










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
 Citado por SciELO
Citado por SciELO Links relacionados
 Similares en
SciELO
Similares en
SciELO Compartir

 Permalink
PermalinkRevista de Investigación e Innovación Agropecuaria y de Recursos Naturales
versión impresa ISSN 2409-1618
RIIARn vol.5 no.Especial La Paz 2018
ARTÍCULOS
Estudio y caracterización de la aptitud de producción de carne en llamas (Lama glama)
Condori G.12; Renieri C.13; Ayala C.14; Rodríguez T.15; Martínez Z.15
12 Investigador del Proyecto DECAMA, Facultad de Agronomía, Universidad Mayor de San Andrés, Bolivia.13 Docente, Universidad de Camerino, Italia.
14 Docentes Investigador, Facultad de Agronomía, Universidad Mayor de San Andrés, Bolivia.
15 Docente, Facultad de Agronomía, Universidad Mayor de San Andrés, Bolivia.
Introducción
La adaptabilidad de la llama a condiciones ambientales adversas con extrema variación climática hace de esta especie un recurso natural de importancia para los habitantes de la región Andina. Se estima que 57.000 familias están involucrados en la cría de más de dos millones de cabezas asentadas en territorio boliviano (FIDA, 1994). El producto principal que permite parte de los ingresos de los pequeños productores es la carne y en menor proporción la fibra. Se estima que en Bolivia anualmente se producen más de 4.800 TM de carne de llama (Rodríguez, 1991). No obstante la excelente calidad nutritiva de la carne proveniente de esta especie, los precios que se pagan por este producto son bajos y su consumo se limita prácticamente a los habitantes del área rural.
Los estudios relacionados a la evaluación de las características de la carne de llama, orientados a mejorar la calidad del producto final y, consecuentemente mejorar su precio y los ingresos de los pequeños productores, son reducidos. Por lo señalado anteriormente, es necesario obtener información precisa sobre la aptitud de la llama en la producción de carne y la edad óptima para faena y obtener un producto de alta calidad y con el mayor rendimiento; información básica que se pretende obtener con la ejecución del presente trabajo que tiene por objetivos:
- Evaluar la calidad nutricional de la carne de llama (Lama glama, Linnaeus) en diferentes periodos de crecimiento.
- Establecer la edad óptima de faena.
- Determinar las características físicas de la carne de llama.
- Evaluar el desarrollo alométrico de llamas machos.
Metodología
El trabajo se realizó en la Estación Experimental de Choquenaira, dependiente de la Facultad de Agronomía -UMSA, ubicada a 35 km. de la ciudad de La Paz, a una altura de 3750 m.s.n.m., con una precipitación anual entre 400 a 600 mm. Concentrada en tres meses del año, una temperatura promedio de 14°C, la temperatura mínima registrada en el mes de junio alcanza a -3°C y la máxima durante los meses de enero y febrero alcanza a 20 °C. Geográficamente se encuentra situada a 16° 40' de latitud sur y 68° 10' longitud oeste.
Para el estudio se utilizan 50 llamas machos de una misma edad, nacidos entre la tercera semana de enero y la primera semana de febrero de 1997, los animales pertenecen al tipo "intermedio" y fueron colectados en la región de Turco del Departamento de Oruro. Los animales desde su arribo a la Estación Experimental de Choquenaira, se mantienen por las noches en un corral dormidero y la alimentación diaria se realiza bajo el sistema de pastoreo común en praderas nativas del tipo pajonal con predominancia de Stipa ichu y Festuca dolicophylla. El destete se realizó a los 11 meses de edad y a los 12 meses se efectúo el castrado del 42% de los animales; posteriormente se formaron 7 grupos de 7 animales (4 enteros y 3 castrados). La matanza seriada de animales de cada subgrupo se efectúa cada 3 meses a partir de los 13 meses de edad.
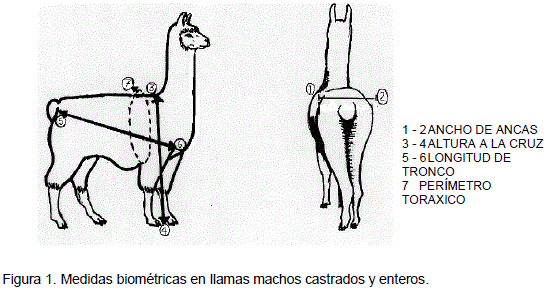
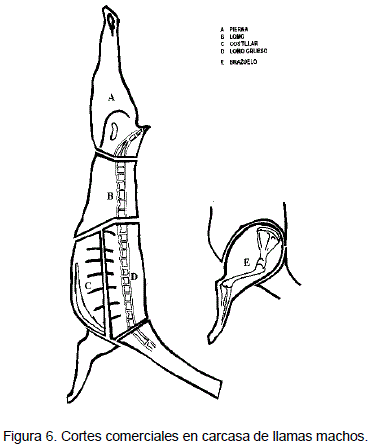
El control de peso corporal se efectuó con una balanza de 100 kg de capacidad y precisión de 250 g. Las medidas biométricas se realizan de acuerdo a la metodología empleada para ovinos por ASPA (Asociación Científica de Producción Animal, 1991) y son: a) Altura a la cruz: se mide con el bastón de Lydken en un plano horizontal al punto más alto del tronco, b) Longitud de tronco: se mide desde la articulación escapo-humeral a la tuberosidad del isquion, en forma diagonal, c) Ancho de Ancas: se mide con el bastón de Lydken entre las articulaciones coxo-femorales y d) Perímetro torácico: se mide con una cinta métrica alrededor del tórax detrás de la espalda al mismo nivel de la altura a la cruz (Figura 1). Tanto el control de peso como las medidas biométricas se realizan cada 31 días.
Antes del faenamiento se realizan los siguientes controles de peso: 1) Control de peso corporal antes de someter a ayuno alimentario no hídrico, 2) Control de peso corporal después del ayuno de 24 horas. El Faenamiento se realizó de acuerdo a la metodología seguida para bovinos y comprende los siguientes pasos: a) Aturdimiento, b) Colgado, c) Degüello, d) Desangrado, e) Desollé y f) Eviscerado.
Posterior al sacrificio se efectúa el control de peso de la carcasa en caliente y 24 horas. después el peso de carcasa en frío, en ambos casos a temperatura ambiente. A continuación se realiza el pesaje de la cabeza (incluyendo la lengua), piel más fibra y, patas; aparato digestivo, que comprende: esófago, estómago, intestinos delgado y grueso, con el contenido alimenticio; posteriormente el contenido alimenticio es vaciado y pesado para determinar el peso vivo neto (PVN); finalmente, se pesan diferentes órganos como ser: corazón, hígado y pulmón. Todos los pesos se efectúan en una balanza de 10 kg de capacidad y 5 g de precisión.
Después que termina la rigidez cadavérica se procede a la división de la carcasa en dos mitades; en el lado izquierdo se practica los cortes comerciales y el derecho está destinado a otras medidas no contempladas en el presente estudio.
En el corte izquierdo se realiza las siguientes mediciones: 1) Largo de carcasa: se mide con un fluxómetro, desde el margen craneal de la sínfisis púbica al punto medio del margen craneal de la primera costilla, 2) Profundidad de tórax: se mide desde el margen dorsal de la 5ta. Vértebra torácica en el punto de la articulación con la 6ta. Al punto medio del margen ventral de la penúltima esternebra, 3) Longitud de Pierna: se mide desde el margen craneal de la sínfisis púbica al tobillo medio (Figura 2).
Posterior a las mediciones se realizan los siguientes cortes comerciales y sus correspondientes pesos.
a) Pierna: Comprende un corte de la región anatómica del muslo y la pierna, tiene como base ósea el sacro pelvis (íleon, isquion, pubis), fémur, rotula tibia y, tarso; el corte será separado del lomo al nivel de la articulación vértebra lumbar y la primera vértebra sacra (Figura 3).
b) Lomo fino: Es un corte que tiene como base ósea las siete vértebras lumbares y como base muscular el largo dorsal; iliopsoas y psoas menor. Este corte se practica entre la última vértebra torácica y la primera vértebra lumbar (entre las articulaciones lumbo-sacral), este corte incluye los músculos abdominales. (Figura 3).
c) Costillar: Este corte se obtiene desde el inicio de la punta anterior del esternón continua por la tercera parte inferior de las costillas y termina al otro extremo en la primera costilla falsa; comprende el esternón el resto de las costillas y los músculos anexos (Figura 3).
d) Lomo grueso: Comprende las doce vértebras torácicas, desde la primera hasta la décima segunda vértebra torácica, tiene su base muscular en el gran dorsal y el largo dorsal y su base ósea en 12 vértebras torácicas y unas dos terceras partes de todas las costillas. (Figura 3).
e) Brazuelo: Comprende la región anatómica de la espalda, brazo y antebrazo, como base ósea tiene a la escápula, humero, radio y carpo; el corte se realiza siguiendo el margen craneal y caudal de la escápula y de
su cartílago. Tiene su base muscular en el trapecio cervical y torácico, homotransverso, tríceps, y deltoides. (Figura 3).
Resultados
Medidas biométricas
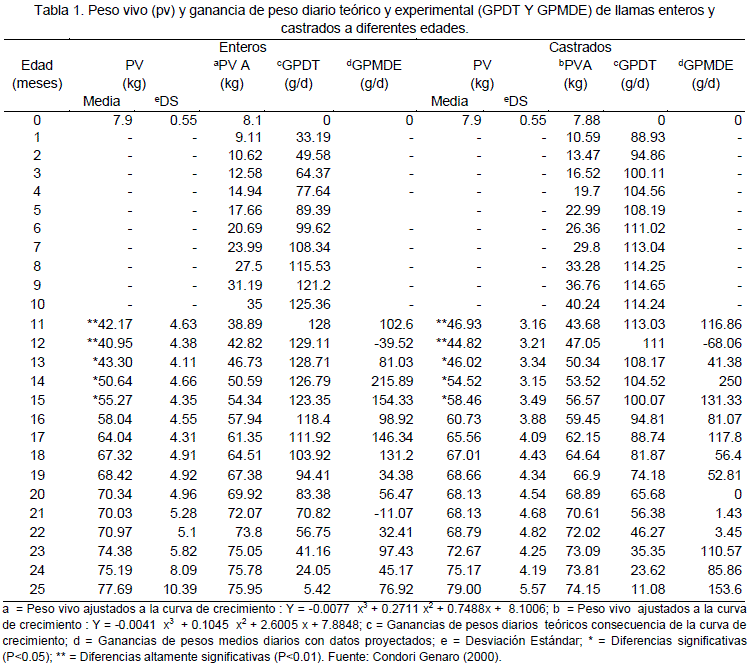
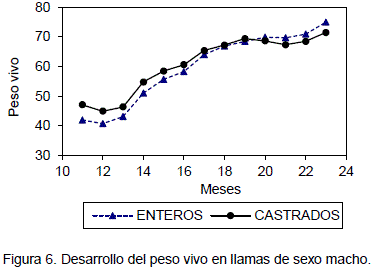
A partir de los 12 meses de edad y hasta los 25 meses de edad, los animales castrados registraron pesos vivos superiores a los animales enteros; después de los 18 meses de edad y hasta los 23 meses, los animales enteros, registraron mayores pesos que los castrados. El peso vivo de los animales castrados a los 12 meses de edad fue de 44.88 kg y a los 23 meses de edad fue 71.4 kg con un incremento de 26.52 kg en 12 meses. El peso vivo en animales enteros de 12 meses de edad fue 40.67 kg y a los 23 meses de edad fue 74.92 kg con un incremento 34.25 kg (Figura 4).
Las medidas de perímetro torácico, largo de tronco, ancho de ancas y altura a la cruz son presentados en la Tabla 1, el incremento en perímetro torácico en un periodo de 11 meses en enteros y castrados fue 15.78 y 14.02 cm en la longitud de tronco fue 13.04 y 9.55 cm, en el carácter ancho de ancas hubo un incremento de 4.98 y 4.66 cm y el incremento para el carácter de altura a la cruz en fue 12.7 y 11.12 cm respectivamente.
Al octavo mes logran un promedio de 37.4 kg, de este al noveno los aumentos de peso son mínimos llegando a 39.45 kg, de ahí al décimo mes existe un crecimiento considerable donde logran un peso de 42.5 kg mientras que para el décimo primer y décimo segundo mes existen incrementos significativos logrando pesos de 45.8 y 48.9 kg, donde estos pesos son mucho mayores que a los del anterior mes, pudiendo deberse esto a factores ambientales como es la alimentación, porque en esos meses comenzó el período húmedo, de ahí adelante llegando al décimo octavo mes los animales logran un promedio de peso de 61.2 kg, en el cual se denota un mediano aumento en el peso, para luego llegar al vigésimo primer mes de edad con 64.9 kg de peso vivo como promedio general. Es importante recalcar que durante los períodos de medición, las llamas q'aras superan en peso vivo a las T'hampullis, sobre todo las llamas q'aras (hembras) que partir del sexto al vigésimo primer mes, muestran un mayor peso vivo en comparación con las llamas q'aras machos y llamas T'hampullis.
Condori et al., (2000), halla con datos de peso vivo desde el nacimiento hasta los 25 meses de edad (Tabla 1) dos ecuaciones por el método de regresión no lineal o función polinomial. La misma (ecuación 1 y 2) para llamas machos sin castrar y llamas castrados respectivamente. Las curvas ajustadas responden a una ecuación de tercer grado en el cual el coeficiente de regresión de la variable "X" elevada al cubo es diferente de cero (d≠0; P<0.01).
![]()
La edad está representada por la variable independiente (X) en meses y el peso vivo como variable dependiente (Y) en kg. En animales de corta edad el término de la ecuación 0.7488 X, en enteros y 2,6005 X en castrados influye en forma preponderante sobre el peso vivo, siendo que esta es imperceptible a la influencia del termino -0.0077 X3 en enteros y -0.0041 X3 en castrados. A medida que aumenta la edad del animal la influencia de este término va aumentando comparativamente más que el anterior y este por ser de signo negativo su efecto tiende a ser contrarrestado (Calzada, 1982).
Según Webster (1989) citado por Di Marco (1993), la curva de crecimiento es una curva sigmoidea y no una recta. Thonney et al., (1981) define que la tasa de ganancia de peso de un animal disminuye cuando aumenta el peso. Asimismo León et al. (1989) ajustan la curva de crecimiento a una ecuación cuadrática (Ecuación 3) y se observa que la tendencia descrita por la misma es diferente a la ecuación 1 y 2 que se muestra anteriormente.
Ecuación 3: Y=-0.30x2 + 10.06x+14.28
Donde Y es el peso vivo estimado y X es la edad en meses. Las ecuaciones cuadráticas de segundo y tercer grado utilizadas como estimadores del peso vivo en función de la edad, son el resultado de la correlación entre las dos variables (León et al. 1989). Si se desea conocer el peso al nacer con cualquiera de estas ecuaciones se reemplaza en X el valor "0" y se tiene los siguientes valores de acuerdo a:
Ecuación 1 Y=8.10kg
Ecuación 2 Y= 7.88 kg
Ecuación 3 Y= 14.28 kg
Ayala (1992) y Rodríguez (1991) reportan una media de 8.4 kg y 9.6 kg respectivamente para peso al nacer en llamas de la Estación Experimental Patacamaya. Estos valores son similares a los resultados de la ecuación 1 y 2 pero diferentes a la ecuación 3, se atribuye esta diferencia a la influencia del medio ambiente puesto que el experimento efectuado por León et al., (1989) fue realizado en el valle de Oregon de los Estados Unidos. Esto nos indica el efecto del medio ambiente sobre la evolución de la curva de crecimiento, especialmente debido a la alimentación.
Curva de crecimiento y ganancia de peso en llamas
Con las ecuaciones ajustadas 1 y 2 para la curva de crecimiento para llamas no castrados y castrados respectivamente, Condori (2000) determinó la ganancia de peso diario para cada mes, (Tabla 2). La curva de desarrollo del peso vivo y la curva de ganancia de peso diario teórico para llamas enteras, en un periodo de crecimiento de 25 meses se observa en la Figura 3.
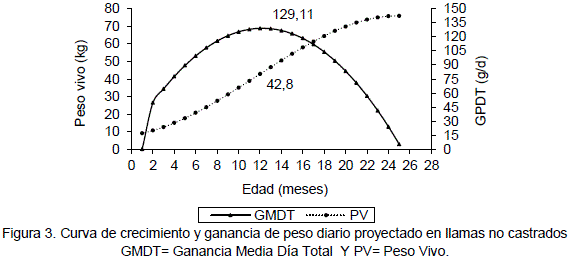
La máxima ganancia de peso diario en llamas no castrados es 129.11 g/d, a la edad de 12 meses. El valor del peso vivo ajustado de 42.82 significa un 36% para llamas machos adultos de 116 kg (Sumar, 1988) citado por Bonavia (1996) y 38% para 110 kg de peso en llamas adultos (Rodríguez, 1981). Estos valores porcentuales son similares al establecido por Lloyd (1982) quien define a la inflexión de la curva como el punto de máxima ganancia de peso donde el animal alcanza el 30% del peso adulto. Posterior a este punto de inflexión se pasa al denominado crecimiento lento.
En la Figura 4 se observa la curva de crecimiento respecto al peso vivo y la curva de la ganancia de peso diario teórico en llamas castrados. En el cual Condori (2000) se determinó la máxima ganancia de peso en 116 g/d a la edad de 9 meses con un peso vivo ajustado de 36.72 kg. El peso vivo corresponde al 31 y 33% del peso adulto de acuerdo a Sumar (1988) citado por Bonavia (1996) y Rodríguez (1983) respectivamente. Este valor es consistente con el señalado por Lloyd (1982). Según Di Marco (1993) citado por Condori (2000), indica en mamíferos en general, los animales logran la máxima eficiencia de crecimiento cuando alcanza el 25% del peso adulto. A partir del cual el incremento del peso se debe a una mayor acumulación de grasa.
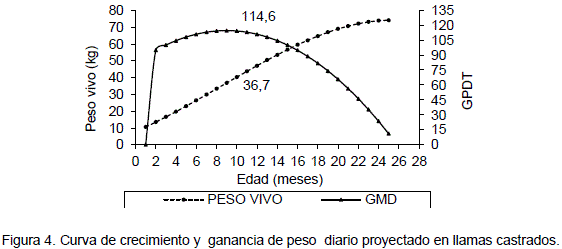
En la mayoría de las especies la máxima ganancia de peso diario, coincide con el inicio de la pubertad Lloyd (1982). De acuerdo a esta afirmación se observó que llamas enteros alcanzan la pubertad a 12 meses de edad, con una ganancia de peso diario de 129 g/d. Este resultado es similar al señalado por Losno y Coyotuba (1981) citados por Sumar (1991) que definen el inicio de la pubertad en alpacas a partir de los 11 meses.
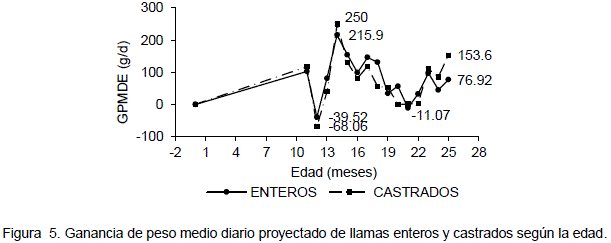
Según Condori (2000) las llamas alcanza la máxima ganancia de peso diario a la edad de 14 meses con una media de 215.8 g/d, con un peso vivo de 50.54 Kg para no castrados y 250 g/d, con 53.52 kg para castrados (Tabla 1). En la Figura 5 se observa los datos de ganancia de peso diario los cuales muestran pendientes bruscas sin embargo es notable el pico máximo que alcanzan a los 14 meses tanto para enteros como para
castrados y una notoria ganancia de peso negativa a los 12 y 21 meses atribuible a las condiciones ambientales.
La máxima ganancia de peso medio diario experimental y el peso vivo alcanzado a los 14 meses de edad para no castrados y castrados son similares a los mencionados por CISA (1996) quienes señalan una ganancia de peso diario de 233 g/d y 51.57 kg. Para llamas jóvenes de 10 meses de edad en etapa de crecimiento y engorde. Sin embargo la edad a la que alcanzan la máxima ganancia de peso diario en ambos experimentos son diferentes, es atribuible esta diferencia al tipo de tratamiento recibido y el medio ambiente.
Factores que afectan el crecimiento y desarrollo
Esta considera que el grado de crecimiento y desarrollo definidos para la edad adulta de cada especie, está sujeto a la herencia, variabilidad individual y nutrición e implica que debe producirse un crecimiento y desarrollo completo y coordinado de todas y cada una de sus partes, fenómenos que requieren un gran número de procesos.

La diferencia en la ganancia diaria de peso está determinada por potencial genético distinto. Comparativamente los machos crecen más rápido que las hembras, debidos precisamente a la mayor potencial de los andrógenos con respecto a los estrógenos. Los machos consumen más alimento que las hembras por una mayor tasa metabólica, pesan al nacer entre el 5 y 7 % más que las hembras, el largo de la gestación es de 3-4 días más en el macho, son más eficientes en la conversión de alimento que las hembras (Varman y Sutherland, 1995).
Efecto de la castración en el crecimiento del ganado de carne
En cuanto al efecto de la castración en machos, se debe tener en cuenta que los andrógenos tienen un efecto miotrófico, es decir favorecen el desarrollo de determinadas áreas corporales y especialmente ciertas masas musculares. A una misma edad, el novillo tiene una mayor altura que un toro de la misma raza, porque se ha eliminado el efecto antagónico de los andrógenos con respecto a la hormona de crecimiento. Los machos enteros son más eficientes para ganar peso que los castrados, dado que las relaciones músculo/hueso y músculo/grasa son siempre favorables a los machos enteros (Prescot, 1982).
Inchausti y Ezequiel (1980) mencionados porCondori (2000) indican que un animal castrado carece de ciertas secreciones internas lo cual mejora la calidad de la carne. En este aspecto, interviene la mansedumbre consecuencia de la inactividad testicular lo cual causa menor ejercicio, consecuentemente las fibras musculares son más blandas y la carne más tierna.
La castración de becerros es para mejorar la textura, suavidad y sabor de la carne. Sin embargo, los machos no castrados presentan un mayor rendimiento de carne magra y menor cantidad de grasa en comparación a los castrados, en un período de 12 a 18 meses de edad (Neuman, 1989 citado por Condori, 2000). En llamas se encontró que la castración tiene gran influencia la ganancia de peso presentando diferencias altamente significativas (P<0.01) entre llamas no castrados y castrados, a partir de los 13 meses hasta los 15 meses de edad (Tabla 1) Condori (2000). Sin embargo en el estudio se encontró que a la edad de beneficiado (24 meses) la castración no tiene un efecto significativo en la diferenciación en el peso en comparación a los no castrados.
Efecto del control hormonal en el crecimiento
Las hormonas anabólicas (que favorecen el crecimiento) son: La somatotrofina, hormona que regula el crecimiento del hueso y músculo. Potencia el incremento la producción de carne y leche. La insulina es la hormona que regula la unión de otras hormonas con sus receptores, actúa a nivel de los receptores hepáticos de la ST. Los andrógenos, regula el crecimiento en ambos sexos. La testosterona, es el andrógeno primario secretado por los testículos y las glándulas adrenales en hembras. La secreción aumenta antes de la pubertad y es responsable del rápido crecimiento en esta etapa. Los machos crecen más rápido que las hembras, debido a estos factores. Los estrógenos secretados por los ovarios, desarrollan el tracto reproductivo en todas las especies, también incrementan el desarrollo muscular en rumiantes. Los glucocorticoides secretado por glándulas adrenales, causan remoción de nutrientes de las reservas corporales para producir energía lo que resultan en pérdida de peso, lo que no ocurre en animales sanos.

Al examen de la Tabla 1 se reporta el coeficiente de correlación entre medidas biométricas, en la canal, como son peso de carcasa, peso de cortes comerciales y peso vivo, el cual nos permite expresar algunas consideraciones. El valor de la correlación entre perímetro torácico y peso vivo (r =0.94), entre peso vivo y carcasa en caliente (r=0.97), entre peso vivo y carcasa en frío (r = 0.97) es superior al reportado por Pinto (1975) r= 0.82, r =0.44 y r = 0.49 respectivamente, la correlación entre longitud de tronco y canal frío r = 0.78 es similar al reportado por Pinto(1975) con r = 0.705, el coeficiente de correlación entre longitud de tronco y largo de carcasa r=0.9 es superior al reportado por Pinto (1975) con r=0.742.
Medidas en carcasa
Las medidas de longitud de carcasa, profundidad tóraxy largo de pierna, en animales enteros y castrados entre la primera y la cuarta faena son presentados en la Tabla 3. El incremento en longitud de carcasa durante el periodo de 9 meses fue de 9 cm, en profundidad de tórax fue de 4 cm y en largo de pierna 7 cm.


Los pesos de cortes comerciales (peso de lomo grueso, lomo fino, pierna, brazuelo y cuello) presentan un coeficiente de correlación alto a muy alto con las medidas biométricas con excepción de las variables altura a la cruz y peso de lomo grueso que tienen una correlación media de 0.51. El peso del corte costillar presenta un coeficiente de correlación muy bajo con respecto a las medidas biométricas.
Las medidas en carcasa (longitud de carcasa, perímetro torácico, largo de cuello, y longitud de pierna), manifiestan coeficientes de correlación alto a muy alto con las variables pesos de lomo grueso, pierna, brazuelo y cuello, las variables largo de cuello y peso de lomo fino tienen un coeficiente de correlación baja, r =0.5. El corte costillar presenta un coeficiente de correlación muy bajo con las medidas de carcasa y un coeficiente de correlación muy bajo y negativo con la medida largo de cuello.
Los pesos de carcasa en caliente y en frío presentan coeficientes de correlación muy alta con los pesos de cortes comerciales con excepción del corte costillar que presenta un coeficiente de correlación bajo 0.44 y 0.42 para peso de carcasa en caliente y frío respectivamente.
Rendimiento de la canal
Los resultados obtenidos para rendimiento de carcasa 57.46 y 53.46% en canal caliente y en frío respectivamente, son superiores a los encontrados por Pinto (1975), 52.12 y 46.64% para carcasa en caliente y en frío (24 horas después del sacrificio) respectivamente, Sin embargo son similares a los reportados por Bravo (1981) 58.1 y 59.6% en animales enteros y castrados de un año de edad, respectivamente. Los promedios de los rendimientos de carcasa en caliente y en frío se aprecian en la Tabla 6.
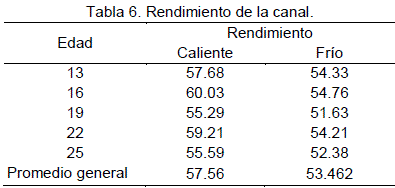
Se evidencio que existe una merma en el rendimiento después de 24 horas de 4.1% este resultado es inferior al reportado por Pinto (1975) 5.48%.
Cortes comerciales
En la Tabla 7 se reporta los valores de los coeficientes alométricos de los pesos de cortes comerciales, con el peso de media canal izquierda, que ha sido fijado como variable independiente; el coeficiente alométrico "b" fue calculado en base a la evaluación de 5 faenas a diferentes edades. En animales enteros el cuello presenta el mayor valor, lo que significa que tiene una mayor velocidad de crecimiento y madurez tardía; mientras que el costillar presenta el valor más bajo. En castrados, el mayor coeficiente alométrico es registrado en brazuelo, en tanto que el costillar, al igual que en enteros, presenta el valor más bajo.
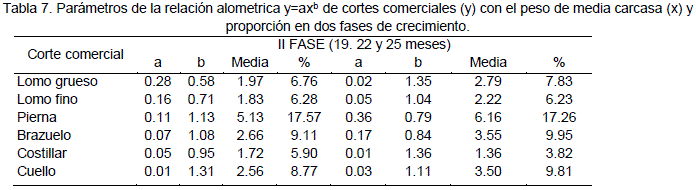
La pierna y el brazuelo presentaron un desarrollo contrario a los anteriores cortes, en la fase 1 mostraron un desarrollo tardío sin embargo en la fase 2 presentaron un desarrollo precoz, entre los dos cortes la pierna presenta mayor precocidad.
El cuello presentó en ambas fases un desarrollo tardío con respecto a otros cortes. De modo general la tendencia de crecimiento de los diferentes cortes en la fase 1, demuestra que existe pleno desarrollo en los miembros anteriores y posteriores propio de animales jóvenes en la fase 2 el crecimiento es mayor en la parte de lomos y menor en los miembros anterior y posterior
En la práctica a medida que aumenta el peso por lo tanto avanza la edad hacia 25 meses el peso de cortes tiende a distribuirse en mayor medida en los lomos y cuello respecto a los otros cortes esta aseveración es confirmado por Geri et al. (1984) quien establece un crecimiento centrípeto para suinosde20 a 200 Kg, los
valores de la segunda fase aquí presentados son corroborados por Giorgetti et al. (1987) en ciervos de Italia cuyos valores son 1.17, 1.15, 0.95 y 0.92 para las partes anatómicas de: cuello, lomo, brazuelo y pierna respectivamente. Entonces queda confirmado que la pierna y brazuelo como corte de interés comercial debido a la abundancia en masa muscular y por la mayor proporción que refleja con respecto a la carcasa (Tabla 5), tiene un crecimiento precoz en este periodo tanto en llamas como en ciervos, el cuello al igual que el lomo grueso tienen una velocidad de crecimiento tardío en ambas especies.
Rendimiento proporcional de carcasa y órganos con respecto al peso vivo neto
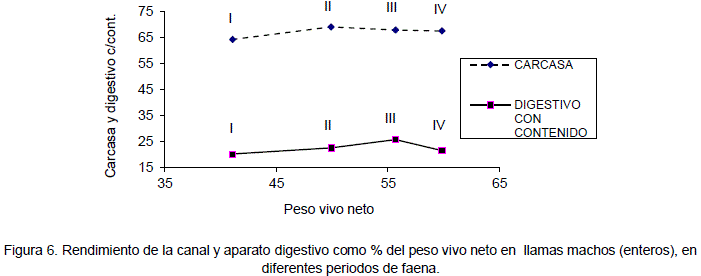
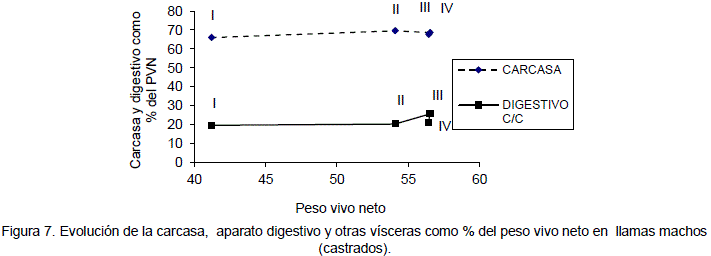
Del rendimiento de la carcasa en animales enteros con respecto al a peso vivo neto tiene un promedio de 64 a 70%. Los rendimientos en relación al peso corporal bruto varían en un rango de 55.6 a 59.1% en llamas macho enteros y 54.8 a 60.9% en llamas de sexo macho castrados, estos rendimientos son superiores a los obtenidos por Pinto et al., 1975 (52.1%) en llamas adultas de sexo macho; sin embargo, son similares a los reportados por Bravo et.al. en 1981 (58.1 y 59.6%) para machos enteros y castrados de 1 año de edad respectivamente. Las proporciones de los otros órganos y partes corporales cuadros son similares a los reportados por Bustinza et al. (1986). La evolución del rendimiento de la carcasa y aparato digestivo con contenido en relación al peso vivo neto se observa en las Figuras 6 y 7.
Bibliografía
ASPA (Associazione Scientifica di Produzione Animale). 1989. Metodiche per La Determinazione Associazione delle Caratteristiche Qualitative della carne. Universita degli Studi di Perugia. [ Links ]
ASPA (Associazione Scientifica di Produzione Animale). 1991. Metodología Relative alla Macellazione degli Animali de Interesse Zootecnico e alla Valutazione e Dissezione della loro Carcasssa. Ismea- Roma. [ Links ]
Ayala, C, 1992. "Crecimiento en peso vivo y en largo de mecha en llamas de la estación experimental Patacamaya Bolivia", Tesis de Grado, Universidad Nacional del Altiplano. Puno, Perú. [ Links ]
Bravo, W.; Franco, E.; Sumor, J. 1981. Rendimiento de canal en llamas. Res. IV Convención Internacional sobre Camélidos Sudamericanos. Punta Arenas, Chile. [ Links ]
Bustinza, V.; Mamani, G. Y Ccoyoto, C. 1986. Peso vivo y rendimiento de canal en llamas.
Res. IX Reunión Científica Anual APP. Tingo María, Perú.
Bonavia, D. 1996. Los camélidos sudamericanos: una introducción a su estudio. IFEA-UPCH, Conservation International. p. 65-68, 496-499. [ Links ]
Clavo, N. y Belon, J. 1970. Algunos aspectos químicos de las carnes de alpaca (Lama Pacos) y llama (Lama glama) que se consume en la ciudad de Puno. Res. VI Congreso Panamericano de Medicina Veterinarias y Zootécnia. Santiago, Chile. [ Links ]
Condori, G. 2000. Determinación de la edad óptima de faeneo y evaluación de la calidad de la carne de llama. Tesis de grado. Facultad de Agronomía - UMSA, La Paz, Bolivia. [ Links ]
Condori, G., Ayala C., Renieri, C., Rodríguez, T. y Martinez, Z. 2003. Alometria de cortes comerciales en carcasas de llama en dos fases de crecimiento. En Memorias del II Congreso Mundial de Camélidos Potosí- Bolivia. Primer Taller Internacional del Proyecto DECAMA Tomo II. Potosí, Bolivia. [ Links ]
Di Marco, O. 1993. Crecimiento y respuesta animal. 1 ed. Buenos Aires, AR. AAPA. p. 11-30. [ Links ]
Leon, J.B; Smith, B.B; Tim, K.L. 1989. Growth characteristics of the llama (Lama glama) from birth to 12 month old. In The Veterinary Record. Oregon. p. 644-645. [ Links ]
Lloyd, L.E; Mcdonald, B; Crampton, E.W. 1982. Fundamentos de Nutrición. Zaragoza, ES. Editorial, Acribia, p. 375-379. [ Links ]
FIDA; 1994. Proyecto de Desarrollo de Criadores de Camélidos en el Altiplano Andino, La Paz, Bolivia. [ Links ]
Pinto, M., Iñiguez, L. y Riera, S. 1975. Estudio de algunas características de la producción de carne de llamas. Res. II Convención Internacional sobre Camélidos Sudamericanos. Juliaca, Perú. [ Links ]
Rodríguez, T., 1991 Situación y Perspectivas de la Producción y Conservación de los Camélidos Sudamericanos en Bolivia, Informe de la Mesa Redonda sobre Camélidos Sudamericanos. Instituto Nacional de Investigación Agraria y Agroindustrial del Perú - INIA. Oficina Regional de la FAO para América Latina y el Caribe. Lima, Perú, 24 al 26 de Septiembre. [ Links ]
Sumar, K. J. 1991. Fisiología de la reproducción del macho y manejo productivo. In Fernández, S. B. ed. Avances y Perspectivas del Conocimiento de los Camélidos Sudamericanos. Santiago Chile. FAO. p. 145. [ Links ]
Varnam, H. A., Sutherland, P. J. 1995. Carne y productos cárnicos: tecnología química y microbiología. Zaragoza ES. Editorial Acribia S.A. p. 5-57, 71-95. [ Links ]

