











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkJournal of the Selva Andina Animal Science
versión impresa ISSN 2311-2581
J.Selva Andina Anim. Sci. vol.7 no.2 La Paz 2020
https://doi.org/10.36610/j.jsaas.2020.070200063
Comunicación Corta
Variaciones en el metabolismo energético de la alpaca (Vicugna pacos). Una evaluación por efecto del ayuno prolongado
Variations in the energy metabolism of the alpaca (Vicugna pacos). An evaluation by effect of prolonged fasting
Coila-Añasco Pedro Ubaldo1*![]() , Ruelas-Calloapaza Domingo Alberto1
, Ruelas-Calloapaza Domingo Alberto1![]() , Guerra-Aguilar Freddy1, Olaguivel Flores César Augusto2
, Guerra-Aguilar Freddy1, Olaguivel Flores César Augusto2![]() , Oha-Humpiri Filiberto3
, Oha-Humpiri Filiberto3![]()
1Facultad de Medicina Veterinaria y Zootecnia. Universidad Nacional del Altiplano-Puno. Dirección: Av. Floral 1153. Puno, Perú. Tel: +051-599430. druelasc@yahoo.es,fredwar77@hotmail.com
2Facultad de Medicina veterinaria. Universidad Nacional de San Cristóbal de Huamanga. Portal Independencia N° 57. Huamanga-Ayacucho. Perú. Tel: (066)312510 – (066)312230. olaguivelflores@gmail.com
3 Facultad de Medicina Veterinaria y Zootecnia. Universidad Nacional Micaela Bastidas de Apurímac. Avenida Inca Garcilazo de la Vega. Abancay 03001, Perú. Tel:+51 83 321965, capetillo_45@hotmail.com
*Dirección de contacto: Facultad de Medicina Veterinaria y Zootecnia. Universidad Nacional del Altiplano-Puno. Dirección: Av. Floral 1153. Puno, Perú. Tel: +051-599430. Móvil: +51 95196540
Pedro Ubaldo Coila-Añasco
E-mail address: pcoila@unap.edu.pe
Historial del artículo.
Recibido julio 2020.
Devuelto agosto 2020.
Aceptado septiembre 2020.
Disponible en línea, octubre 2020.
ID del artículo: 079/JSAAS/2020
J. Selva Andina Anim. Sci. 2020; 7(2):63-71.
Resumen
El estudio tuvo como objetivo de evaluar el efecto de un ayuno prolongado (5 días) sobre algunos componentes bioquímicos sanguíneos involucrados en el metabolismo energético. Se utilizaron 20 alpacas macho de dos años de edad en dos tratamientos (experimental y control). Se tomaron muestras de sangre en horas de la mañana por cinco días para cuantificar los niveles séricos de glucosa, proteínas y urea por espectrofotometría y cuerpos cetónicos mediante tiras reactivas en forma semicuantitativa. Los resultados muestran que el promedio de glucosa, proteínas y urea de las alpacas sometidas a ayuno son diferentes al grupo control (P≤0.05). Los niveles de glucosa disminuyen mientras que los niveles de proteínas y urea aumentan con el paso del tiempo. La presencia de acetoacetato no fue detectada durante los cinco días de ayuno. Se concluye que el ayuno prolongado en alpacas altera el proceso metabólico del animal, tanto cuantitativa como cualitativamente, modificándose los sustratos y productos implicados en la producción de energía.
Palabras clave: Ayuno, glucosa, proteína, urea, acetoacetato
Abstract
The study aimed to evaluate the effect of a prolonged fast (5 days) on some blood biochemical components involved in energy metabolism. Twenty male alpacas of two-year-old were used in two treatments (experimental and control). Blood samples were taken in the morning for five days to quantify serum glucose, protein and urea levels by spectrophotometry and ketone bodies using semiquantitative test strips. The results show that the average of glucose, protein and urea of the alpacas subjected to fasting are different from the control group (P≤0.05). Glucose levels decrease while protein and urea levels increase over time. The presence of acetoacetate was not detected during the five days of fasting. It is concluded that prolonged fasting in alpacas alters the animal's metabolic process, both quantitatively and qualitatively, modifying the substrates and products involved in energy production.
Keywords: Fasting, glucose, proteins, urea, acetoacetate.
Introducción
Durante años se ha reconocido que los Camélidos Sudamericanos (CSA), suelen tener concentraciones de glucosa en sangre más altas que los rumiantes domésticos, lo cual es desconcertante debido a que los CSA, al igual que los otros rumiantes, son también fermentadores, no es posible, pensar que los carbohidratos de la dieta escapen de la fermentación microbiana gástrica, por lo que estas especies dependen de la gluconeogénesis para suministrar el sustrato necesario a los tejidos dependientes de la glucosa. Además, en los últimos años, se han identificado enfermedades en alpacas y llamas, que parecen ser el resultado de un metabolismo anormal de carbohidratos, algunas de estas alteraciones se parecen a las de los otros animales, pero otras no, estas diferencias apoyan la conclusión de que los CSA procesan los carbohidratos de manera diferente a otros animales1.
Ante una disminución en el consumo de alimentos, el organismo animal activa mecanismos que conducen a la producción de sustratos que aseguren la provisión de energía a los órganos vitales, con el propósito de asegurar la sobrevivencia del animal, y, si el ayuno continúa, el metabolismo va cambiando tanto cualitativa como cuantitativamente2. Durante el ayuno, inicialmente los sustratos energéticos derivan de la degradación proteica y de grasas, pero en las siguientes fases la degradación proteica se ralentiza, maximizándose la lipólisis3. La gluconeogénesis es una vía metabólica que se encuentra activa permanentemente en los rumiantes, siendo el ácido propiónico el principal precursor de la síntesis de glucosa4. Asimismo, parece que las alpacas dependen de sus adaptaciones digestivas, metabólicas para obtener, conservar eficientemente la energía para sobrevivir en ambientes donde la alimentación disponible varía en calidad nutricional5.
Otra importante fuente de energía son los aminoácidos, los cuales se movilizan después de una hipoglicemia sanguínea con la finalidad de sostener la gluconeogénesis hepática y, por tanto, la producción de energía, debido a estas condiciones, la glucosa, los aminoácidos se mantienen, pero se movilizan grasas, cuerpos cetónicos (CC), para la producción de energía6. Por otro lado, la urea es el producto de degradación de los aminoácidos, en rumiantes esta urea puede ser reciclada y volver al rumen a través de la saliva o la pared ruminal, una vez presente en el rumen, es reconvertida en amoniaco la cual puede servir para la síntesis de aminoácidos por la microbiota ruminal, y estos, a su vez, en fuente de energía después de su absorción intestinal4.
La crianza de CSA constituye el principal medio de uso de productivo de grandes extensiones de pasturas naturales, en las zonas altas de los andes donde no es posible la crianza de otras especies domésticas ni de la agricultura, la alpaca, convierte con eficiencia la vegetación natural de estos ambientes en fibra y carne para el consumo humano7. No obstante, las condiciones de crianza de los CSA, la variabilidad estacional de los recursos alimenticios determinan que tengan que pasar por limitaciones nutricionales durante la época seca y, en muchos casos, por periodos de ayuno prolongados8. La alpaca, al igual que los demás CSA, son especies que han evolucionado eficientemente en ambientes con serias limitaciones ambientales, pero que aún no son bien entendidos9. Durante el ayuno, estos animales pueden utilizar ácidos grasos libres (AGL) y CC de manera más efectiva y no necesitan depender completamente de las vías bioquímicas de los rumiantes clásicos para sus requerimientos de energía y glucosa10.
En un estudio se reportó que las concentraciones plasmáticas de glucosa, creatinina y urea en llamas fueron más altas que ovejas y cabras, pero muy bajas en CC (β-hidroxibutirato)11. El alto nivel de glicemia, se puede explicar en parte por una pobre respuesta a la insulina o por una sensibilidad reducida del tejido a la insulina12. Además, los CSA cubren sus requerimientos de glucosa a través del proceso de gluconeogénesis a partir de los aminoácidos de la proteína microbiana y no del propionato como ocurre en los rumiantes9.
Estas características implican la necesidad de un esquema dietético basado en la relación proteína-grasa y no en la relación proteína-carbohidratos como en los rumiantes9. Por esto, el estudio se planteó con el objetivo de determinar algunas variaciones en el metabolismo energético por influencia del ayuno prolongado en alpacas mediante las determinaciones séricas de glucosa, proteínas totales, urea y CC.
Materiales y métodos
El estudio se realizó en el Centro de Investigación y Producción La Raya de la Universidad Nacional del Altiplano-Puno (UNAP), ubicado en el distrito de Santa Rosa, provincia de Melgar, departamento de Puno, Perú, a una altitud de 4200 m. y entre las coordenadas geográficas de 10°1333 S y 20°5712 O. El centro se encuentra en la región de Puna, con dos épocas bien marcadas: época de lluvias (de diciembre a marzo) y época de secas (de abril a marzo). La precipitación pluvial promedio es de 932 mm/año y la temperatura varía entre -7 ºC y 15 ºC. La precipitación pluvial y temperatura determinan que el clima sea del tipo húmedo frío. En el suelo de la zona predominan las especies Stipas, Festucas, Muhlenbergias, Calamagrostis y Distichias, que sirven de sustento alimentico para la crianza extensiva de alpacas y llamas.
Figura 1 Rebaño de alpacas machos de dos años de edad (tuis)

El estudio fue realizado en el mes de mayo del 2018. El análisis de las muestras se realizó en el Laboratorio de Bioquímica de la Facultad de Medicina Veterinaria y Zootecnia de la UNAP.
Figura 2 Grupo de animales en ayuno

Se utilizaron 20 alpacas machos de dos años de edad de raza Huacaya en aparente buen estado de salud, con un peso vivo promedio de 47.7 kg. Los 20 animales se distribuyeron en dos tratamientos de 10 animales cada uno en forma aleatoria (experimental y control), los cuales fueron mantenidos en corrales construidos con malla y postes de madera con un área de 25 m2. El grupo experimental permaneció confinado durante los cinco días que duró el experimento, sin acceso a ningún tipo de alimento excepto de agua y sin periodo de acostumbramiento previo; entretanto, que los animales del grupo control fueron pastoreados conjuntamente con los demás animales, consumiendo los pastos naturales de la zona. Los animales fueron proporcionados por el Instituto de Investigación y Promoción de Camélidos Sudamericanos (IIPC) de la UNAP.
Figura 3 Animales del grupo control

Se tomaron muestras de sangre a las 0, 24, 48, 72 y 96 h de iniciado el experimento (hora 0 a partir de las 06:00 h) a todos los animales. Las muestras en el grupo control se obtuvieron antes de su salida a los campos de pastoreo. La sangre (5 mL) se obtuvo de la vena yugular en tubos sin anticoagulante, y se trasladó al laboratorio en una caja refrigerada. El suero se obtuvo por centrifugación a 1600 g por 10 min, y se decantó en viales criogénicos para su conservación a -20 ºC hasta su procesamiento. Para la utilización de animales y obtención de muestras sanguíneas se contó con la autorización del Comité de Ética de la Facultad de Medicina Veterinaria y Zootecnia de la Universidad Nacional del Altiplano.
Las determinaciones de glucosa, proteínas totales y urea en el suero sanguíneo se realizaron mediante colorimetría-espectrofotometría utilizando reactivos de Wiener Lab®. La determinación de cuerpos cetónicos se hizo en forma semicuantitativa utilizando tiras reactivas Multistix 10 SG Siemens ® (Bayer) que detecta la presencia acetoacetato con una sensibilidad de 5 a 10 mg/dL y una reproducibilidad de 95% según el inserto para uso del paquete.
El experimento fue conducido en un diseño bloque completo al azar con sub-muestreo (anidado), considerando como tratamiento a los grupos (experimental y control) y bloques a las horas de muestreo (0, 24, 48, 72 y 96 h). Para la comparación de medias de los efectos principales (grupos y horas de muestreo) se utilizó la prueba de significancia de Duncan13 y para los efectos simples el LS Means del software SAS v. 9.114.
Resultados
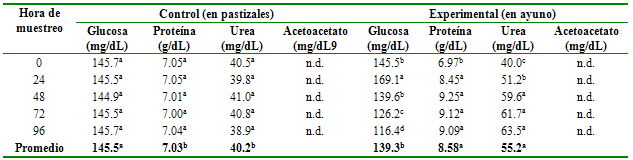
Las concentraciones de glucosa, proteína, urea y acetoacetato por grupo según la hora de muestreo (ayuno) se muestran en la tabla 1. Los niveles de glucosa fueron inferiores en animales con ayuno respecto a animales sin ayuno (p≤0.05); y existe diferencias entre las horas de muestreo (p≤0.01) para el caso de los animales en ayunas, pero no para los animales del grupo control. En contraste, los niveles de proteínas y urea son más altos en animales con ayuno que en el grupo control (p≤0.01), así como entre los días de muestreo para los animales en ayunas, pero no para sus controles (p≤0.01). Las lecturas de concentración de cuerpos cetónicos fueron negativos para ambos tratamientos (no detectados).
| Tabla 1 Perfil bioquímico en suero sanguíneo de alpacas con y sin ayuno |
|
|
| n.d.: No detectado |
Discusión
El aumento de glucosa a las 24 h del ayuno coincide con lo reportado por Wensvoort et al.10 quienes indican que en condiciones de ayuno aumenta la concentración de glucosa sanguínea en llamas, camellos y ovinos. Por su parte, Tadich et al.15, confirman que el aumento de la glucosa sanguínea en el primer momento son debidos principalmente al estrés y no al efecto del ayuno, ya que la hipoglucemia inicial actúa como factor liberador de catecolaminas, promoviendo la glucólisis y la gluconeogénesis, durante el estrés hay un aumento del cortisol que a su vez es hiperglicemiante. De igual modo, Crossley et al.16, al evaluar la respuesta fisiológica adaptativa de la alpaca al cambio de ambiente, encontraron que la concentración de glucosa sanguínea aumenta, hecho que se atribuye al estrés causado por el ayuno, al encierro y a la toma de muestras.
El promedio de glucosa al inicio del experimento, se encuentra dentro del rango establecido como normales de 112-260 mg/dL por Siguas et al.17, y, ligeramente inferior a los reportados por Kiani et al.11 y Al-Rehaimi et al.18 quienes encontraron promedios de glucosa de 127.8 y 138.0 mg/dL en llamas y camellos, respectivamente, diferencias que se atribuyen a la especie. Sin embargo, es necesario indicar que la glicemia en alpacas, al igual que en otros camélidos, es más alta que otros rumiantes (vaca, oveja y cabras) tal como lo reporta Kiani et al.11 Esta hiperglicemia fisiológica en alpacas también se debería a la pobre respuesta de la insulina y a la baja concentración de insulina en el suero, propuesta por Cebra et al.1.
Los niveles de glucosa van disminuyendo después de las 24 h, lo que concuerda con Albero et al.2, quienes indican una disminución rápida de la glicemia, pero que tras 48-72 h de ayuno se produce un declinar lento hasta llegar a estabilizarse. Se señala que, en esencia, toda la glucosa disponible para los rumiantes, indistintamente de la dieta, procede de la gluconeogénesis, siendo el precursor más importante de glucosa el ácido propiónico, y sumado a la constante gluconeogénesis, los rumiantes también cubren sus necesidades de glucosa mediante una eficaz conservación de la misma3.
El nivel sérico de proteínas a las 0 horas (6.96 g/dL) coincide con los reportes de Oblitas et al.19 quienes reportan una concentración promedio de 6.6 g/dL. En los animales privados de alimento, los niveles de proteínas totales suben a partir de las 24 h. sin embargo, este aumento no se debería a un incremento real de las proteínas en el plasma sanguíneo sino a la deshidratación del individuo20. Otros autores señalan que en el ayuno, el riñón se transforma en la principal fuente de eliminación de agua, observándose que en los 5 a 10 primeros días se observa un balance fuertemente negativo de agua, disminuyendo incluso la densidad de la orina21.
La concentración sérica de urea al inicio del experimento (40.25 mg/dL), se encuentra dentro del rango reportado por Siguas et al.17 (32.2 y 89.8 mg/dL). Asimismo, es similar al valor reportado por Kiani et al.11 quien determina una uremia de 40.2 mg/dL para llamas.
Los niveles de urea aumentan conforme transcurren los días de ayuno. Según Bustamante22, cuando se perciben factores estresantes, se inicia una cascada de eventos, la mayoría relacionados con el eje hipotálamo-hipófisis-corteza adrenal, que finalmente lleva a la liberación de corticoides con determinados efectos fisiológicos, entre ellos la gluconeogénesis hepática catalizando aminoácidos y proteínas y, por lo tanto, una subida en los valores de concentración de urea. Por su parte Chowdhury & Ørskov23 indican que cuando el ayuno se agudiza en rumiantes, se estimulan los glucoreceptores en el hipotálamo lo que estimula a la médula adrenal para aumentar la secreción de adrenalina, ésta, a su vez, activa la glucogenólisis, lipólisis, gluconeogénesis, por lo tanto, se producirá un mayor uso de intermediarios para producir energía, dando como resultado una mayor excreción urinaria de nitrógeno durante el ayuno. De la misma forma, Wensvoort et al.10 indican que el incremento en los niveles de urea durante el ayuno, es señal de un aumento de la oxidación de aminoácidos para suministrar precursores de glucosa por la gluconeogénesis.
Es probable que la cetogénesis se en periodos más prolongados de ayuno, razón por la cual no fue detectada su presencia en el presente estudio. El resultado concuerda con Tadich et al.15, quienes tampoco hallaron variaciones en cuerpos cetónicos (β-hidroxibutirato) en el ayuno de novillos durante su transporte al matadero, posiblemente debido a la utilización del pool circulante de β-hidroxibutirato aportado por el rumen. Por su parte, Wensvoort et al.10 indican que los camélidos podrían tener la capacidad de suministrar suficientes metabolitos para el mantenimiento del ciclo del ácido cítrico y que podrían usar menos u otros metabolitos en relación a otras especies de rumiantes. Al respecto, se señala que durante los primeros días de ayuno, las reservas energéticas del animal y la gluconeogénesis sirven para proveer de energía al animal y la cetogénesis es activada cuando se agotan estas reservas.24
Chandrasena et al.25 reporto concentraciones plasmáticas de 0.089 y 0.148 mg/dL de β-hidroxibutirato y acetoacetato en camellos, que son aproximadamente 33 y 4 veces más bajos que para los valores respectivos en ovejas, mantenidos en las mismas condiciones, sugiriendo que la cetogénesis en camellos es muy baja, hecho que también estaría ocurriendo en alpacas, puesto que a las 96 h de ayuno los cuerpos cetónicos permanecen indetectables. Del mismo modo, Kiani et al.11 concluyeron que los niveles de β-hidroxibutirato en sangre de llamas, era extremadamente bajo para un animal rumiante, tal como el encontrado en alpacas del presente estudio. Entonces, es probable que el metabolismo de CC en la alpaca muestre variaciones con respecto al de otros rumiantes.
Fuente de financiamiento
El estudio fue financiado con recursos propios de los autores.
Conflictos de intereses
Los autores declaramos no tener conflictos de intereses con respecto a la investigación, autoría y/o publicación de este artículo.
Agradecimientos
Se agradece al Instituto de Investigación y Promoción de Camélidos Sudamericanos (IIPC) de la Facultad de Medicina Veterinaria y Zootecnia de la UNA-Puno por habernos brindado los animales para el estudio. De igual forma, a los técnicos Martín Chaiña y Vicente Flores por su apoyo en la construcción de las instalaciones y toma de muestras del estudio.
Aspectos éticos
El estudio contó con la aprobación del Comité de Ética de la Facultad de Medicina Veterinaria y Zootecnia de la Universidad Nacional del Altiplano y se siguió sus pautas establecidas por este Comité.
Literatura citada
1. Cebra CK, Tornquist SJ, Van Saun RJ, Smith BB. Glucose tolerance testing in llamas and alpacas. Am J Vet Res 2001;62(5):682-6. DOI: https://doi.org/10.2460/ajvr.2001.62.682 [ Links ]
2. Albero R, Sanz A, Playán J. Metabolismo en el ayuno. Endocrinol Nutr 2004;51(4):139-48. DOI: https://doi.org/10.1016/S1575-0922(04)74599-4 [ Links ]
3. Cunningham J. Fisiología Veterinaria. Tercera ed. Madrid, España: ELSEVIER; 2003. DOI: https://doi.org/10.1016/B978-84-9022-317-8/00056-7
4. Dukes HH. Dukes´phisiology of domestic animals. 13th ed. John Wiley & Sons, Inc.; 2015.
5. Lund KE, Milton JTB, Maloney SK, Blache D. Alpacas fed calcium propionate seem to moderate their energy intake. J Anim Physiol Anim Nutr (Berl). 2014;98(6):1088-94. DOI: https://doi.org/10.1111/jpn.12180 [ Links ]
6. Mompeán O, Manjón Collado MT, Pérez Sánchez A, Pérez Huertas R. Fisiopatología del ayuno corto y prolongado y del estrés. Rapd Online 2009; 32(1): 45-58. [ Links ]
7. Quispe EC, Rodríguez TC, Iñiguez LR, Mueller JP. Producción de fibra de alpaca, llama, vicuña y guanaco en Sudamérica. Anim Genet Resour Inf 2009;45:1-14. DOI: https://doi.org/10.1017/s1014233909990277 [ Links ]
8. San Martín F, Van Saun RJ. Applied digestive anatomy and feeding behavior. In: Cebra C, Anderson D, Tibary A, Van Saun R, Johnson LR, editors. Llama and alpaca care: Medicine, surgery, reproduction, nutrition, and herd health. Philadelphia: Elsevier; 2014. p. 51-8. DOI: https://doi.org/10.1016/B978-1-4377-2352-6.00008-0
9. San Martín F. Adaptación nutricional y metabólica de los camélidos sudamericanos. En: Aranibar Aranibar MJ, Urviola Sánchez JM, editores. VII Congreso mundial en camélidos sudamericanos. Libro de resúmenes: 28, 29 y 30 de octubre de 2015. Facultad de Medicina Veterinaria y Zootecnia [Internet]. Puno: Universidad Nacional del Altiplano; Corporación Meru EIRL; 2015. [citado 3 de mayo de 2019]. p. 45. Recuperado a partir de: file:///C:/Users/usuario/Downloads/LibroResumenes%20(1).pdf
10. Wensvoort J, Kyle D, Ørskov E, Bourke D. Biochemical adaptation of camelids during periods where feed is withheld. Rangifer 2001;21(1):45-8. DOI: https://doi.org/10.7557/2.21.1.1527 [ Links ]
11. Kiani A, Alstrup L, Nielsen MO. Differential metabolic and endocrine adaptations in llamas, sheep, and goats fed high- and low-protein grass-based diets. Domest Anim Endocrinol 2015;53:9-16. DOI: https://doi.org/10.1016/j.domaniend.2015.03.006 [ Links ]
12. Elmahdi E, Sallmann HP, Fuhrmann H, von Engelhard W, Kaske M. Comparative aspects of glucose tolerance in camels, sheep, horses and ponies. Comp Biochem Physiol A Physiol 1997;118A(1):147-51. DOI: https://doi.org/10.1016/s0300-9629(96)00449-5 [ Links ]
13. Duncan DB. Multiple Range and Multiple F Tests. Biometrics. 1955;11(1):1-42. DOI: https://doi.org/10.2307/3001478 [ Links ]
14. Inc. SI. SAS/STAT® 9.1 Users Guide. NC: SAS Institute Inc.; 2004.
15. Tadich N, Gallo C, Echeverría R, van Schaik G. Efecto del ayuno durante dos tiempos de confinamiento y de transporte terrestre sobre algunas variables sanguíneas indicadoras de estrés en novillos. Arch Med Vet 2003;35(2):171-85. DOI: https://doi.org/10.4067/S0301-732X2003000200005 [ Links ]
16. Crossley JC, Marin MP, Ferrando G, Raggi LA. Modificaciones adaptativas de algunas constantes fisiológicas de alpaca (Lama pacos) sometidas a cambios de ambiente. Arch Zootec 1994;43 (163):215-23. [ Links ]
17. Siguas O, Paucar R, Olazabal J, San Martín F, Vélez V. Valores bioquímicos sanguíneos en alpacas en dos épocas del año en condiciones de Huancavelica: Aportes al perfil metabolico de la especie. Arch Latinoam Prod Anim [Internet]. 2007 [citado 5 de octubre de 2019];15 (Suppl. 1): 496-497. Recuperado a partir de: https://www.researchgate.net/publication/284411697_Valores_bioquimicos_en_alpacas_en_dos_epocas_del_ano_en_condiciones_de_Huancavelica_aportes_al_perfil_metabolico_de_la_especie
18. Al-Rehaimi AA, Al-Ali AK, Mutairy AR, Dissanayake AS. A comparative study of enzyme profile of camel (Camelus dromedarius) hump and sheep (Ovis aries) tail tissues. Comp Biochem Physiol 1989;93B(4):857-8. DOI: https://doi.org/10.1016/0305-0491(89)90057-6 [ Links ]
19. Oblitas GF, Pedrozo PR, Wittwer MF, Böhmwald H, Ludwing H. Valores sanguíneos en alpacas (Lama pacos) reintroducidas en el sur de Chile. Vet Méx 1998;29(4):411-4. [ Links ]
20. Hervás Abad E, Sánchez Polo MT, García Lopez PJ. Nutrición, ayuno y ejercicio [Internet]. Fundacion para la Formacion e Investigacion Sanitarias de la Region de Murcia. 2012 [citado 3 de mayo de 2019]. Recuperado a partir de: http://www.ffis.es/volviendoalobasico/tema_15_nutricin_ayuno_y_ejercicio.html [ Links ]
21. Saz Peiró P, Ortiz Lucas M. Fisiología y bioquímica en el ayuno. Med Natur 2007;1(1):10-9. [ Links ]
22. Bustamante H. Determinación del efecto de diferentes tiempos de ayuno y transporte terrestre sobre algunas variables sanguíneas indicadoras de estrés en bovinos en el período Otoño-Invierno [tesis licenciatura]. [Valdivia]: Universidad Austral de Chile; 2001.
23. Chowdhury SA, Ørskov ER. Implications of fasting on the energy metabolism and feed evaluation in ruminants. J Anim Feed Sci 1994;3(3):161-9. DOI: https://doi.org/10.22358/jafs/69830/1994 [ Links ]
24. Dukes HH. Dukes Physiology of domestic animals. 30o. (Reece W, ed.). Oxford: John Wiley & Sons, Inc.; 2015.
25. Chandrasena LG, Emmanuel B, Hamar DW, Howard BR. A comparative study of ketone body metabolism between the camel (Camelus dromedarius) and the sheep (Ovis aries ). Comp Biochem Physiol 1979;64(1):109-12. DOI: https://doi.org/10.1016/0305-0491(79)90192-5 [ Links ]
______________
Nota del Editor:
Journal of the Selva Andina Animal Science (JSAAS) se mantiene neutral con respecto a los reclamos jurisdiccionales publicados en mapas y afiliaciones institucionales.
