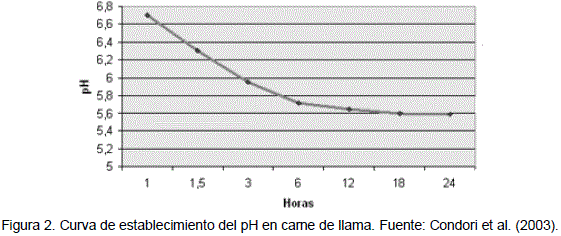
De acuerdo a la Figura 2, los valores de pH obtenidos hasta las 24 horas, según Condori et al. (2003), son similares a la tendencia de la curva de pH normal, presentada por Mora et al. (1994). Con una leve diferencia a las 6 horas el pH de la carne de llama no presenta este descenso pronunciado. La caída del pH de 6.71 a 5.61 y después a 5.55 en 48 horas es una tendencia corroborada por Limonta (1987). Quien indica que los valores de pH después del faeneo varían dependiendo de la cantidad de ácido láctico producido durante la glucólisis el cual baja desde pH = 7,5 a 5,7. Al respecto Aldana (1984 citado por Condori 2000) reporta valores de pH = 5.8 a 5.6 al cabo de 48 horas para carne de bovinos.
El efecto no significativo (P>0.01) del factor condición (castrado y sin castrar), para un pH final discrepa con Varnam y Sutherland (1995) citados por Condori (2000), quienes señalan que el pH final en vacunos es más alto en machos enteros que en los castrados.
Tomando en cuenta que las medidas se han efectuado en el músculo longissimus dorsi. Con estos resultados se establece que la carne de llama proveniente de una matanza adecuada presenta una evolución de pH normal que desde el punto de vista higiénico y sanitario, tienen una importancia económica ya que se encuentran dentro los rangos requeridos de un pH final bajo (5.55) hasta las 48 horas. Puesto que valores de pH cercanos a la neutralidad (pH = 7) o alcalinos (pH > 7), favorecen la reproducción de microorganismos entre ellos los de la putrefacción (Aldana, 1984).
En la Tabla 4 se presenta el análisis de varianza de las mediciones de pH efectuados en diferentes tiempos después del faenamiento del animal. El análisis estadístico confirma que existe diferencias altamente significativas para el factor edad a las 48 horas y diferencias significativas para la interacción edad por condición.
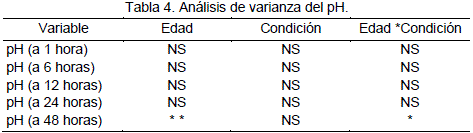
En la Tabla 5 se presentan los valores promedios de pH a diferentes tiempos después de la muerte del animal, el valor promedio obtenido a 1 hora es 6.71 y a 24 horas 5.59, este valor es corroborado por Penny (1984) que establece un valor de 5.5 a 24 horas.
Los valores de pH obtenidos hasta las 24 horas son consistentes con la curva de pH normal descrito por Mora et al (1994). De las pruebas de F y de la observación de la Tabla 6 se deduce que hay diferencias altamente significativas entre los efectos de las edades en la condición del animal, el efecto de la edad a5 y a3 son similares en la condición b1 y b2 de la misma forma hay diferencias en su comportamiento de la condición en cada una de las edades.

El efecto de las edades a5 en animales enteros hace que presente un pH alto por lo tanto mayor probabilidad a la descomposición en cambio en condición castrado a la misma edad el valor de pH es menor contrariamente en la edad a2 el entero presenta menor valor de pH y el castrado es más susceptible a la descomposición. A la edad a4 los animales presentan valores similares tanto en enteros como en castrados, a la edad a3 en condición castrado se presenta el valor más bajo de pH.
De la inspección de la Figura 2 se puede inferir que los castrados y enteros a una edad a2 presentan un valor de pH diferente y el castrado presenta el valor más alto, por lo tanto son carnes más susceptibles al desarrollo de microorganismos. Los enteros y castrados de 22 meses son los que presentan los valores más bajos de pH.
Capacidad de retención de agua
Una de las propiedades del músculo para retener agua tiene importancia en la apariencia de la carne antes de la cocción y su comportamiento durante la misma. Este aspecto da la impresión de jugosidad y de suculencia en el curso de la masticación de la carne. La sensación corresponde a la mayor o menor cantidad de líquido liberado por la carne pero además la impresión de suculencia está ligada a la "calidad del jugo exprimido". La capacidad de retención de agua después de la muerte del animal, generalmente disminuye a través de exudaciones (Visier, 1980).
Visier (1980) menciona que la capacidad de retención de agua evoluciona después de la muerte del animal, generalmente disminuye a través de exudaciones. Para Mora et al. (1994) la capacidad de retención agua depende de la cantidad de ATP que contiene la carne. Cuando la carne se encuentra en etapa pre-rigor, existe abundante ATP que mantiene separados la actina y la miosina creando un espacio libre entre el filamento grueso y delgado, en este espacio se almacena agua. Cuando alcanza la rigidez cadavérica, el pH declina y el ATP se consume gradualmente, se combina la miosina con la actina y forman el complejo actomiosina. Consecuentemente los músculos se contraen y se tornan firmes, rígidos y los espacios entre los filamentos gruesos y delgados se acortan y así se reduce la capacidad de retención hídrica.
La capacidad de retención hídrica es mínima en el punto isoeléctrico (PI) de las proteínas; el PI es el pH en el cual todos los grupos de las cadenas laterales están cargadas eléctricamente. En el pH el número de cargas positivas es igual al número de cargas negativas y se sitúa en un rango de pH = 5.0 y 5.4 coincidiendo con la rigidez cadavérica establecida (Mora et al., 1994).
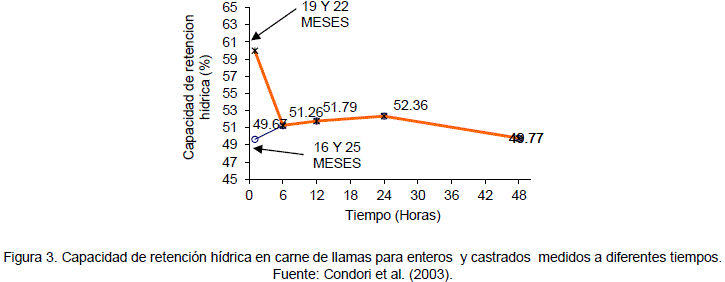
Para la carne de llama la capacidad de retención hídrica, presenta diferentes valores de acuerdo al momento de la medición así en la Figura 3 se observa que la mayor capacidad de retención de agua para carne de llamas enteros y castrados de 19 y 22 meses de edad se presenta en al cabo de una hora con un valor promedio de 59.99% de retención de agua que posteriormente desciende hasta un valor promedio de 51.26% a las 6 horas y un pH de 6.53 (Condori 2000). El valor de la capacidad de retención hídrica para el periodo señalado, para carnes rojas en general, se encuentra alrededor del 51%.
En llamas de 16 y 25 meses de edad, el valor promedio es 49.67% a una hora, después este asciende hasta las seis horas presentando un valor de 51.26%. A partir de las 6 horas no existen diferencias para el factor edad, presentando un leve descenso a las 48 horas (Condori et al2003). La diferencia de la capacidad de retención hídrica a diferentes edades se atribuye al desigual contenido ATP antes del sacrificio. Se señala esto, porque esta depende de la cantidad de ATP que queda en la carne, porque el ATP tiene la acción de separar la miosina de la actina produciendo un espacio libre donde se retiene el agua (Mora et al., 1994).
Capacidad de Retención hídrica
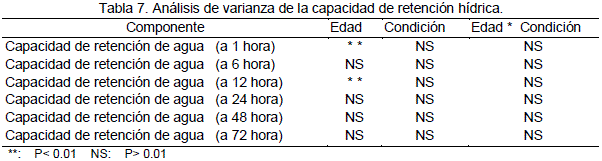
En la Tabla 7 se presenta el análisis de varianza de la variable capacidad de retención hídrica.
En la Tabla 8 se presentan los valores promedios del porcentaje de la capacidad de retención hídrica de muestras extraídas del músculo largo dorsal de la carcasa de llama.
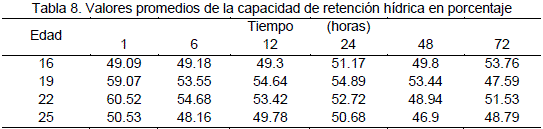
De la prueba de comparación de medias para el factor edad con respecto a la capacidad de retención hídrica efectuados a un tiempo de 1 hora después de la muerte del animal, se establece que la mayor capacidad de retención hídrica se presenta en animales de 19 y 22 meses de edad en la prueba efectuada a 12 horas después del sacrificio se determinó que la mayor capacidad de retención hídrica se presenta en animales de 19 meses de edad sin distinción de la condición.
Al examen de la Figura 3 se observa que la mayor capacidad de retención de agua en animales de 19 y 22 meses de edad se presenta a los momentos que siguen al sacrificio determinándose al cabo de una hora un valor promedio de 59.99% de retención de agua que posteriormente desciende hasta 51.26% a las 6 horas este resultado es consistente con lo señalado por Visier (1980) para carnes rojas en general.
Los animales de 16 y 25 meses de edad presentaron el valor promedio para una capacidad de retención de agua de 49.67% a una hora el cual asciende hasta las seis horas presentando un valor de 51.26%. A partir de las 24 horas no existen diferencias para el factor edad, presentando un leve descenso hasta las 48 horas, posteriormente tiende a permanecer constante.
En la Figura 4, se observa que al examen físico de la carcasa este alcanza la rigidez cadavérica en un periodo de 16 a 20 horas momento en el cual el pH tiene un valor de 5.8, y culmina cuando las articulaciones vuelven a ser flexibles a la presión que ocurre de 36 a 48 horas a una temperatura promedio ambiental de 15.29°C, la presión ocasiona la rotura del complejo actomiosina de acuerdo a Varnam (1995) este periodo es un estado en el que la maduración de la carne ha concluido.
Rigidez cadavérica
La rigidez cadavérica se define como la muerte de los músculos, que se presenta inmediatamente después del sacrificio, un estado de flacidez elástica y relajada, posteriormente se tornan duras e inextensibles. Transcurrida algunas horas de su iniciación, la rigidez desaparece, entonces los músculos se tornan nuevamente flácidos y deformables (Aldana 1984 citado por Condori et al., 2003), cuya velocidad de aparición según Visier (1980) está influenciada por la caída de pH, destrucción del ATP, contenido de glucógeno y la temperatura de la carne. Cuando la rigidez esta instaurada en los músculos, comienza la verdadera maduración de la carne. Según Varnam y Sutherland (1995), una consecuencia del cese de la glucolisis después de la muerte y del descenso de los niveles de ATP y ADP, es la rigidez cadavérica y cuya velocidad de la glucólisis está afectada por numerosos factores como el genotipo, la edad, especie, tipo de músculo y otros factores externos como la aplicación de drogas y temperatura.
La rigidez no se manifiesta en todos los músculos al mismo tiempo si no que transcurre por etapas y en un orden. Comienza por los músculos del cuello, tronco, extremidades anteriores y finalmente en las extremidades posteriores. Se comprueba la rigidez cadavérica tratando de flexionar los músculos del brazo y la espalda, en la canal. Cuando la rigidez cadavérica esta instaurada esta operación no es posible realizar (Aldana, 1984 citado por Condori et al., 2003).
La carne caliente en general presenta un pH = 6,2 lo que también favorece la cohesión del agua, y grasas además que existe una concentración limitada de ATP, esto en vacunos sucede a las 4 horas y en cerdos a 1 hora. La capacidad de cohesión se traduce en la facilidad de disolución de las moléculas cortas de actina y miosina que se mantienen separadas por contener aún ATP en el tejido muscular. Esta propiedad se puede aprovechar para la transformación de la carne en embutidos (Lieven, 1985).
Al examen físico de los músculos del brazo y la espalda de las carcasas de llama Condori (2000) observó que la rigidez cadavérica es instaurada por completo entre 16 a 20 horas transcurridas después de la muerte del animal (Figura 4) y un pH que fluctúa entre 5,78. Este resultado es similar al señalado por Penny (1984) y Charley (1995) quienes coinciden en mencionar que la rigidez se instaura aproximadamente hasta las 24 horas en vacunos y mucho menos tiempo en cerdos y según Téllez(1992) el pH puede descender hasta un pH = 5.4. Se ha comprobado que cuando se establece la rigidez cadavérica se produce una disminución de la flacidez de la carne (Penny 1984 citado por Condori 2000). La rigidez cadavérica culmina cuando las articulaciones vuelven a ser flexibles a la presión (Aldana 1984). En la carcasa de llama ocurre de 32 a 42 horas después de la muerte del animal.
La rigidez cadavérica en carcasas de llama tiene una duración entre 16 a 22 horas, resultado similar a los reportados por CISA (1996) que señala una duración entre 20 a 24 horas. Después de este tiempo la presión ejercida en las articulaciones de la carcasa rompen el complejo actomiosina que de acuerdo a Varnam y Sutherland (1995) citados por Condori (2000) es el período en el que la maduración de la carne ha concluido. Lo que quiere decir que la carne esta lista para su consumo en fresco.
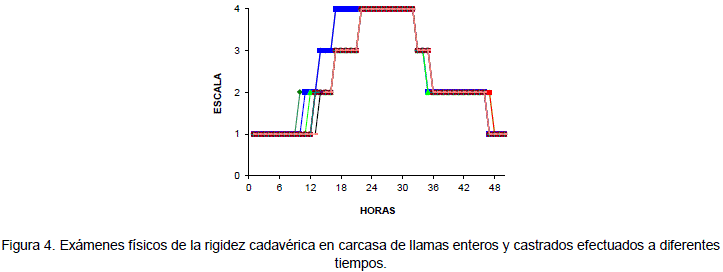
CM: Articulación de la espalda y brazo de carcasa con movimiento (CM) (carne en caliente)
LM: Articulación de la espalda y brazo con leve movimiento (LM) (pre rigidez)
MLM: Articulación de la espalda y brazo de carcasa con muy leve movimiento (MLM) (pre rigidez)
SM: Articulación de la espalda y brazo sin movimiento (SM) (instauración de la rigidez cadavérica)
Glicolisis post mortem
La glicolisis es un proceso anaeróbico irreversible, que ocurre cuando el oxígeno es permanente removido del músculo del animal muerto. La secuencia química que sigue al glicógeno es la conversión esencialmente en acido láctico, tanto en post mortem o en vivo, ante la ausencia del oxígeno temporalmente comienza la inadecuada provisión de energía al musculo, excepto cuando el animal haya sufrido un proceso de inanición o ejercicio antes de la matanza, en este caso se puede apreciar la disminución de reservas de glicógeno en el musculo.
La conversión de glucógeno en ácido láctico continua hasta que el pH desciende, y los enzimas provocan una ruptura lo que la inactiva. En los mamíferos el pH está alrededor de 5.4 a 5.5, y a estos niveles el glucógeno es considerado ausente (Bate - Smith Lawrie, 1955)
En la canal y en varios músculos de esta, se pueden tener diferentes valores de temperatura post morten, esto se relaciona con la temperatura del medio del medio, como resultado de estos valores la glicolisis post mortem, tiene la tendencia de que exista una mayor glicolisis cuando la temperatura es menor y viceversa.
La glicolisis es un proceso donde el musculo comienza a distenderse, es importante hablar del proceso de rigor mortis que recientemente es tomado en cuenta. El rigor mortis tiene una alta correlación con la desaparición del ATP del músculo. En ausencia del ATP, la actina y la miosina, combinan un cambio rígido a actomiosina, la que comprende los siguientes periodos (fase de inicio y fase rápida), en el primero mantiene un bajo nivel. El tiempo del proceso de la fase rápida o de rigor mortis, depende directamente de los niveles de ATP (temperatura), inmediatamente después del periodo de post mortem, es donde comienza el periodo no contráctil, con bajos niveles de ATPasa activando la miosina.
El nivel de ATP puede ser mantenido por algún tiempo por una resintesis de ADP y Creatina fosfato (CP) donde las reservas de CP son utilizadas en la glicolisis post mortem y puede resintetizar ATP, pero este es inactivo y no cubre los niveles que faltan, esto pasa generalmente con pequeñas cantidades de glicógeno, con este glicógeno de resintesis de ATP que proviene de la glicolisis no mantiene los niveles suficientemente altos y se prevé la formación actomiosina (Lawrie, 1985).
Conociendo la temperatura inicial de almacenamiento del glicógeno y los niveles iniciales de ATP y CP, el tiempo del proceso de rigor mortis, puede ser precedido por el proceso de rigor mortis y acompañado por el proceso de capacidad de retención de agua, más el pH y la desnaturalización de las proteínas sarcoplasmatica. Cuando el rigor mortis comienza con elevado pH y una menor capacidad de retención de agua, es cuando desaparece el ATP y comienza la formación de actomiosina.
Relación entre pH, capacidad de retención de agua y rigidez cadavérica
Existe una estrecha relación entre la capacidad de retención hídrica, pH y la rigidez cadavérica. Según Mora et al (1994) citado por Condori et al (2003) señala que cuando se establece la rigidez cadavérica la carne alcanza la mínima capacidad de retención hídrica en el punto isoeléctrico de las proteínas. Este punto es el pH en el cual todas las cargas en la carne están equilibradas siendo las cargas positivas igual a las negativas, en general las carnes presentan un pH de 5 a 5.4 en este punto.
Tomando en cuenta este aspecto en la carne de llama se alcanza la rigidez cadavérica a un pH de 5.78 el cual es levemente superior al reporte del autor anteriormente mencionado. Sin embargo no coincide con la mínima capacidad de retención hídrica, la diferencia se atribuye al tipo de músculo utilizado para la determinación de este parámetro (Condori, 2000). El establecimiento de la rigidez cadavérica se manifiesta dependiendo del tipo de músculo, por lo tanto no es igual para músculos de la pierna comparados con músculos del lomo (Aldana 1984).
Maduración de la carne
La suavidad de la carne es un factor muy importante que afecta a la calidad, la suavidad aumenta con la maduración durante el almacenamiento (desarrollo de la rigidez cadavérica). Los factores más importantes que están implicados en la velocidad del ablandamiento es la proteólisis y el aumento de la presión osmótica después de la muerte del animal (Varnam y Sutherland, 1995 citado por Condori, 2000). A este proceso la temperatura tiene un efecto importante para la velocidad de ablandamiento de la carne según Penny (1984) la elevada temperatura de almacenamiento aumenta la velocidad de ablandamiento, sin embargo esta velocidad varia en las distintas especies.
La maduración de la carne está íntimamente relacionada con la fragmentación de las miofibrillas consecuentemente con el ablandamiento de la carne. El debilitamiento de la estructura miofibrilar como resultado de la degradación de los discos Z es un factor importante en la reducción de la dureza en el proceso de maduración (Penny, 1984).
La conversión de los músculos en carne
Los términos de carne y musculo son diferentes porque existe distinción entre ellos pero no son bien enfatizados, Carne refleja la estructura química y natural de los músculos pero después del proceso de post mortem, aspectos que detallan una serie de cambio químicos y bioquímicos, iniciados en un animal muerto, son los detalles de la conversión de musculo a carne (Lawrie, 1985). Otro concepto de carne lo define como un producto animal usado en la alimentación; en la práctica esta definición es restringida a mas de 3000 especies de mamíferos, en la cual están incluidas a parte de los músculos, los órganos como el hígado los riñones, las membranas y otros tejidos. Se debe considerar una gran variabilidad en las propiedades de la carne, así como refleja diferencias en la composición y condición del tejido muscular durante el proceso post mortem aspecto que es gradual y bien conocido Lawrie (1985).
Cambios bioquímicos en el músculo durante la maduración
La maduración de la carne comienza cuando el músculo entra en rigor y se liberan iones de Ca+2 del retículo sarcoplasmico. Cuando la concentración de Ca alcanza un nivel elevado se activa el CAF (proteasa neutra activada por el Ca) que ataca lentamente a la troponina T y a las proteínas de la región de la línea Z. Con el paso del tiempo, la degradación de las proteínas se hacen notorias y se alteran las propiedades de cohesión de las proteínas miofibrilares. Estos cambios en la organización miofibrilar acompañan a una reducción considerable de la dureza (Penny 1984). Al respecto Varnam y Sutherland (1995) citados por Condori (2000), señalan que la desaparición de la troponina T y la degradación de la desmina son indicadores de que se producen alteraciones estructurales en la miofibrilla así como en el disco Z. Específicamente, la degradación de la desmina conduce a la fragmentación de las miofibrillas. La degradación proteolítica de las proteínas de las miofibrillas musculares es el principal mecanismo de ablandamiento de la carne.
En el ablandamiento se ha demostrado la actividad de dos enzimas, las calpainas (proteínas activadas por el Ca) y cistein-proteinasas lisosomales (catepsinas) frente a la miofibrilla. Las calpainas son más activas a un pH por encima de 6 mientras que las catepsinas son más activas a valores de pH más bajos. Por esta razón, se deduce que las calpainas activadas por la liberación de los iones Ca, desde el retículo sarcoplásmico, son más importantes durante las primeras etapas de la glucólisis. Las catepsinas liberadas de los lisosomas a pH ácido se convierten en las responsables de la proteólisis a pH por debajo de 6 (Varnam y Sutherland, 1995 citados por Condori, 2000).
Existe degradación de la miosina a temperaturas superiores a 15 °C, por esta razón, se ha postulado que las calpainas son responsables de la proteólisis durante la maduración a bajas temperaturas. Las catepsinas son las más importantes durante la maduración prolongada a elevadas temperaturas sin embargo es probable que la acción de las dos enzimas sea sinérgica (Varnam y Sutherland, 1995 citados por Condori, 2000).
El efecto de la edad y castración en la calidad de la carne
Inchausti y Ezequiel (1980) citados por Condori (2000) indican que en la calidad de la carne de bovino (Bos taurus) influye la edad, a mayor edad el tejido muscular adquiere mayor firmeza, y por lo tanto la carne resulta menos tierna. Los animales de 18 a 36 meses en razas precoces proporcionan carnes de mejor calidad. Sin embargo, los animales de mayor edad pueden brindar carne de buena calidad aunque más dura por el mayor ejercicio. En este caso la grasa no es intersticial sino de cobertura o depósito.
Por otro lado la edad cronológica a la que el ganado alcanza la maduración fisiológica no es constante ni fija. En becerros, el crecimiento muestra una pronta maduración y uniformidad hasta los 14.5 meses. A medida que el crecimiento esquelético y muscular se hace más lento, la acumulación de grasa se incrementa. Al aumentar la grasa de depósito disminuye la porción comestible de la canal por tanto a mayor edad mayor grasa de depósito (Neuman 1989 citado por Condori 2000). Además la carne de animales viejos llega a ser de menor calidad en cuanto a terneza y composición de grasa y se destina a la elaboración de productos cárnicos transformados (Varman y Sutherland, 1995). En una experiencia propia Rivera et al. (1996) citado por Condori (2000) señala que la carne de camélidos viejos de preferencia está destinado a la elaboración de charque y la carne de ganado joven es destinada al consumo en fresco.
En cuanto a la castración de los machos es un hecho muy conocido que las características organolépticas de la carne cambian con la edad de los animales faenados pues el sabor de la carne puede ser influenciado por la presencia de feromonas cuando los machos llegan a la madurez sexual, los cuales están ausentes en machos castrados (Gerken, 1998). Se ha encontrado en una prueba de degustación (panel test) realizada con carne de llamas castrados y sin castrar faenados a los 25 meses de edad, que existe preferencias de los degustadores (sin entrenar) hacia la carne de animales castrados (Gerken et al., 2000).
Composición química y bioquímica de la carne
Análisis químico de la carne
El análisis estadístico confirma que existe una diferencia altamente significativa para el factor edad para todas las variables de respuesta consideradas en el análisis químico de la carne. En la Tabla 9 se presenta el análisis de varianza y el efecto de los diferentes factores en estudio.

El análisis de varianza de componentes químicos (Tabla 9) de la carne de llama refleja que no hay diferencias significativas para el efecto de condición y la interacción edad xcondición en contenido graso, humedad, ceniza y colesterol, los resultados de los análisis de varianzas de los componentes químicos se observan en la Tabla 8.
En la Tabla 10, se reporta los valores del porcentaje de humedad en diferentes periodos de sacrificio. El promedio del porcentaje de humedad 71.49 es similar al reportado por Pinto (1975) 69.17%; y Vilca (1991) que hace un resumen de trabajos encontrando un rango desde 69,17% a 73.83%, para carne de llama.
De la prueba de comparación de medias para el factor edad se desprende que el mayor porcentaje de humedad se presenta a la edad de 13 meses y el menor porcentaje de humedad se presenta a la edad de 16 meses para enteros y castrados.
En la Tabla 9, se reporta los valores promedios de porcentaje de grasa, humedad, ceniza y colesterol, los resultados del porcentaje de grasa y ceniza en la carne de llama tiene un valor promedio de 3.6% y 1.42% respectivamente estos resultados son similares a los encontrados por Pinto (1975) 3.69% y 1.41 % para grasa y ceniza respectivamente y también se encuentra en el rango de valores resumidos por Vilca (1991) 1.21% a 4.8% para grasa y 1.17 a 1.71% para ceniza; sin embargo los valores obtenidos por Torres (1998) para porcentaje de grasa 0.57% en carne de llama, discrepa con los datos hallados en el presente trabajo se atribuye esta diferencia marcada a la metodología empleada para la determinación de este componente.
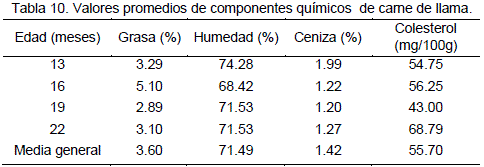
De la prueba de comparación de medias en edades para el componente porcentaje de ceniza se deduce que a la edad de 13 meses la carne de llama presenta un mayor contenido de ceniza. De acuerdo a la comparación
de medias para el factor edad el mayor contenido de grasa se presenta a los 16 meses el cual tiene una diferencia altamente significativa con las demás edades.
En la Figura 5 se presentan los valores de % grasa y % de humedad observándose que a los trece meses de edad los animales presentan un valor promedio de porcentaje de humedad de 74,28 % para enteros y castrados, valores que disminuyen a los 16 meses contrariamente al contenido de grasa que tiende a aumentar en este periodo, a los 19 meses de edad la tendencia de ambas variables es opuesta y posteriormente tiende a estabilizarse.
Esta tendencia es consistente con Garnica (1993) quien indica que las proporciones de agua en los músculos es menor cuanto mayores sean los niveles de grasa, para alpacas, sin embargo el curso de la curva del porcentaje de grasa no coincide con la descrita por el mismo autor en relación a una mayor edad, mayor contenido de grasa en carne de alpaca. El aumento del valor de la variable porcentaje de grasa a los 16 meses se atribuye a la época de faena el mismo que coincide con una estación seca del año.
Los resultados del contenido de colesterol en carne de llama son presentados en el cuadro No. 9 el cual muestra un promedio de 55.7 mg/100g este resultado es similar al reportado por Garnica (1993) 56.49%, para alpacas y es inferior a la concentración promedio de 88.12 mg/100g para ovinos y 76.53 mg/100g para vacunos, valores reportados por el mismo autor.
De la prueba de comparación de medias para las edades se extrae que el mayor contenido de colesterol se encuentra a una edad de 22 meses y el menor contenido a una edad de 19 meses. En la Tabla 9, se aprecian diferencias altamente significativas entre edades con respecto al porcentaje de proteína. Además confirma las diferencias como altamente significativas para el efecto de la interacción edad por condición.
En la Tabla 10, se presentan los valores promedios de porcentaje de proteína a diferentes edades y la media general para la carne de llama, 24.64 y 25.69% para enteros y castrados respectivamente, estos valores son consistentes con los obtenidos por Pinto (1975) 24.82% de proteína y Vilca (1991) con 19.46 a 24.82%, sin embargo existe una leve diferencia con Torres (1998) que presenta un valor de 21.49% de proteína.
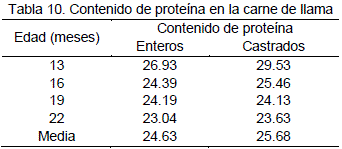
De la prueba de comparación de medias para los tratamientos se deduce que el mayor contenido de proteína es a los 13 meses de edad para enteros y castrados y el menor contenido de proteína se registró a una edad comprendida entre 19 y 22 meses tanto para enteros y castrados esto significa que a medida que avanza la edad del animal el contenido proteico en la carne tiende a disminuir, esta aseveración coincide con lo señalado por Garnica(1993) quien indica que a mayor edad menor contenido de proteína en carne de alpaca.
El análisis de varianza (Tabla 9) muestra un efecto significativo para el factor condición y la interacción edad por condición, al análisis de los efectos simples de la interacción (Tabla 10) se obtiene los siguientes resultados.
La composición química de la carne corresponde a los nutrientes que posee en términos de porcentuales, la carne está constituida por el músculo estriado del animal que viene a ser, aproximadamente el 35% de su peso y que tiene las características que a continuación se expresan en las Tablas 11 y 12:

La composición química de la carne varía según la especie y las distintas partes de donde procede la carne, la musculatura contiene, en líneas generales la composición de la carne puede ser aproximadamente, como muestra la Tabla 11, 75% agua, 19% proteínas, 3.5% de sustancias no proteicas (Hidratos de carbono), 2.5% de grasa, 1% de minerales y escasas vitaminas, Lawrie (1985).
En la Tabla 12, se muestra que los porcentajes de componentes químicos, de los músculos específicos, y por especie, lo cual nos indica que estos componentes no solo varían de la carne de una especie a otra, sino que también varían de acuerdo al musculo, así como también la composición química es diferente en las vísceras.
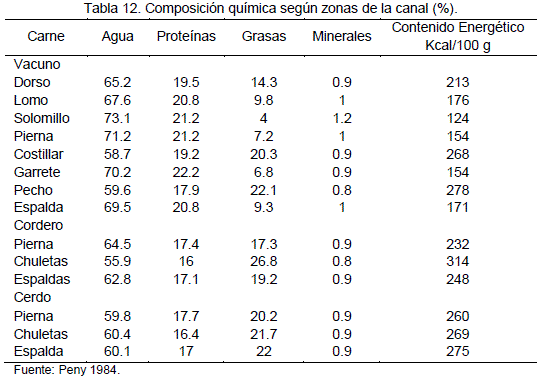
Pinto (1975) y Vilca (1991) presentan un resumen de la composición química de la carne de llama obtenida por diferentes autores, mencionan valores promedios de la composición química de carne de llama de ambos sexos y diferentes edades y (Tabla 13).

Para la carne de llama Condori et al (2003) presenta los siguientes valores en muestras extraídas del músculo longissimus dorsi. El análisis químico se realizó en SELADIS. Los valores promedios de los componentes químicos de la carne de llama son presentados en la Tabla 14.
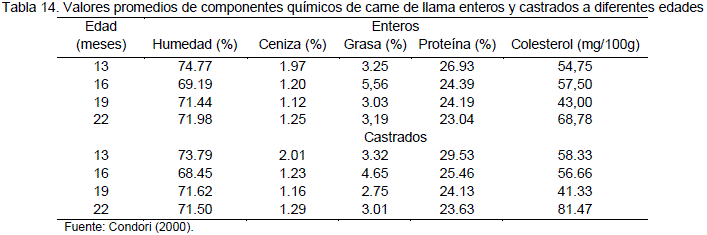
En la Tabla 15, se presenta el resumen del análisis de varianza y el efecto de los diferentes factores en estudio. El análisis estadístico confirma que existe una diferencia altamente significativa (p<0.01) para el factor edad para todas las variables de respuesta consideradas en el análisis químico de la carne. No hay diferencias significativas para el efecto de condición e interacción edad por condición para el contenido graso, humedad, ceniza y colesterol. La covariable peso inicial no tiene efecto significativo (p>0.01) en los componentes químicos de la carne (Condori, 2000).

Proteínas
Las proteínas ocupan un lugar preferente por sus características nutritivas, el porcentaje en las carnes resulta superior al de otros alimentos, especialmente los de origen vegetal. Las proteínas de las carnes se caracterizan por su extraordinaria digestibilidad, sin embargo, las proteínas de las vísceras, especialmente de riñón, bazo y pulmón resultan de digestión difícil.
Las proteínas desempeñan un mayor número de funciones intracelulares de todos los seres vivos. Forman parte de la estructura básica de los tejidos (músculos, tendones, piel, uñas, etc.), desempeñan funciones metabólicas y reguladoras (asimilación de nutrientes, transporte de oxígeno y de grasas en la sangre, inactivación de materiales tóxicos o peligrosos, etc.). Definen la identidad de cada ser vivo en base a la estructura del código genético (ADN) y de los sistemas de reconocimiento de organismos extraños en el sistema inmunitario (Lawrie, 1985).
Las proteínas son moléculas de gran tamaño formadas por largas cadenas lineales de aminoácidos. Existen unos 20 aminoácidos distintos, que pueden combinarse en cualquier orden. Una proteína media está formada por unos cien o doscientos aminoácidos. Según la secuencia de aminoácidos las propiedades pueden ser totalmente diferentes. Tanto los glúcidos como los lípidos tienen una estructura relativamente simple comparada con la complejidad y diversidad de las proteínas (Varam et al., 1995).
Cada especie animal o vegetal tiene su propio tipo de proteínas, incompatibles con los de otras especies, para asimilar las proteínas estas deben ser fraccionadas en sus diferentes aminoácidos. Esta descomposición se realiza en el estómago e intestino, bajo la acción de los jugos gástricos y los diferentes enzimas. Los aminoácidos obtenidos pasan a la sangre, y se distribuyen por los tejidos, donde se combinan de nuevo formando las diferentes proteínas específicas de nuestra especie.
Las proteínas del cuerpo están en un continuo proceso de renovación. Uno se degradan hasta sus aminoácidos constituyentes y otro se utilizan estos aminoácidos conjuntamente los obtenidos en la dieta para formar nuevas proteínas, en base a las necesidades del momento. A este mecanismo se le llama recambio proteico (Lawrie, 1985).
La proteína es imprescindible para el mantenimiento de la vida, es la principal causa del consumo energético en reposo, en ausencia de glúcidos en la dieta de la que proviene la glucosa, es posible obtenerla a partir de la conversión de ciertos aminoácidos en el hígado. El sistema nervioso y los leucocitos de la sangre no pueden consumir otro nutriente que no sea glucosa, el organismo puede degradar las proteínas de nuestros tejidos menos vitales para obtenerla, las proteínas específicas que se pueden encontrar en la carne son como se reflejan en la Tabla 16.
Cuando las proteínas son consumidas en exceso, los aminoácidos constituyentes son utilizados para obtener energía. A pesar de la versatilidad de las proteínas, los humanos no estamos fisiológicamente preparados para una dieta exclusivamente proteica. Estudios realizados en este sentido pronto detectaron la existencia de importantes dificultades neurológicas.
Garnica (1993a) indica que se requiere conocer muchos factores como la edad y el régimen alimenticio que influyen sobre la carne. A medida que el animal envejece, el porcentaje de proteína disminuye, sin embargo, las diferencias no son significativas. El porcentaje de proteína es menor en carnes grasas, que en las carnes magras. Para Varnam y Sutherland (1995) citados por Condori (2000) la calidad de la proteína en la carne es alta por los tipos y proporciones de los aminoácidos son similares a los que requiere el crecimiento y mantenimiento del tejido humano. De los aminoácidos esenciales para el hombre la carne aporta una cantidad importante de lisina y treonina y cantidades adecuadas de metionina y triptófano.

La carne en general contiene del 15 al 20% de proteína, el contenido graso varía de 5 al 40% dependiendo del tipo, raza, especie, alimentación y edad del animal. Los cortes magros de músculo son excelentes fuentes de fósforo y hierro, el contenido de agua es aproximadamente cerca del 75% (Charley, 1995).
En la Tabla 17, se presentan los valores promedios del contenido de proteína, que refleja una diferencia altamente significativas (P<0.01) para el efecto de la edad y condición de los animales. No significativa (P>0.01) para la interacción de edad por condición.
La prueba de comparación de medias (Duncan) de la Tabla 17, refleja que el mayor porcentaje de proteína se presenta en carne de llamas castrados de 13 meses de edad. Que presentan un valor promedio de 29.53%, de proteína, seguido de carne de llamas enteros de la misma edad que contienen 26.93% de proteína. La carne de los animales de 16 a 19 meses de edad entre castrados y enteros que no muestran diferencias significativas, presentan un valor promedio de 24.54% de proteína. Otro grupo de 16 a 22 meses de edad presenta un promedio de 23.87% de contenido proteico. Estos dos últimos valores son similares con los obtenidos por Pinto (1975) 24.82% de proteína en carne de llama y también se encuentra dentro del rango de valores resumidos por Vilca (1991) 19.46 % a 24.82%.

Se evidencio una diferencia notable con otras especies como el vacuno (21,01%), ovino (18.91%), porcino (19.37%) y alpaca (21.88%), mencionado por Rivera et al. (1994) citado por Condori (2000) lo cual confirma una diferencia marcada a favor de la carne de llama. Otros valores hallados por Ramírez (1973) son 17.5% y 15,7 % de contenido proteico para vacuno y ovino respectivamente. Madueño y Leyva (1988) citados por Tichit (1991) indican valores de 21 y 17% de proteína para vacuno y ovino respectivamente. Por lo tanto queda nuevamente confirmado la superioridad de la carne de llama referente al contenido de proteína.
En la figura No. 5, se observa la tendencia del contenido de proteína de la carne de llama en enteros y castrados el cual presenta una marcada disminución a los 22 meses. Significa que a medida que avanza la edad de animal el contenido proteico en la carne tiende a disminuir. Esta aseveración coincide con Garnica (1993a), quien señala que a mayor edad menor contenido de proteína en carne de alpaca.

Ecuación ajustada de llamas enteros: y = -6.8883Ln(x) + 44.2
Ecuación ajustada de llamas castrados: y = -11.162Ln(x) + 57.425
La relación observada entre la mayor ganancia de peso diario a los 14 meses de edad para enteros y castrados y el alto contenido proteico a los 13 meses de edad. Es corroborado por Di Marco (1993) quien explica que la ganancia de peso en animales jóvenes es debida a la mayor retención de proteína. En cambio en animales adultos la mayor ganancia de peso se debe al incremento de grasa de depósito en decremento de la retención proteica.
El alto valor proteico observado en animales jóvenes de 13 meses de edad se atribuye a una menor degradación de proteína. La cantidad de proteína retenida en el tejido muscular está en función del balance entre las tasas de síntesis y degradación, por lo tanto el mayor contenido no se debe exclusivamente a una mayor síntesis sino a una menor degradación. Este proceso de síntesis y degradación está controlado por hormonas específicas Di Marco (1993). Según Trenkle (1983) citado por el mismo autor indica que una de las funciones importantes de la somatotropina, una de las hormona del crecimiento, es favorecer la retención proteica sin alterar su degradación. El efecto de esta hormona es aumentar el transporte de aminoácidos dentro de las células y principalmente determina el tamaño de un animal, presumiblemente es una hormona que actúa en el crecimiento acelerado del animal joven.
Balance de nitrógeno
El componente más preciado de las proteínas es el nitrógeno, con él podemos reponer las pérdidas obligadas que sufrimos a través de las heces y la orina. A la relación entre el nitrógeno proteico que ingerimos y el que perdemos se le llama balance nitrogenado. Debemos ingerir al menos la misma cantidad de nitrógeno que la que perdemos. Cuando el balance es negativo perdemos proteínas y podemos tener problemas de salud. Durante el crecimiento o la gestación, el balance debe ser siempre positivo.
El conjunto de los aminoácidos esenciales sólo está presente en las proteínas de origen animal. En la mayoría de los vegetales siempre hay alguno que no está presente en cantidades suficientes. Se define el valor o calidad biológica de una determinada proteína por su capacidad de aportar todos los aminoácidos necesarios para los seres humanos. La calidad biológica de una proteína será mayor cuanto más similar sea su composición a la de las proteínas de nuestro cuerpo. De hecho, la leche materna es el patrón con el que se compara el valor biológico de las demás proteínas de la dieta.
Lípidos
La Tabla 18, muestra los lípidos que se clasifican de la siguiente manera:

Aunque el contenido de grasas saturadas es variable en las diferentes especies todas, por su poder acidificante, puede ser perjudicial para quienes sufren trastornos hepáticos y renales, la Tabla 19 muestra la variabilidad del porcentaje de grasa, en la carne de diversas especies animales.
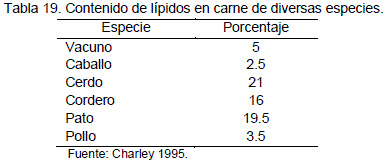
No existen diferencias significativas (P>0.01), para el contenido de grasa, el análisis de varianza refleja que no existen diferencias para el efecto de condición y la interacción edad por condición. Sin embargo existe una diferencia altamente significativa (P<0.01) para el efecto de la edad.
En la Tabla 20, se presentan los valores promedios del contenido de grasa y de acuerdo a la comparación de medias (Duncan) para el factor edad, el mayor contenido de grasa se presenta a los 16 meses (Tabla 20).
Tabla 20. Valores promedios del contenido de grasa en carne de llama a diferentes edades sin distinción de la condición.
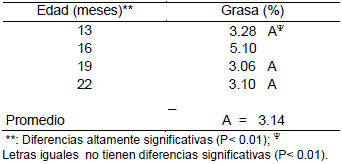
Los valores promedios del contenido de grasa son de 3.14% para edades 13, 19 y 22 meses, similares a los reportados por Pinto (1975) 3.69 % para llamas de diferentes edades. También se encuentran en el rango de valores resumidos por Vilca (1991) de 1.21 a 4.8% de contenido de grasa en carne de llama.
El porcentaje de grasa promedio a los 16 meses es de 5.1 %, diferente a los valores señalados anteriormente por los dos autores. Sin embargo en forma general los valores obtenidos por Torres (1998) 0.57% para contenido de grasa en carne de llama discrepa sustancialmente con los datos hallados en el presente trabajo. Esta diferencia marcada puede atribuirse a la metodología empleada para la determinación de este componente.
Aunque el contenido de grasas saturadas es variable en las diferentes especies todas, por su poder acidificante, puede ser perjudicial para quienes sufren trastornos hepáticos y renales, la Tabla 19 muestra la variabilidad del porcentaje de grasa, en la carne de diversas especies animales.
En otras especies Rivera et al. (1996) citado por Condori (2000) menciona niveles de grasa de 4.84% para vacunos, 6.53% en ovinos, 5.13 % en alpacas y 20.06% para porcinos. La carne de llama presenta valores notablemente inferiores con excepción de aquellos animales de 16 meses (5.1%) cuyos valores se asemejan a la carne de alpaca y de vacuno.
El bajo nivel de grasa en la carne de llama se atribuye a las condiciones medio ambientales y la poca disponibilidad de alimento o poca vegetación en las zonas donde se desarrolla la crianza de este camélido. Es probable que esta situación tenga efecto en la composición química. Puesto que cuando el nivel nutricional disminuye en el ganado bovino, los niveles de grasa son las que se ven más afectadas, aunque mucho más que la proteína, por lo tanto el animal es más magro. Como consecuencia de ello se tiene menor grasa subcutánea y grasa intramuscular consecuentemente menor brillo de la carne Di Marco (1993). Esta podría ser la razón por la que la carne de llama tiene apariencia un tanto oscura y pálida ya que el color varia con la especie Rivera et al. (1994).
En la figura 6, se presentan los valores de porcentaje de grasa y porcentaje de humedad en la carne de llama conforme avanza la edad sin distinción de la condición. Se observa que a los trece meses de edad los animales presentan un valor promedio de 74.28 % de humedad para enteros y castrados. Valores que disminuyen a los 16 meses, contrariamente al contenido de grasa que tiende a aumentar en este período. A los 19 meses de edad la tendencia de ambas variables es opuesta y posteriormente tienden a estabilizarse.
La tendencia señalada coincide con lo que indica Garnica (1993a), que la proporción de agua en los músculos es menor cuanto mayor es el nivel de grasa en carne de alpacas. Existe una divergencia en cuanto al contenido de grasa respecto a la edad por que señala que a mayor edad, mayor contenido de grasa en carne de alpaca. Esta diferencia podría deberse al rango de edad de los animales de 18 a 53 meses de edad obteniendo valores de 2.75 a 3.13% de grasa.

El alto valor del porcentaje de grasa a los 16 meses se atribuye a la época de faeneo (mes de junio). Suponiendo que meses antes los animales estuvieron relativamente con buen forraje por la época lluviosa entonces llegaron al sacrificio con índices de alto contenido de grasa intramuscular. Esta afirmación es consistente con Di Marco (1993) quien señala que la alimentación es uno de los factores externos que puede afectar el contenido y distribución de la grasa.
Ácidos grasos
El cuerpo puede fabricar la mayoría de los ácidos grasos (componente de las grasas) que necesita (ácidos grasos no esenciales), por lo tanto no es preciso incorporarlos con los alimentos; pero hay un tipo de ácidos grasos que el cuerpo no puede fabricar y son los llamados esenciales, imprescindibles para una buena salud y que solo se puede obtener a través de la alimentación.
Se trata de las familias de omega-3 y omega-6, que necesita el cuerpo en poca cantidad, 15 gr/día, el equivalente a una cucharada sopera de aceite vegetal.
Funciones de omega-3 y omega-6: Forman parte de sustancias que actúan en todos los procesos corporales. Aseguran el crecimiento y desarrollo apropiado en infantes y niños. Forman parte de todas las membranas de las células. Intervienen en la regulación de los niveles del colesterol sanguíneo. Regulan la presión arterial. Disminuyen el riesgo de infarto y cáncer. Aumentan las defensas del organismo. Protegen la piel. Favorece la cicatrización de heridas. Se necesita para el buen funcionamiento hepático y renal (Varman y Sutherland 1995).
Los ácidos grasos omega-3 son más poli insaturados, se encuentran en cantidades importantes en las semillas de lino, pescados de mar, aceite de pescado, frutas secas y soja. La falta de omega-3 se asocia a problemas neurológicos, pérdida de memoria, hormigueo de brazos y piernas, disminución de la visión, baja las defensas del organismo, es decir, menor resistencia a las infecciones), aumenta el colesterol en sangre y retrasa el crecimiento en los niños.
Los omega-6 se encuentran en la mayoría de los aceites vegetales: maíz, sésamo, en el germen de trigo y las frutas secas. La falta de omega-6, puede manifestarse en problemas de piel: eczemas, dermatitis seborreica, pérdida anormal de cabello, problemas hepáticos, renales, cicatrización de heridas, puede afectar el sistema circulatorio, aumentando el colesterol sanguíneo, provocando hipertensión, endurecimiento y obstrucción de las arterias.
Grasa intramuscular, llamada también tejido adiposo que en conjunto son los esteres del glicerol y los ácidos grasos que llegan a ser el 99% de la grasa intramuscular, aunque otros tejidos metabólicos activos, tienen un considerable contenido de fosfolípidos e insaponificables constituyentes como el colesterol. Solo tres o cuatro ácidos grasos están presentes sustancialmente en la carne de los animales (oleico, palmítico, esteárico) y cuatro tipos de glicéridos, Berg (1978).

Los esteres del glicerol y los ácidos grasos llegan a conformar el 99% de la grasa intramuscular, aunque otros tejidos metabólicos activos, tienen un considerable contenido de fosfolípidos e insaponificables constituyentes como el colesterol. Solo tres o cuatro ácidos grasos están presentes sustancialmente en la carne de los animales (oleico, palmítico, esteárico) y cuatro tipos de glicéridos.

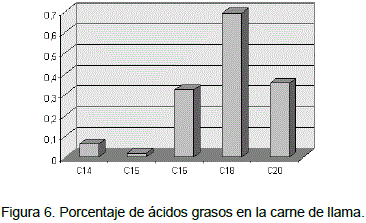
Los ácidos grasos en la carne de llama reportados en el presente trabajo, de los cuales se indica un promedio para enteros y castrados el cual detalla en la Tabla 22 y que a diferencia de las carnes de otras especies animales de se puede apreciar que existen rangos como por ejemplo en el caso del acido miristico que va desde 0.9% en vacunos a 3.26% en el cerdo, o como en el palmítico que va de 22.34% en cerdos a 24.14 en pollos, así como para esteárico de 4.28% en el cerdo a 13.18% y el araquidonico que va de 0.67% en el pollo a 2.30 en el cordero.
Los rangos señalados anteriormente para ácidos grasos presentes en la carne de diversas especies, en referencia a la carne de llama como se puede apreciaren la Figura 6, estos no se encuentran dentro los rangos mencionado para otras especies, valores que están por muy debajo de la media de algunos ácidos grasos, donde el promedio más alto corresponde a acido palmítico con un 0.32% y el más bajo para el acido pantodecanoico con 0.18%.
Colesterol
Según Garnica (1993b), los problemas cardiovasculares en el hombre, es por alta concentración de colesterol en la alimentación. Las grasas saturadas de alimentos como las carnes, lácteos y yemas de huevo tienden a aumentar el colesterol en la sangre negativamente, en cambio las grasas no saturadas tienden a bajar el nivel. La carne de alpaca contiene una concentración baja de colesterol 56,49 mg/100g en comparación con ovinos de razas mejoradas que presentan un rango de valor de 70 a 105 mg/100g y en vacunos de 90 a 125 mg/100g según Bender(1973) citado por Garnica (1993b).
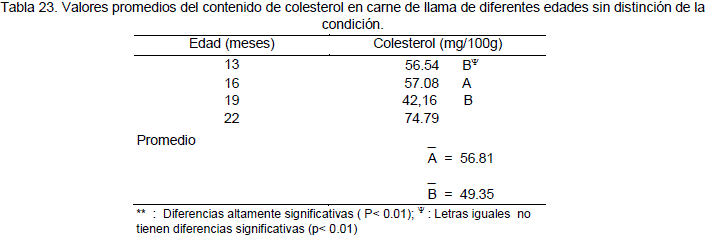
El análisis de varianza refleja que no existen diferencias significativas (P>0.01) para el efecto de la condición e interacción edad por condición. Sin embargo existe una diferencia altamente significativa (P<0.01) para el efecto de la edad. En la Tabla 20, se presentan los valores promedios del contenido de colesterol. De la prueba de comparación de medias (Duncan), en la Tabla 23, se puede notar que el mayor contenido de colesterol se encuentra en la carne de animales de 22 meses y el menor contenido en animales de 19 meses.
En general los resultados obtenidos en el presente trabajo son notablemente diferentes a los valores reportados por Vilca (1991) de 160 mg/100g para carne de llama. Diferencia que se atribuye al método utilizado para su determinación.
Sin embargo el valor promedio reportado en el presente trabajo es de 56.81 mg/100g en llamas de 13 y 16 meses de edad, similar al reportado porGarnica (1993b) 56.49 mg/100g para alpacas. En llamas de 19 meses de edad el contenido de colesterol es 42.16 mg/100g este valores inferior al reporte del autor mencionado. El reporte señala sobre el rango del contenido de colesterol que va desde 56.54 mg/100g a 74.79 mg/100g de carne de llama a los 22 meses, este rango superior es similar al valor reportado por Garnica (1993b), para alpacas y el más bajo para vacunos de 76.53 mg/100g. Sin embargo es inferior comparado con la carne de ovinos que tiene un contenido de 88.12 mg/100g de colesterol valor reportado por el mismo autor.
Los niveles de colesterol en la carne de llama son los más bajos reportados en las especies domésticas, esta aptitud pueda deberse a la rusticidad del animal, la misma que no le permite la acumulación de grasa de cobertura e intramuscular, ya que son animales que utilizan la energía al máximo, lo cual no les permite esta acumulación, ya que deben buscar el alimento recorriendo muchos kilómetros y deben combatir las inclemencias del tiempo del medio de Los Altos Andes.
Contenido de Humedad
Para el contenido de humedad el análisis de varianza refleja que no existen diferencias significativas (P>0.01) para el efecto de la condición e interacción edad por condición. Sin embargo existe una diferencia altamente significativa (P<0.01) para el efecto de la edad.
Los valores promedios del contenido de humedad de la carne de llama a diferentes edades y la prueba de comparación de medias (Duncan), son reportados en la Tabla 24. El mayor contenido de humedad se presenta a la edad de 13 meses y el menor a los 16 meses para enteros y castrados respectivamente.
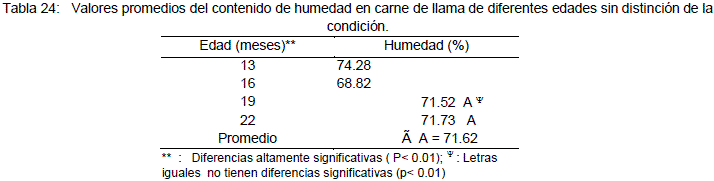
La media del porcentaje de humedad es de 68.82% a los 16 meses, similar al reportado por Pinto (1975) de 69.17% para carne de llama. La canal de animales jóvenes presentan un valor de 74.28% a los 13 meses de edad y los animales de 19 y 22 meses presentan un promedio de humedad de 71.63% (Tabla 18). Valores que coinciden con los Vilca (1991) quien señala un resumen de trabajos, con un rango de 69.17 a 73.83%, para carne de llama.
Los valores obtenidos son similares a carnes de otras especies como en el vacuno y ovino 72.72 y 72.24 % respectivamente mencionado por Rivera et al. (1996). Existe una marcada diferencia con la especie porcina 59.18% mencionado por el mismo autor. Esta diferencia es confirmada por Madueño y Leyva (1988) citados por Tichit (1991) quienes reportan un valor de 47.5 % de humedad para carne de cerdo.
Contenido de Ceniza
En la Tabla 25 se presentan los valores promedios del contenido de ceniza. El análisis de varianza refleja que no existen diferencias significativas (P>0.01) para el efecto de la condición e interacción edad por condición sin embargo existe una diferencia altamente significativa (P<0.01) para el efecto de la edad.
De la prueba de comparación de medias (Duncan) para el componente porcentaje de ceniza se observa que a la edad de 13 meses la carne de llama presenta un alto contenido de ceniza (Tabla 25) el valor promedio alcanza 1.99%.

Tabla 25: Valores promedios del contenido de ceniza en carne de llama de diferentes edades sin distinción de la condición
Diferencias altamente significativas ( P<0.01); * : Letras iguales no tienen diferencias significativas (p< 0.01)
Los valores promedios del contenido de ceniza 1.2 % para edades 16, 19 y 22 meses son similares a los reportados por Pinto (1975) 1.41 % de ceniza. También se encuentra dentro del rango de valores resumidos por Vilca (1991), 1.11 a 1.71 % de ceniza.
El rango de contenido de ceniza de 1.2 % a 1.99 %, comparado con otras especies estas son superiores a los indicados por Ramírez (1973), quien menciona un valor de 0.8 % y 0.9 %, de ceniza para ovinos y vacunos respectivamente. Esta diferencia es atribuible a la mayor tasa de infiltración de grasa en la carne de ovinos y vacunos. Consecuentemente el porcentaje de ceniza es menor en Garnica (1993a).
En alpacas de 18 a 54 meses existe una disminución del contenido de ceniza de 1.23 % a 1.07 % de ceniza (Garnica 1993a), esta afirmación coincide con la tendencia de disminución observada de 1.9 a 1.2 %de ceniza en carne de llama del presente trabajo.
Vitaminas
Las vitaminas se clasifican: vitaminas liposolubles e hidrosolubles. El descubrimiento de la vitaminas se produjo a raíz de la observación de que, mientras que una dieta sintética (con carbohidratos, proteínas, lípidos y minerales, exclusivamente) no mantenía el crecimiento de animales de experimentación, la adición de leche a la mezcla producía un alimento suficiente. El fraccionamiento de la leche permitió encontrar la fracción grasa como la acuosa, ambas indispensables (Bender, 1973).
No es fácil estimar las necesidades de cada vitamina, ya que éstas varían con factores como la edad, el peso, la situación fisiológica e incluso por la influencia de la dieta. La deficiencia severa de algunas vitaminas en la dieta da lugar a la aparición de enfermedades carenciales, que pueden ser muy graves.
Algunas enfermedades carenciales se observan con cierta frecuencia en poblaciones mal alimentadas, otras solamente se han observado en situaciones experimentales: La carencia de vitamina A produce defectos en la visión, y si esta carencia es muy severa, produce xeroftalmia (sequedad de la córnea) que puede terminar en ceguera. La carencia de vitamina D produce defectos en la calcificación de los huesos y, consecuentemente raquitismo (en los niños) u osteomalacia. Puesto que se sintetiza en la piel por la exposición a la luz solar.
La carencia de niacina produce la enfermedad conocida como pelagra. Enfermedad era muy frecuente en algunas poblaciones que basaban su alimentación en el maíz, ya que la niacina presente en él está en una forma no biodisponible.
La carencia de tiamina produce daños neurológicos, y en los casos más serios, la enfermedad conocida como beri-beri. Enfermedad apareció al modificarse los sistemas de descascarillado del arroz, ya la vitamina se concentra en la cascarilla, que en el sistema antiguo parte pasaba con el grano, con el nuevo se perdía totalmente.
La carencia de ácido fólico tiene como consecuencia la aparición de anemia megalobástica, enfermedad que se encuentra con cierta frecuencia entre las mujeres gestantes de los países subdesarrollados, ya que el embarazo aumenta mucho (se estima que alrededor del doble) las necesidades de esta vitamina en particular. La carencia de vitamina C produce alteraciones en la síntesis del colágeno, lo que da lugar al escorbuto. Esta enfermedad fue muy grave entre los marineros de siglos pasados, pero ahora prácticamente no se producen casos.
En los casos de las demás vitaminas. La vitamina K es sintetizada por la propia flora bacteriana, en cantidades más que suficientes para cubrir nuestras necesidades. La vitamina B12 se encuentra en los alimentos de origen animal, y es sintetizada también por las bacterias del tubo digestivo, aunque no está claro el grado de biodisponibilidad de esta última. Las situaciones carenciales están relacionadas o con problemas de absorción (la biotina, y la vitamina B12, en particular) o con dietas muy desequilibradas
Minerales
El porcentaje de cenizas en la carne de alpaca en forma general oscila entre 0.8 a 1.8 %. Se encuentran representadas por el fósforo, potasio, sodio, cloro, magnesio, calcio, azufre, hierro y silicio. Además se encuentran otros oligoelementos en concentraciones bajas como el cobre, zinc etc. La importancia del Ca radica en la ocurrencia de la contracción muscular, en la rigidez cadavérica, en el proceso de maduración de la carcasa y en la capacidad de hidratación de la carne. El Fe participa activamente en la coloración de la carne (Garnica 1993a).
La carne contiene todas las sustancias minerales que son necesarias para el organismo humano, entre las que destacan el Fe y P. Aunque las especies animales no ofrecen diferencias significativas entre sí, en cuanto al aporte de los nutrientes minerales, cabe resaltar la riqueza en fósforo de la ternera y los animales de caza. Se ven las diferencias de composición entre diferentes tipos de carne en la tabla 26.
El Calcio componente fundamental de los huesos, participa en la trasmisión nerviosa y forma parte de la estructura de varios enzimas. El Fósforo junto con el calcio forma la estructura de los huesos y diente, forma parte metabolismo en la obtención y transmisión de energía, mantiene y trasmite el mensaje genético. El Potasio participa en la presión osmótica, especialmente en el interior de las células, y en la trasmisión nerviosa. El Cloro interviene en el equilibrio iónico y osmótico. Forma parte del jugo gástrico. El Sodio participa en la presión osmótica en compartimentos extracelulares, en la transmisión nerviosa y en el equilibrio ácido-base. El Magnesio forma parte de los huesos, es también necesario para la actividad de muchos enzimas, especialmente de aquellos que utilizan ATP. En estos enzimas, el Mg está unido realmente al ATP, y no al propio enzima.
Hierro forma parte de la hemoglobina y mioglobina. Almacena cantidades importantes de este elemento como reserva, asociado a una proteína llamada ferritina. Flúor forma parte de la estructura de los dientes y huesos, aunque no es un elemento estrictamente esencial. Zinc forma parte de bastantes enzimas, como la carbónico-anhidrasa o la fosfatasa alcalina. Cobre forma parte de algunos enzimas, como la tirosinasa. Manganeso forma parte de algunos enzimas, entre ellos la superoxido dismutasa mitocondrial. Selenio forma parte de la selenocisteina, un aminoácido peculiar, (equivalente a la cisteína, pero con azufre en lugar de selenio) presente en la glutatión peroxidasa. Molibdeno forma parte de un cofactor específico necesario para tres enzimas, entre ellos la xantín-oxidasa. Yodo su única función biológica es como componente de las hormonas tiroideas. Cobalto se encuentra exclusivamente formando parte de la vitamina B12. Cromo solamente interviene, en forma de Cr+++, como constituyente del "factor de tolerancia a la glucosa" Existen otros elementos (silicio, boro, vanadio, estaño, arsénico y níquel) de los que no se conoce con precisión su función biológica, si es que la tienen, ni enfermedades carenciales en humanos, puedan desempeñar alguna función en nuestro organismo, Varman (1995). (el silicio, en el desarrollo óseo, el boro en el metabolismo del calcio).
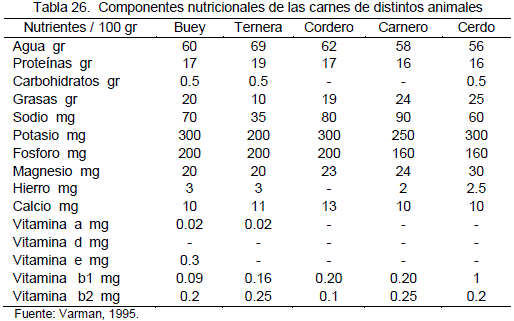
Composición química de las vísceras de llama
En la antigüedad el consumo de las vísceras era de forma habitual, tal es el hecho que como por ejemplo el hígado era suministrado a las personas enfermas aunque en forma cruda. En los últimos años el consumo se ha atenuado, no solo por su difícil conservación, aunque son difíciles de digerir.
Las vísceras son los órganos y partes no musculares de los animales. Están constituidas por fibras más cortas, por lo que su masticación resulta más fácil. Su sabor es más fuerte que el de la carne. Existen dos grandes grupos: las vísceras rojas hígado y riñones y las vísceras blancas sesos, criadillas y tuétanos. Hoy día existe un menor consumo, debido fundamentalmente, al mayor poder adquisitivo de la población (Massi, 1987).
El valor nutricional de las vísceras, el hígado y los riñones tienen un valor nutricional semejante a la carne magra, aunque tienen menos grasa, más col este rol, más agua, más minerales sobre todo Fe, Cu y K, así como más vitamina B12, A, D y C. Los sesos, criadillas y el tuétano tienen un alto contenido en grasas y col este rol, Tabla 27.
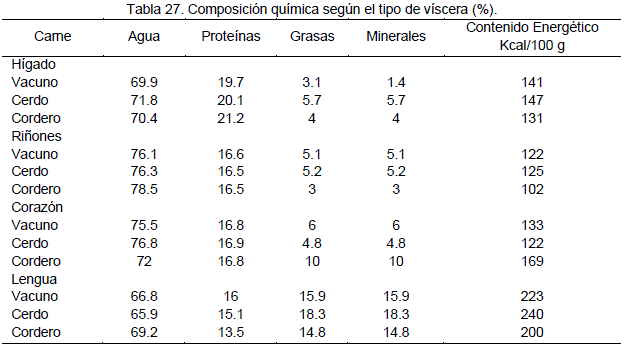
La importancia del análisis químico de las vísceras, tiene singular importancia, porque el consumo de las mismas se realiza cotidianamente dentro las poblaciones donde se crían las llamas. El resumen de las características químicas de los diversos órganos analizados se puede apreciar en la Tabla 28.
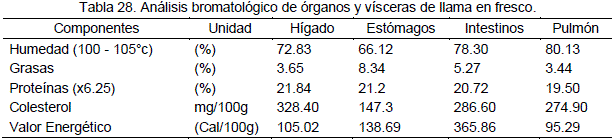
Hígado
Para el caso del hígado en particular, este órgano dentro un contexto general se puede decirque históricamente este órgano era recomendado para combatir la anemia y estados carenciales en general, en humanos por tanto esta víscera es apreciada por su gran contenido nutricional. Massi (1987) reporta los porcentajes de proteína del hígado de diversas especies, 21 % para los bovinos, 23% porcinos, 19% ovinos, mientras que en el presente trabajo se reporta un 21.84% de proteína para el hígado de llama fresco, por tanto la misma se encuentra dentro los parámetros anteriormente mencionados para las otras especies.
Para el contenido de lípidos, el mismo autor reporta un rango de 4 a 5%, mientras que otros autores reportan rangos de 3.1% para terneros, 5.7% para porcinos y 4% para ovinos, para el caso de los camélidos sudamericanos, como la llama se reporta una media de 3.65 %, este porcentaje puede variar de acuerdo a la edad del animal, porque a mayor edad el animal el incremento o la acumulación de lípidos es mayor. El aporte de caloríasen diversas especies se reporta rangos de 121 cal/100 gr en ovinos y de 137 cal/100 gr en porcinos, para el caso de las llamas se reportan una media 105.02 cal/100 gr, valor que es menor en comparación con las otras especies domésticas. Un componente importante es el colesterol que en otras especies se esta se encuentra en una media de 300 mg/100 gr, mientras que para el caso del hígado en el presente trabajo se reporta 328.40 mg/100 gr, valor alto que la media de otras especies domésticas. En el presente trabajo también se reportan datos del contenido de humedad para el hígado que es de 72.83%.
Sistema digestivo
Que anatómicamente se refiere a los tres compartimientos gástricos que presentan los camélidos sudamericanos. Massi (1987) reporta un 17.8% para el contenido de proteína en el ganado bovino, mientras que para el caso de las llamas se reporta un 21.2% de proteína, siendo un porcentaje mayor en favor de los camélidos. Para el caso de los lípidos el mismo autor, para bovinos indica un 6%, mientras que en el caso de las llamas se reporta una media de 8.34%. En el caso del aporte calórico para vacunos se reporta 122 cal/100 gr mientras que para las llamas se reporta una media de 138.69 cal/100 gr, para otros componente no se tiene referencias, pero los niveles encontrados en llamas señalan un 66.12% de humedad, el contenido de colesterol es de 147,3 mg/100gr.
Intestinos
Conocida comúnmente como tripa que corresponde a los intestinos delgado y grueso. Massi (1987), reporta un valor de 15.8% para el contenido de proteína en el ganado vacunos, mientras el reporte del trabajo para llamas señala 20.72% existiendo una diferencia mayor a favor de las tripas de las llamas en el contenido de proteína. Para el contenido de lípidos el mismo autor y para la misma especie señala un 5%, un similar valor se reporta en el intestino de las llamas de 5.27%. También el mismo autor indica que el aporte calórico de las tripas en ganado vacuno es de 109 cal/100 gr, mientras que en presente trabajo podemos indicar que el aporte calórico es de 365.86 cal/100 gr sustancialmente mayor en favor de las llamas. Para el porcentaje de humedad se reporta 78.30%, el contenido de colesterol, alcanza un nivel bastante alto como es de 286.60 mg/100 gr
Los pulmones
En relación al análisis de los pulmones son escasos los reportes del análisis químico para este órgano, porque es poco requerido para el consumo, en la mayoría de los casos la demanda es por los órganos de animales muy jóvenes. Se reportan los siguientes análisis, el porcentaje de humedad es del 80.13 %, el contenido de lípidos es de 3.44%, las proteínas reportan una media de 19.50 %, el nivel del colesterol es de 274,90 mg/100 gr, así como el aporte calórico es de 95 cal/100 g.
En base a los valores del porcentaje de proteína de la carne de llamas, para enteros y castrados se efectúo un ajuste de la curva mediante una función logarítmica. Tomando en cuenta el valor del coeficiente de determinación de la relación entre el contenido de proteína y la edad, se obtiene que r2 = 0.70 y r2=0.85 para llamas enteros y castrados respectivamente. Valores que indican que un 70 % y 85 % las variaciones del contenido de proteína en la carne es por la variación de la edad de los animales (Calzada, 1982).
Bibliografía
Aldana, L. (1984). Tecnología de la carne y sus productos. Habana, CU. Editorial Pueblo y Educación. p. 23-33. [ Links ]
Associazione Scientifica di Produzione Animale ASPA. 1971. Zootecnia y producción animal. Roma-Italia [ Links ]
Ayala, C 1992. Estudio detallado de la incidencia de sarcocystiosis en el Altiplano de Bolivia. Seminario Internacional de Tres Zoonosis. Oruro-Bolivia [ Links ]
Calzada, B.J. 1982. Métodos estadísticos para la investigación. Lima, PE. Editorial MILAGROS S.A. p. 209-428, 560-592. [ Links ]
Charley, H. (1995). Tecnología de alimentos: procesos químicos y físicos en la preparación de alimentos. México D.F. Limusa S.A.. p. 519-552. [ Links ]
CISA (Coordinadora Inter-Institucional del Sector Alpaquero, PE); PRORECA (Programa Regional de Camélidos, BO). (1996). Propuestas tecnológicas para la producción y comercialización de carne de camélidos domésticos andinos. Arequipa, PE. p. 125-140. [ Links ]
Condori, G. (2000). Determinación de la edad óptima de faeneo y evaluación de la calidad de la carne de llama. Tesis de grado. Facultad de Agronomía - UMSA, La Paz Bolivia. [ Links ]
Condori, G., Ayala C., Renieri, C., Rodríguez, T. y Martínez, Z. (2003). Evaluación de las características físicas de la carne de llama en diferentes periodos de crecimiento. En Memorias del II Congreso Mundial de Camélidos Potosí- Bolivia. Primer Taller Internacional del Proyecto DECAMA Tomo II. Potosí Bolivia. [ Links ]
Di Marco, O. (1993). Crecimiento y respuesta animal. 1 ed. Buenos Aires, AR. AAPA. p. 11 -30. [ Links ]
Garnica, P. J. (1993a). Composición Química de la Carne. In Bustinza, V. ed. Carne de Alpaca. Puno, PE. EPG-UNA. p. 105-120. [ Links ]
_____. 1993b. Concentración de Colesterol en la Carne de Alpaca. In Bustinza, V. ed. Carne de Alpaca. Puno, PE. EPG-UNA. p. 129-133.
Gerken, M.; Rodríguez, T.; González, M.; Torres, Z.; Ayala, C.; Barreda, E.; Condori, G.: (2000). Evaluación de la calidad organoléptica de carne de camélidos, resultados de panel test en Perú y Bolivia. Simposium Internacional Camélidos Sudamericanos Domésticos. Seminario Final Proyecto Supreme, Arequipa - Perú. [ Links ]
Gerken, M. (1998). Metodología de estudio y análisis de la calidad de carne de camélidos y análisis sensorial. In Seminario Internacional de Camélidos Sudamericanos Domésticos (3), Seminario Proyecto SUPREME (1). Resúmenes. Córdoba, AR. UCCOR, PLANCAD, ENEA, CCE. p. 7-9. [ Links ]
González, M. (1996). Experiencia de DESCO en la crianza de camélidos en Caylloma: perspectivas del desarrollo pecuario en Caylloma. In Crespi D. Marquina, R. eds. Construyendo Nuestro Futuro. Lima, PE. ERPB, USAID, FGCPC, AECI. p. 118-244. [ Links ]
Lieven, C. (1985). Aprovechamiento de la carne de llamas, alpacas y capibaras en regiones tropicales. Tesis de Ph. D. Alemania. Universidad Ludwing-Maximilian de Munich Facultad de Veterinaria. p. 34-38. [ Links ]
Limonta, L. M. (1987). Análisis de la carne y sus productos. Habana, CU. Editorial Pueblo y Educación. p. 30 [ Links ]
Martínez, F. J; Alzerreca, H. (1983). Evaluación del rendimiento en carcasa de alpacas (Lama pacos) en la Estación Experimental de Ulla Ulla. In Reunión Nacional de Pasto y Forrajes (7), Reunión Nacional de Ganadería (5). Potosí, BO. IBTA, INFOL, BCB, ABOPA. p. 45-49. [ Links ]
Mora, E; Ureña, J; Chávez, J. (1994). Utilización de técnicas básicas para el procesamiento de productoscárnicos. Costa Rica. CITA Programa Cooperativo Universidad Costa Rica, Ministerio de Agricultura y Ganadería. p. 98-105 [ Links ]
Penny, I.F. (1984). Enzimología de la Maduración. In Lawrie, R. ed. Avances de la Ciencia de la Carne. Zaragoza ES. Editorial Acribia S.A. p. 149-177. [ Links ]
Pinto, V. M. (1975). Estudio de algunos caracteres de la producción de carne en llamas. Tesis. Lic. Agr. Cochabamba, BO. Universidad Mayor de San Simón. [ Links ]
Rodríguez, T. (1983). Importancia de la influencia de factores ambientales sobre algunos caracteres de producción de carne y lana en llamas (Lama glama). In Reunión Nacional de Pasto y Forrajes (7), Reunión Nacional de Ganadería (5). Potosí, BO. IBTA, INFOL, BCB y ABOPA. p. 81. [ Links ]
Rodríguez, T. (1995). Importancia de la producción de carne de camélidos en la región andina de Bolivia. In I Mesa Redonda Sobre la Comercialización y el Consumo de la Carne de llama. memorias. s.l., IICA, PRORECAp. 11-25. [ Links ]
Téllez, J.G. (1992). Tecnología e industrias cárnicas. Lima PE. Tomo II. p. 346-347. [ Links ]
Tichit, M. (1991). Los Camélidos en Bolivia. La Paz BO. Editorial EDOBOL. p. 16-20. [ Links ]
Torres, S. D. (1998). Análisis de la potencialidad de producción y caracterización cualitativa de la carne de camélidos. In Seminario Internacional de Camélidos Sudamericanos Domésticos (3), Seminario Proyecto SUPREME. (1): resúmenes. Córdoba, AR. UCCOR, PLANCAD, ENEA, CCE. p. 12-13. [ Links ]
Varnam, H. A; Sutherland, P. J. (1995). Carne y productos cárnicos: tecnología química y microbiología. Zaragoza ES. Editorial Acribia S.A. p. 5-57, 71-95. [ Links ]
Vilca, L. M. (1991). Producción, tecnología e higiene de la carne. In Fernández, S. B. ed. Avances y Perspectivas del Conocimiento de los Camélidos Sudamericanos. Santiago Chile. p. 387- 411. [ Links ]
Visier, A. (1980). Industria de la carne. Barcelona ES. Editorial AEDOS. p. 23-36. [ Links ]

