










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
 Citado por SciELO
Citado por SciELO Links relacionados
 Similares en
SciELO
Similares en
SciELO Compartir

 Permalink
PermalinkRevista de Investigación e Innovación Agropecuaria y de Recursos Naturales
versión impresa ISSN 2409-1618
RIIARn vol.1 no.1 La Paz 2014
ARTÍCULOS
Variación fenotípica intra- e inter-poblaciones en siete
poblaciones de quinua del Altiplano Boliviano
Intra and inter population phenotypical variation in seven
communities of the Bolivian Altiplano
Carmen R. Del Castillo Gutiérrez1 y Thierry Winkel2
1 Facultad de Agronomía, Universidad Mayor de San Andrés,
Héroes del Acre N°1850, La Paz, Bolivia.
2 IRD, CEFE-CNRS, F-34293 Montpellier cedex 05, Francia
RESUMEN
La quinua (Chenopodium quinoa Willd.) muestra una gran variación genética, tanto molecular como morfológica, cuya organización sigue poco conocida. Por medio de ANOVAs y dendrograma caracterizamos la estructura de la variación de marcadores morfo-fenológicos registrados desde la emergencia hasta la madurez fisiológica en siete poblaciones de quinuas cultivadas dos años en jardín común. Los resultados muestran que la varianza morfo-fenológica se concentra en niveles de población y familia, cuando los rasgos relacionados con la producción de biomasa muestran alta variación a nivel interindividual. Estos resultados tienen implicancias agroecológicas relacionadas con el origen geográfico de las poblaciones. La comparación de clasificaciones fenotípica y molecular sugiere que no hay erosión genética en las poblaciones de quinua muestreadas.
Palabras clave: Análisis jerárquico, Bolivia, Chenopodium quinoa, población, variación fenotípica
ABSTRACT
Quinoa (Chenopodium quinoa Willd.) shows great genetic variation, both molecular and morphological, which organization remains poorly documented. Through nested ANOVAS and dendrogramme we characterized the structure of the variation in morpho-phenological traits registered from seedling emergence until plant maturity in seven quinoa populations grown in common garden for two years. Results show that the variance in morpho-phenological traits was concentrated at the population and family levels, while traits related to biomass production showed high inter-individual variation. These results have agroecological implications related to the geographic origin of the populations. A comparison of phenotypic and molecular classifications suggests that there is no genetic erosion in the sampled quinoa populations.
Keywords: Hierarchical analysis, Bolivia, Chenopodium quinoa, population, phenotypic variation
INTRODUCCIÓN
La variación genética de las plantas cultivadas contribuye a la estabilidad de la producción agrícola, permitiendo: i) elegir variedades adaptadas a condiciones ambientales, ii) guiar el mejoramiento genético hacia variedades tolerantes a la sequía, la salinidad, o el frío. Pero a nivel de parcela, la variación entre plantas se considera generalmente como desfavorable para la producción. Esta variación intra-población parece, sin embargo, adaptada a las variaciones imprevisibles del clima (e.g. Winkel et al., 2009). Existe entonces una discrepancia respecto al interés agronómico de la variabilidad genética intra-varietal. Aunque preferentemente autógama, la quinua (Chenopodium quinoa Willd.), pseudocereal originaria de los Andes, muestra una notable variación genética inter- e intra-población, fácilmente observable en parcelas campesinas, y cuantificable por marcadores moleculares (e.g. Del Castillo et al., 2007). Cuanto a marcadores morfológicos, los estudios sobre la diversidad genética de la quinua incluyen: Risi y Galway (1989) sobre los patrones de la diversidad genética en los Andes, Bonifacio (1992) sobre la herencia del color de granos y de hojas, Canahua Murillo et al. (2004) sobre las clasificaciones campesinas de cultivares, y Bhargava et al. (2007) sobre los rasgos morfológicos y de calidad de varios Chenopodium recientemente introducidos en la India. Pero ninguno de estos estudios buscó explicar la distribución de la variación genética entre los varios niveles de organización de la especie: poblaciones de varias ecoregiones, familias de una población y, finalmente, individuos dentro de las familias. Los análisis multivariados son herramientas poderosas para desvelar los factores de organización en cada nivel. Sirven también para clasificar variaciones cuantitativas y cualitativas en especies cultivadas y examinar las relaciones entre genotipos (e.g. Rojas et al., 2000; Bhargava et al., 2007).
Los objetivos de este estudio han sido: i) analizar la estructura de la variación de marcadores morfológicos y fenológicos en siete poblaciones de quinua del altiplano boliviano cultivadas en jardín común, ii) comparar la variación fenotípica con la variación de marcadores moleculares, y iii) relacionar la variación fenotípica con el origen geográfico de las poblaciones.
MATERIAL Y MÉTODOS
El estudio se realizó en las gestiones agrícolas 2004-2005 (Año 1) y 2005-2006 (Año 2), en la Escuela Agropecuaria Superior de Viacha de la Facultad de Agronomía de la Universidad Mayor de San Andrés (16° 39' sur, 68° 18' oeste, altitud 3.830 m).
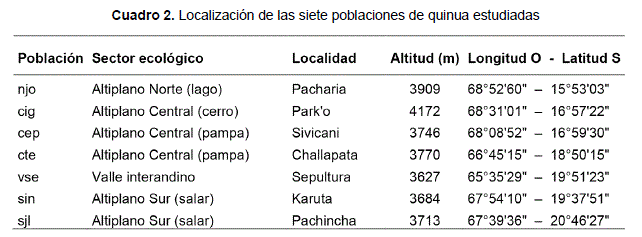
Siete poblaciones, representativas de las zonas agro-ecológicas del cultivo de quinua en Bolivia, han sido muestreadas en parcelas de agricultores el 2002 y 2003 (Cuadro 1). En cada población, 10 plantas-madres han sido recolectadas para representar la diversidad de tamaño y color de las plantas en la parcela. En los ensayos en jardín común descritos más abajo, para cada población, los granos de 10 plantas-madres diferentes han sido sembrados en macetas separadas con 10 repeticiones por planta-madre. Estas 10 repeticiones por planta-madre resultaron así en 10 «plantas-hijas» de una misma familia, y cada población con 10 familias, o sea 100 individuos por población.
La siembra se realizó en macetas el 7 de noviembre 2004 en el primer ensayo y el 30 de octubre 2005 en el segundo. Cada maceta de 11.4 L contenía una sola planta cultivada en una mezcla de turba y tierra franca. Las macetas estaban dispuestas con una densidad de 10 plantas/m2, al aire libre, debajo de una tela anti-granizo. Los datos meteorológicos se registraron en la estación experimental de Choquenaira (Universidad Mayor de San Andrés) distante de 2 km: las temperaturas media, mínima y máxima absolutas fueron de 10.8 °C, 3.0 °C y 19.2 °C el Año 1, y 13.7 °C, 2.5 °C y 24.8 °C el Año 2, con precipitaciones cumuladas en el periodo de ensayo de 311 y 471 mm respectivamente.
Las siete poblaciones de quinua han sido caracterizadas por una serie de rasgos morfo-fenológicos definidos en el Cuadro 2, y medidas en cada individuo separadamente. La biomasa final ha sido medida después de secar en 65 °C durante 72 horas.
Después de un análisis descriptivo convencional, la estructura de la variación fenotípica ha sido examinada por: 1) un análisis de varianza jerárquico («nested ANOVA») con efectos puramente aleatorios separando la varianza en tres niveles: inter-poblaciones, inter-familias dentro de las poblaciones, e intra-familias; 2) una clasificación jerárquica por el método del promedio no ponderado (dendrograma UPGMA). El test de Levene sirvió para verificar la homogeneidad de las varianzas intra-familia dentro de cada población. Un análisis de contingencia permitió comparar esta clasificación UPGMA en base a rasgos morfo-fenológicos con la clasificación UPGMA en base a marcadores RAPD establecida por Del Castillo et al. (2006) para seis de las siete poblaciones estudiadas. Estos análisis han sido realizados con los programas SAS System versión 8, Statistica versión 6.0 y Excel Stat.
RESULTADOS Y DISCUSIÓN
Estadísticas descriptivas de los rasgos morfo-fenológicos
De manera general, los rasgos relacionados con la producción de biomasa o el número de granos estaban variables (CV > 60%) que los relacionados con el desarrollo fenológico de las plantas (CV < 20%) (Cuadro 1). Los rasgos morfológicos de las plantas (HP, DP, NR, P100, IR, IF) presentaron niveles de variación intermediarios, entre 20 y 30%.

Definiciones: JAS: emergencia, número de días desde la siembra hasta la aparición de cotiledones; DEF: inicio de floración = fecha de brote del 50% de las flores de la panoja (número de días después de la siembra) ; DUF: duración de floración = duración desde DEF hasta brote del 80% de las flores de la panoja; MF: madurez fisiológica = fecha en la cual los granos tienen consistencia harinosa (número de días después de la siembra); NR: número de ramificaciones primarias del tallo principal; DP: diámetro de la panoja, medido en la parte media el día de la cosecha; HP: altura de la planta, desde la base del tallo hasta la extremidad de la inflorescencia el día de la cosecha; PP: peso de la panoja; PG: peso de granos; PT: peso total de la parte aérea ; IR: índice de cosecha = PG/ PT; IF: índice de fertilidad = PG/PP; NG: número total de granos por planta, sólo para el Año 2; P100: peso de 100 granos = PG/NG * 100, sólo para el Año dos.
Las medias y máxima observadas muestran que el Año dos fue menos favorable para la producción de biomasa que el Año uno, probablemente a causa del exceso de lluvia al inicio del ciclo. Pero la reducción de biomasa (prácticamente -50%) no conllevó diferencias notables en el desarrollo de las plantas, ni en su morfología (DEF, MF, HP, NR, IR e IF similares entre los dos años).
Estructura general de la variación fenotípica
Según la importancia relativa de la varianza inter-poblaciones, inter-familias e intra-familias, el ANOVA jerárquico para el conjunto de poblaciones permite diferenciar dos clases de rasgos (Figura 1):
- los relacionados con la morfología y la fenología de las plantas (HP, NR, DP, P100, DEF, MF): dominados por la variación inter-poblaciones, estos rasgos son ante todo característicos de las poblaciones mismas, lo que justifica su uso para la clasificación de ecotipos y variedades;
- los relacionados con la producción y el reparto de la biomasa (PP, PG, PT, NG, IR, IF): dominados por la variación intra-familias, estos rasgos dependen más que todo del nivel individual y de las capacidades fisiológicas realizadas en las condiciones particulares del ciclo de cultivo. De hecho, este patrón se fortaleció el Año 2 cuando las condiciones de cultivo se volvieron menos favorables.
Para JAS, la variación inter-individual dominaba, ya que esta variable no es un rasgo fenológico sencillo, sino más bien un proceso (la emergencia) complejo y rápido, fuertemente influenciado por el micro ambiente de las semillas.
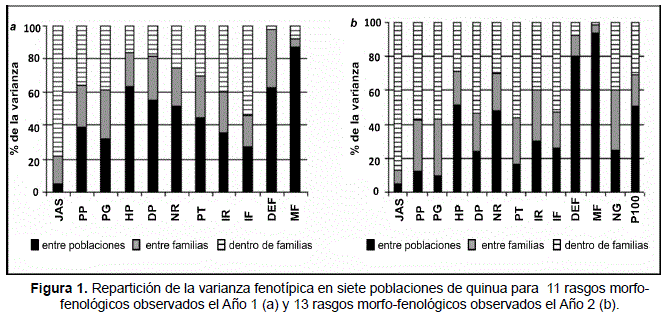
Variación inter-poblaciones, inter- e intra-familias
Una síntesis de los resultados obtenidos con las comparaciones de medias entre poblaciones, así como las ANOVAs sucesivas (Del Castillo, 2008) muestra que:
- las quinuas del sur del altiplano (sin, sjl, cte) fueron precoces, mientras que la del valle inter-andino (vse) fue la más tardía, siendo aquellas del centro y del norte (njo, cig, cep) intermedias.
- las poblaciones menos productivas en biomasa (cte, cig, sjl, sin) tenían rasgos más estables entre los dos años: su pérdida media de biomasa total en el Año dos fue del -39% (-41% para producción de granos) mientras que fue del -54% (-56% en granos) en las poblaciones más productivas (njo, cep, vse). Las poblaciones menos productivas provenían de sitios alejados por la distancia, pero todos ecológicamente limitantes para la agricultura: áridos y fríos para sin, sjl y cte, muy elevados y fríos para cig (Cuadro 2).
- las poblaciones más productivas (njo, vse) tenían los índices de cosecha y de fertilidad (IR, IF) más bajos, lo que sugiere una limitación de la producción de granos por la estructura de las panojas más que por la capacidad fotosintética. La estabilidad interanual de los índices IR e IF traduce la constancia de la relación entre biomasa total y producción de granos, a pesar de posibles variaciones en la producción total. Estos rasgos podrían entonces ser característicos de las poblaciones, pero solamente se podría concluir con ensayos bajo estreses controlados.
- en general, la variabilidad inter-familias superaba la variabilidad intra-familias para el conjunto de las poblaciones, lo que sugiere una herencia elevada para los rasgos considerados. Sin embargo, en el Año 2, la variabilidad intra-familias fue dominante en las poblaciones cep, cte, sin y sjl (Figura 2): la herencia de los rasgos morfo-fenológicos sería entonces menos elevada en estas cuatro poblaciones.

- la homogeneidad de las varianzas intra-familias de una misma población dependía de los rasgos y de los años. En la población cep, por ejemplo, para los dos años y para la mayoría de los rasgos, la varianza intra-familias se diferenciaba significativamente entre las 10 familias. Al contrario, las poblaciones cig y sin mostraron varianzas homogéneas entre sus 10 familias respectivas.
Clasificación jerárquica
El dendrograma, basado en las medias de los rasgos calculadas sobre los dos años en cada familia, diferencia tres grupos principales (Figura 3). Los grupos A y B corresponden a poblaciones del altiplano norte a menos de 4000 m de altura (njo y cep) y del valle inter-andino (vse). Las poblaciones del grupo C se sitúan en el altiplano sur (cte, sin, sjl), o en el norte a más de 4.000 m de altura (cig).
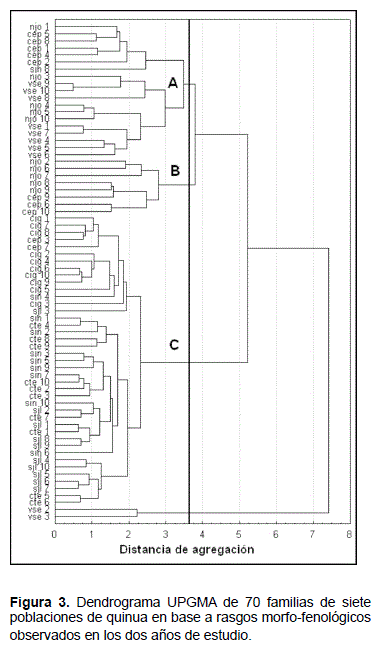
Análisis de contingencia morfología / genética
Para este análisis la variable dependiente fue el vector de tres grupos generados por el análisis molecular (Del Castillo et al., 2007), y la variable independiente fue el vector de tres grupos producidos por la clasificación jerárquica de los rasgos morfo-fenológicos (Figura 3). Este análisis se aplicó sólo en las seis poblaciones comunes a los estudios con marcadores RAPD y marcadores fenotípicos, o sea: njo, cig, cep, vse, sin y sjl. El Cuadro 3 muestra que los grupos moleculares dos y tres corresponden cada uno a fenotipos distintos (A y C, respectivamente).
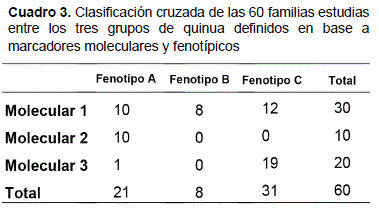
Pero estos mismos fenotipos A y C son genéticamente heterogéneos y agrupan cada uno una decena de familias del grupo molecular uno. El fenotipo B, al contrario, está compuesto sólo por familias del grupo molecular uno. Según Roldán-Ruiz et al. (2001), tal divergencia entre clasificaciones molecular y fenotípica no debe ser considerada como un defecto de los métodos: fenotipos similares no son necesariamente homogéneos en el plan genético, y una correspondencia estrecha entre parentescos molecular y morfológico sería más bien el indicio de erosión genética. Al contrario, la divergencia entre clasificaciones encontrada en el presente estudio sugiere, entonces, que no hay erosión genética notable en las siete poblaciones estudiadas.
CONCLUSIONES
Los rasgos morfo-fenológicos, cuantitativos, y sensibles al medio ambiente permiten evidenciar las relaciones entre limitaciones ambientales, rasgos de vida, y capacidades de adaptación de las poblaciones (Carvajal-Rodríguez et al., 2005). Nuestros resultados muestran una estructura poblacional en estos rasgos, lo que justifica su uso como descriptores de poblaciones. A pesar de discrepancias, las clasificaciones molecular y fenotípica revelan una estructuración geográfica de las poblaciones que, en el caso de los marcadores morfo-fenológicos, separa las quinuas de los sectores más limitantes para la agricultura (altiplano sur y zonas frías del altiplano norte) de aquellas cultivadas en zonas más templadas del lago Titicaca o de los valles inter-andinos. La variación intra-población de estos rasgos también muestra estructura, con una variación inter-familias predominante, lo que sugiere una elevada herencia. Al contrario, rasgos relacionados con la producción de biomasa (panojas y granos en particular), presentan una fuerte variabilidad inter-individual y son sensibles a cambios ambientales. Una elevada variación intra-población en los rasgos cuantitativos indica una presión selectiva poco intensa, posible resultado de una importante varianza ambiental. Este argumento parece válido en el caso del altiplano boliviano, donde el clima tiene un papel mayor en la variabilidad ambiental. De hecho, hemos observado una gran variación fenotípica en las poblaciones cep, njo y vse provenientes de regiones templadas pero sujetas a sequías y heladas eventuales, mientras que las poblaciones cig, sjl, sin y cte de sectores constantemente áridos y fríos mostraron menor variación intra-población. La diferenciación de poblaciones de quinua en el altiplano boliviano estaría así ampliamente determinada por la selección bajo factores ambientales, en particular climáticos, y por la variabilidad local de los mismos.
AGRADECIMIENTOS
Los autores agradecen el apoyo de la Escuela Agropecuaria Superior de Viacha en la gestión del entonces Director Ing. Moisés Quiroga. Este trabajo ha sido co-financiado por la Facultad de Agronomía de la UMSA (Bolivia) y el IRD (Institut de Recherche pour le Développement, Francia). CDC se benefició de una beca doctoral del IRD.
BIBLIOGRAFÍA
Bhargava A., Shukla S., Rajan S.,y Ohri D., 2007. Genetic diversity for morphological and quality traits in quinoa (Chenopodium quinoa Willd.) germplasm. Genetic Resources and Crop Evolution54: 167-173. [ Links ]
Bonifacio A., 1992. Herencia mendeliana en la quinua. In: Actas del VII Congreso Internacional de Cultivos Andinos. Orstom, La Paz, Bolivia. [ Links ]
Canahua Murillo A., Mujica A.S., Apaza V., y Quispe M., 2004. Revaloración de quinuas de color (Chenopodium quinoa Willd.) en el altiplano de Puno, Perú: experiencias y perspectivas. CD-Rom: XI Congreso Internacional de Cultivos Andinos, Fundación PROINPA, Cochabamba, Bolivia, Cochabamba, Bolivia. [ Links ]
Carvajal-Rodríguez A., Rolán-Alvarez E., y Caballero A., 2005. Quantitative variation as a tool for detecting human-induced impacts on genetic diversity. Biological Conservation 124: 1-13. [ Links ]
Del Castillo C., 2008. Diversité génétique et réponse aux contraintes du climat: une étude de cas à partir de la biologie des populations de quinoa (Chenopodium quinoa Willd.) de Bolivie. Tesis de Doctorado. Facultad de Ciencias Agronómicas, Gembloux, Bélgica. [ Links ]
Del Castillo C., Winkel T., Mahy G., y Bizoux J.P., 2007. Genetic structure of quinoa (Chenopodium quinoa Willd.) from the Bolivian Altiplano as revealed by RAPD markers. Genetic Resources and Crop Evolution 54: 897-905. [ Links ]
Risi J., y Galwey N.W., 1989. The pattern of genetic diversity in the Andean grain crop quinoa (Chenopodium quinoa Willd.). I. Associations between characteristics. Euphytica 37: 147-162. [ Links ]
Rojas W., Barriga P., y Figueroa H., 2000. Multivariate analysis of the genetic diversity of Bolivian quinua germplasm. Plant Genetic Resources Newsletter 122: 16-23. [ Links ]
Roldán-Ruiz I.,vanEeuwijkF.A.,GillilandT.J., Dubreuil P., Dillmann C., Lallemand J., De Loose M., y Baril C.P., 2001. Acomparative study of molecular and morphological methods of describing relationships between perennial ryegrass (Lolium perenne L.) varieties. Theoretical and Applied Genetics103: 1138-1150. [ Links ]
Winkel T., Lhomme J.P., Nina Laura J.P., Mamani Alcón C., Del Castillo C., y Rocheteau A., 2009. Assessing the protective effect of vertically heterogeneous canopies against radiative frost: the case of quinoa on the Andean Altiplano. Agricultural and Forest Meteorology 149: 1759-1768. [ Links ]

