











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
Permalink
INTRODUCCIÓN
Las cianobacterias son microorganismos procariotas con capacidad fotosintética, que ocupan diversos nichos ecológicos, adaptándose a diversos ambientes extremos, como altas o bajas temperaturas, pH muy ácido o básico, altas concentraciones de sal y desecación (Wada et al., 2013). Estos organismos son recursos biológicos importantes por su capacidad de producir una gran gama de compuestos bioactivos como ser: antivirales, fitohormonas, antifúngicos, sideróforos, inmunosupresores, alguicidas, fotoprotectores, y antimicrobianos. Pero además han llamado la atención por su capacidad de formar floraciones densas y producir toxinas comúnmente llamadas cianotoxinas, las cuales son dañinas para la salud humana y otros organismos (Wada et al., 2013; Whitton & Potts, 2012).
En la presente recopilación, se describirá la importancia de la cianotoxina microcistina en salud humana y la implicación del riego en sembradíos agrícolas. Además de desarrollar su degradación por procesos de fotocatalíticos y procesos bacterianos. Finalmente mencionar algunos métodos para la detección de las microcistinas.
METODOLOGÍA
La determinación del estado de arte para el tema de cianobacterias se basó en una búsqueda bibliográfica en la base de datos especializada como Elservier, Springer, Google académico y MDPI basadas en palabras clave en español e inglés "microcistinas", "degradación de MC" y “cuantificación y detección de MC”.
1. GENERALIDADES
a. Definición de cianobacterias
Las cianobacterias también conocidas como algas verde azuladas, son los primeros organismos fotosintéticos en la tierra, que han contribuido en la producción de oxígeno durante los últimos 3 millones de años (Zahra et al., 2020). Se encuentran presentes en distintos recursos hídricos como agua dulce, agua salobre y aguas residuales, por lo general en cantidades bajas o moderadas, pero a razón de la actividad humana hay un incremento de las poblaciones cianobacterianas formando floraciones, debido a que estos microrganismos tienen una gran taza de crecimiento, llegando aprovechar los nutrientes generados por la eutrofización. Los géneros más comunes son: Anabaena, Aphanizomenon, Cylindrospermopsis, Lyngbya, Microcystis, Oscillatoria, Phormidium and Planktothrix (WHO, 2015; Zahra et al., 2020).
Además de ser una fuente esencial de oxígeno, las cianobacterias se han considerado una fuente de nutrientes y biocombustible, también se destacan por su potencial de producir distintos metabolitos secundarios. Entre las primeras investigaciones de estos metabolitos se observa su capacidad de sintetizar cianotoxinas (Figura 1) que continúan llamando la atención por su gran impacto en la salud y economía (Mazard et al., 2016).

b. Cianobacterias productoras de cianotoxinas
A pesar de la importancia que pueden tener la cianobacterianas en el campo biotecnológico, no se puede ignorar el aspecto nocivo de ciertas especies, por su capacidad de sintetizar cinaotoxinas, las cuales pueden causar intoxicación aguda grave en mamíferos afectando distintos sistemas/aparatos como el hepato-pancreático, digestivo, endocrino, dérmico y nervioso. Además la formación de floraciones también puede alterar la claridad del agua, afectando negativamente el habitad de su entorno, generando un agotamiento de oxígeno generando un ambiente anóxico, provocando la muerte de los peces (Paerl & Otten, 2013). La diversidad de cianobacterias es muy extensa, de las cuales solo un determinado grupo de cianobacterias tiene la capacidad de producir estas toxinas, además algunos géneros sintetizan más de una cianotoxina (tabla 1) (Chorus, 2001; Paerl & Otten, 2013).
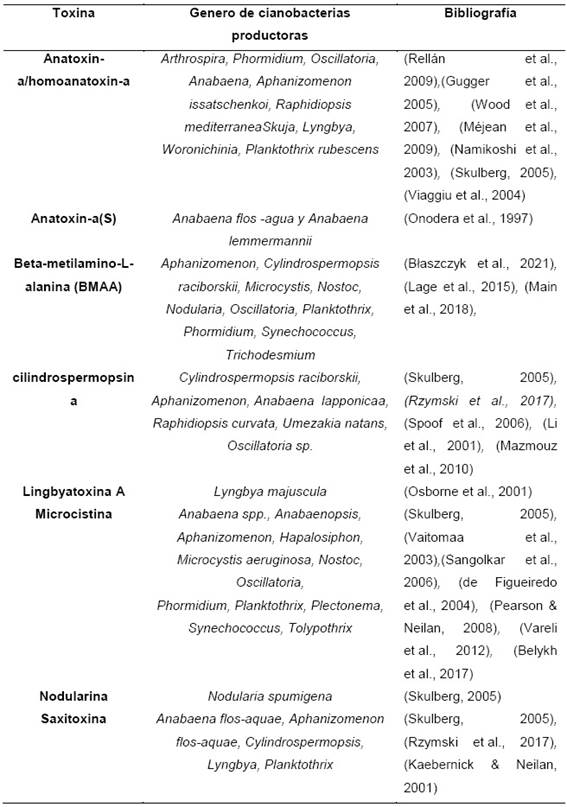
c. Importancia de las cianotoxinas
La exposición de estas toxinas afecta a distintos organismos vivos, alterando su entorno y su fisiología. En el ser humano y otros mamíferos han observado que la exposición a de estos metabolitos provoca diversas enfermedades con síntomas como: Dolor abdominal, vómitos, diarrea, irritación en la piel, debilidad, dolores musculares y en otros (Drobac et al., 2013).
Estos metabolitos que pueden encontrarse intracelularmente, como es el caso de la anatoxina-a y las variantes de microcistina sintetizados en la etapa de desarrollo de la floración, pero también pueden ser extracelulares y presentando más complicaciones para su eliminación, ya que son absorbidos por la arcilla y la materia orgánica presente en la columna de agua, siendo el caso de Cylindrospermopsis, Aphanizomenon y Umezakia (Bhattacharyya et al., 2015).
2. Microcistina y su impacto en sembradíos agrícolas
Entre las diversas cianotoxinas las microcistinas (CM) son extremadamente tóxicas, alterando los cuerpos de agua, los ecosistemas marinos, y poniendo en riesgo la salud pública, registrándose envenenamientos y muertes, afectando destinos órganos como el intestino, cerebro, riñones, pulmones, corazón, sistema reproductivo y teniendo como principal diana al hígado (Massey et al., 2018).
La contaminación por MC puede llegar afectar toda la red de los distintos cuerpos de aguas continentales, afectando las desembocaduras de los ríos, lagunas costeras y también encontrarse afectado el trasporte de la red de agua dulce (El Herry et al., 2007). Los efectos de riego sobre sembradíos agrícolas con aguas contaminadas, presentan una reducción significativa en las raíces, alteraciones fisiológicas y morfológicas, una reducción del tamaño con concentraciones bajas de clorofila y una repercusión en el ciclo de vida de las plantas agrícolas. Además han observado una absorción en el tejido vegetal y consecuentemente una bioacumulación, aunque estas alteraciones pueden variar de planta en planta, debido a que algunas especies tienen mecanismos de desintoxicación (Crush et al., 2008). Por lo tanto su acumulación en sembradíos agrícolas e incluso en organismos acuáticos, no pasa desapercibido ya que ingresan a la cadena alimentaria, tanto para animales como para los seres humanos, generando una ingesta con concentraciones que pueden exceder los límites tolerables para la salud (Saqrane & Oudra, 2009).
a. Propiedades químicas
Las MCs poseen una estructura cíclica, en común se presentan de 7 aminoácidos unidos a péptidos formando un heptapéptidos monocíclicos, con una estructura general de ciclo-(-D-Ala-LXD-MeAsp-LZ-Adda-D-Glu-Mdha), donde X y Z representan aminoácidos altamente variables, D-MeAsp representa ácido D-eritro-b-metilaspártico, Adda representa (2S, 3S, 8S, 9S) 3-amino-9 metoxi-2,6,8-trimetil-10- ácido fenildeca-4,6-dienoico y Mdha representa N-metildehidroalanina (Fig 2). Debido a su estructura tiene la capacidad de resistir factores químicos y físicos, teniendo estabilidad en distintos ambientes de temperatura, sequedad, pH, e incluso a la hidrolisis bioquímica por la pepsina, tripsina y quimiotripsina. Estas moléculas tiene un peso molecular entre 800-1100Da con carácter hidrofílico, dificultando el ingreso a través de la membrana celular y entre las más representativas por su frecuencia y toxicidad son las MC-LR, MC-RR y MC-YR (Chen & Xie, 2016; Massey & Yang, 2020). Finalmente destacando de su estructura la región X y Z que le brindan gran versatilidad a las MC, debido a ello se han identificado 246 variantes con distintos grados de toxicidad (Spoof & Catherine, 2016).
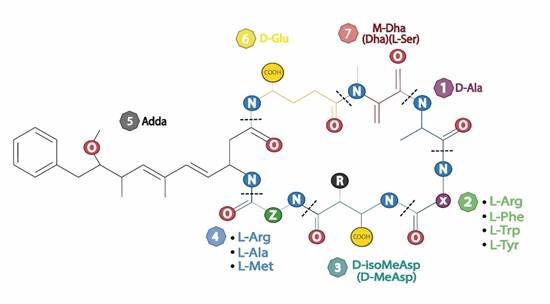
b. Toxicidad
Los efectos de MC sobre el metabolismo y fisiología varían según: La especie, antecedes genéticos, tipo de célula, niveles y duración en la exposición. Por sus características químicas el ingreso la MC dentro de las células se da principalmente a través de unos transportadores específicos, conocidos como la familia de polipéptidos transportadores de aniones orgánicos (Oatps) (Chen & Xie, 2016). La MC-LR son inhibidores altamente
específicos de las fosfatasas (PP), particularmente la PP1 y PP2A, las cuales están involucradas en la desfosforilación de proteínas reguladoras, destacando su importancia en la regulación celular tanto en mamíferos como en plantas (MacKintosh et al., 1990). Por otro lado es un desencadenante de daño oxidativo por las especies reactivas de oxigeno (ROS), debido a una disminución del glutatión (GSH), el cual cumple múltiples funciones tales como: Eliminación de radicales libres, conjugación con xenobióticos para su desintoxicación y está vinculado con la organización del citoesqueleto, entonces no es de sorprenderse que su disminución genere estas especies reactivas, provocando fuga de LDH y un aumento malondialdehído (MDA) por peroxidación lipídica provocando citotoxicidad (Ding & Nam Ong, 2003). Entre tanto la genotoxicidad inducida por MC puede estar asociado con la transformación hacia la malignidad celular, debido al da daño cromosómico permanente, provocando fragmentación cromosómica o aneuploidia (Dias et al., 2014). Finalmente, la exposición prolongada de esta cianotoxina puede desencadenar la apoptosis dentro de la célula, debido a la alteración en distintos organelos vinculados con este proceso e incluso provoca el des-ensamblaje del citoesqueleto, generando alteraciones morfológicas severas. En las mitocondrias se observa formación de ROS, perdida de potencial mitocondrial (MMP) y liberación de factores apoptóticos debido a la transición de permeabilidad mitocondrial (MPTC). Otras alteraciones observadas son hinchazón y vacuolización del retículo endoplasmático (RE), que juntamente con el aparto de Golgi detectan y señalizan las vías de reparación o muerte celular (Alverca et al., 2009; Ding et al., 2001).
c. Biosíntesis
La biosíntesis de MC está regulado por un policétido híbrido (PKS/NRPS) compuesto por un complejo de proteínas multifuncionales, que están agrupados y coordinadas en sitios enzimáticos formando módulos (Tillett et al., 2000). Su regulación está condicionada por factores ambientales y nutricionales, como altas concentraciones de nitrógeno y un déficit de hierro generan síntesis de MC. La región reguladora mcy también posee motivos de secuencia para proteínas de unión a ADN Fur (regulador de captación férrica) y NTC A (regulador de nitrógeno total), donde un incremento o deceso de concentración estimulan a la vez a la región myc (Neilan et al., 2013).
Para la síntesis de proteínas se tiene un grupo de genes de aproximadamente 55KDa e incluyen 10 genes (mcy A-J). Donde mcy A y myc D presenta una bidireccionalidad, y además el mcy presenta dos operones (myc A-C y myc D-J). Los péptidos no ribosómicos sintasa (NRPS), están involucrados con myc A-C, policétido sintasa (PKS) con los genes myc D, los híbridos PKS/NRPS myc E y G, epimerización myc F, deshidratación myc l, O-metiltransferasa myc J y un transportador ABC myc H (Tillett et al., 2000; Zhou et al., 2021).
La síntesis comienza con la formación de los aminoácidos Adda, mediante la captura de distintos precursores, como el ácido ciánico, fenilalanina y cumarato, mediantes los módulos Myc G, D y F se adiciona malonil-CoA, posteriormente la cadena lateral de aminoácidos Adda es modificada por metilación gracias a Mcy J (O-metiltransferasa). La cadena metilada es transformada en β-aminoácidos por el módulo Myc E aminotransferasa). Después de la síntesis del grupo Adda se activan y se adicionan aminoácidos a la cadena lateral por 6 módulos de adenilación, proteína transportadora, metiltransferasa, aminotransferasa, condensación y epimerización. También se presenta una modificación de la D-Glu por D-MeAps mediante la Mcy F (Racemasa) y la Mcy l (2-hidroxiacido deshidrogenasa), finalmente la cadena lineal alargada de aminoácidos es ciclada, transferida y regulada por el transportador ABC de la Myc H (Zhou et al., 2021).
3. Tratamientos de degradación de microscistina
a. Fotocatálisis
Las MCs pueden sufrir distintas vías de degradación, pero entre las que se destacan es la vía fotocatalítica, la cual puede ser directa o sensibilizada. En la fotocatálisis directa el compuesto orgánico absorbe la luz para su degradación, pero se ha observado que no es muy significativo en relación con la fotocatálisis sensibilizada, en donde a través de reacciones con intermediarios reactivos (IR), se genera distintas especies reactivas como ser: ROS, radical hidroxilo (*OH), halógenos reactivos (RHS) y radicales de carbonato, que contribuyen a una degradación más eficiente. El radical más significativo generado en el espectro UV-A, UV-B y PAR es *OH el cual actúa en cuatro regiones de las MCs, en el anillo de benceno (formando mono-dihidroxilados), grupo dieno-conjugado (formando 1,2 y 1,4 dioles isoméricos tetrahidroxilos), grupo metoxi en los aminoácidos Adda (genera radicales peroxilo) y el doble enlace carbono-carbono de los aminoácidos Mdha (formando productos bihidroxilados). Además de los radicales *OH otros compuestos como la ficocianina, genera una transferencia de energía la región dieno de la porción Adda, formando isómeros 6(z)-Adda MC-LR y puede transferir electrones al oxigeno formando un radical superóxido singlete, que ataca el grupo Adda de la MC formando hidroxiperóxidos que ya no son tóxicos (Kurtz et al., 2021). La fotocatálisis mediada por oxido de titanio ( TiO 2 ) (Fig 3A) promete ser un catalizador de importancia para generar ROS, con las nuevas alternativas de dopaje en la matriz sustituyendo el oxígeno (O) por carbono (C) se amplía el espectro de trabajo en la región VIS, lo que conduce a una mineralización de MC-LR hasta CO 2 , H 2 O he iones inorgánicos como ser NO 3 − y SO 4 −2 (Fotiou et al., 2016). Entre los problemas asociados en el proceso de fotocatálisis se presentan condiciones experimentales estrictas, supervisión experta, toxicidad de sub-productos y requiere energía adicional para el proceso por la materia orgánica presente que igual es tratada (Kumar et al., 2018).
b. Degradación biológica bacteriana
Los tratamientos fisicoquímicos por oxidación avanzada y convencional para MC, son generalmente costosos, ineficaces en su tratamiento, lo que ha dado paso al tratamiento biológico siendo entre sus características, respetuosos con el medio ambiente, por su eficiencia en la eliminación de metabolitos. Las bacterias que han llamado por su capacidad de metabolizar cianotoxinas como MCs, entre los géneros que se destacan son: Acinetobacter, Arthrobacter, Bacillus, Novosphingobium, Paucibacter, Pseudomonas, Sphingopyxis y Stenotrophomona. Estos grupos de bacterias se caracterizan por poseer el grupo de genes mlr con cuatro genes mlr ABCD. El gen más destacado es el gen mlr A el cual se encarga de sintetizar una metaloproteasa, que realiza la escisión del enlace Adda-Arg, convirtiendo a la MC-LR en una MC-LR lineal, su importancia de la enzima radica en que se encarga de debilitar la estructura cíclica de la MC-LR, la cual le dotaba de resistencia a distintos factores fisicoquímicos. El gen mlr BEI sintetiza una serina peptidasa, hidrolizando el enlace Ala-Leu formando tetrapéptidos (H-Adda-Glu-Mdha-Ala-OH). El gen mlr C sintetiza una metalopéptidasa con capacidad de hidrolizar el tetrapéptido en péptidos y aminoácidos pequeños. Además, se sabe que el gen mlr D sintetiza un transportador de la familia PTR2, que facilita el transporte de MC y los productos de su degradación (Fig 3B). Los productos formados pueden ser muy variables entre las distintas especies pero en ellas se destaca a la cepa Sphingopyxis sp. por su capacidad de degradar al grupo Adda hasta Acetil Coa y finalmente CO 2 , demostrando una eliminación total de la MC-LR (Kormas & Lymperopoulou, 2013; Massey & Yang, 2020).

4. Métodos de detección de microcistina
El monitoreo para la evaluación de MCs en la calidad de agua a menudo es complicada debido a que se pueden presentar múltiples variantes y a bajas concentraciones. En la actualidad se presentar numerosos estudios para su evaluación, siendo métodos sensibles, rápidos y confiables, de los cuales los más destacados serán descritos en esta sección (Massey et al., 2020).
a. Ensayo de inhibición de la proteína fosfatasa (PPI)
Este ensayo aprovecha la capacidad de las MCs de inhibir las enzimas de la defosforilzación de las fosfoproteínas intracelulares principalmente la PP1 y la PP2A. Permitiendo cuantificar el grado de inhibición mediante un ensayo colorimétrico, utilizando como sustrato al fosfato de p-nitrofenol. La técnica presenta una buena sensibilidad (Tabla 2), siendo viable para análisis de aguas permitiendo un análisis por debajo del valor aceptable de 1 µg/L según la Organización Mundial de la Salud (OMS) (Heresztyn & Nicholson, 2001). Las limitaciones de esta técnica están enfocadas en la poca información que brinda, sobre la toxicidad de las variantes de MC, además requerir un método adicional de confirmación para un análisis de especificidad. Aun así continua siendo una técnica sencilla, rápida, de bajo costo y altamente sensible (Massey et al., 2020).
b. Inmunoensayo
Los métodos inmunológicos son una gran herramienta por su alta sensibilidad, detectando una amplia gama de analitos, siendo selectivo por la interacción antígeno-anticuerpo. Entre los primeros métodos para la detección de MCs, se tiene al ensayo por inmunoabsorción ligado a enzimas (ELISA), en el cual requiere dos antígenos (Adda) compitan por un sitio de unión del anticuerpo, en este ensayo se presentan dos métodos; ELISA competitivo directo e indirecto, generando una alta sensibilidad (Tabla 2) y pero presentando debilidades como ser preparación laboriosa, alto costo, baja precisión y fácil perturbación (Heussner et al., 2014).
Las técnicas de inmunoensayo continúan desarrollándose y es así que se presentan otros métodos como biosensores colorimétricos, en donde se utilizan otras tecnologías como la aplicación de vesículas de poliacetileno (VPA). Las VPA son polímeros conjugados que muestran un cambio de color bajo un estímulo de estrés, en donde los cambios de forma de la molécula pueden ser aprovechados durante un reconocimiento que desencadenaría la respuesta. Para la detección de MCs, estas vesículas retendrían la actividad del anticuerpo y al encontrarse con su antígeno (MC-LR), se observaría el cambio de color a una longitud de onda de 540nm. Por lo tanto los procedimientos de reconocimientos son muy simples y ahorran trabajo, la variación crómica de VPA muestra resultados con una buena sensibilidad (Tabla 2) de forma directa y rápida (Xia et al., 2010).
Las tecnologías de fluorescencia se presentan nuevos métodos para la detección de MCs, donde es posible marcar a los anticuerpos o antígenos con pigmentos fluorescentes como la hidrazida de ácido 4-(1-pireno) butanoico (PBH), sin afectar su actividad de los anticuerpos. La florescencia se emite a través de la excitación pulsada aprovechando el tiempo de vida de la florescencia longitudes entre 440-540nm. Estos nuevos métodos mejoran el tiempo de respuesta del análisis, además se integran con nuevas tecnologías o con los métodos tradicionales (Tabla 2) mejorando la sensibilidad, precisión, capacidad de respuesta visual en tiempo real (Liu et al., 2022).
c. Cromatografía acoplada a masas
Los métodos cromatográficos aprovechan las características fisicoquímicas de las MCs, donde el de mayor aplicabilidad es la técnica de cromatografía acoplada a masas (MS), ya que proporciona una solución al problema para la identificación de las distintas variantes de MC, presentando huellas digitales químicas características en los espectros. Además exactitud entre variantes se debe destacar su alta sensibilidad de los diferentes métodos tales cómo, la cromatografía de gases acoplada a espectrometría de masas (GC-MS) y la cromatografía líquida con ionización electro-spray y espectrometría de masas tándem (LC-ESI-MS ) por su capacidad de detección en nanogramos (ng) (Tabla 2) (Kaushik & Balasubramanian, 2013). Las técnicas como cromatografía de alta resolución acoplada a la espectrometría de masas (HPLC-MS) y la cromatografía liquida-espectrometría de masas en tándem (LC-MS/MS), permiten la detección de MCs bajo distintas matrices. A pesar de su capacidad de análisis estos métodos son muy costosos, debido a que requieren estándares de referencia, instrumentación especializada y además su caracterización estructural requiere mucho tiempo, por lo tanto estos métodos solo se hacen aplicables para laboratorios de investigación y no así para análisis de rutina (Kleinteich et al., 2018).
d. Detección genética
La técnica molecular de la reacción en cadena de la polimerasa (PCR) en las últimas décadas ha demostrado ser rápida, rentable y facilita la identificación sin la dependencia de un cultivo. Para la detección de cianobacterias tóxicas se amplifican regiones del ADN genómico, el rRNA 16S y los genes myc (A, B, C, E y G) que están involucrados en la síntesis de MCs. La amplificación de cebadores universales son las secuencias conservadas más apropiadas como objetivos para la identificación por PCR, en este caso se hablamos del gen myc A, después se tiene a los genes específicos (Kesari et al., 2022). Los enfoques basados en PCR permiten su observación (electroforesis), manipulación (clonación) y su secuenciación, además con el avance de la tecnología se desarrollan nuevas técnicas como la PCR digital de gotas (ddPCR) la cual es una presentación avanzada de la qPCR donde una muestra de DNA ambiental (DNAe) se divide en miles de nanogotas antes de someterse amplificación, con resultados más eficientes. Así mismo se tiene la secuenciación de alto rendimiento (HTS) presentando una alta resolución taxonómica en la evaluación de muestras DNAe, permitiendo detectar simultáneamente todas las especies en una muestra ambiental generando datos de la biodiversidad de una comunidad, aunque el HTS está vinculado con el análisis bioinformático siendo una herramienta poderosa para investigación y monitoreo (Feist & Lance, 2021).
Tabla 2 Métodos de detección MCs, aplicadas en fuentes aguas contaminadas.
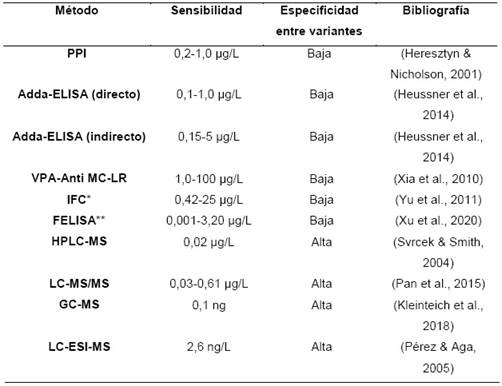
*IFC: Fluoroinmunoensayo de resolución temporal. **FELISA: Inmunoensayo ligado a enzimas de fluorescencia sin marcado de nanopartículas fluorescentes.
Es necesario comprender los mecanismos de la eutrofización ayudaría a prevenir y remediar el exceso uso de nutrientes (Yang et al., 2008). La necesidad de desarrollar sensores activos y en tiempo real permitiría controlar el estado de las fuentes agua además de identificar al agente causante (Jung et al., 2021). Y los recursos que contienen altas concentraciones de P, N, etc., requieren un tratamiento eficaz entonces el desarrollo de estrategias es de importante necesidad, por ejemplo con la aplicación de biocarbón impregnado de con 𝐹𝑒𝐶 𝑙 3 que ha demostrado contener al N y P por su alta porosidad y su capacidad de complejación superficial, precipitación e interacción electrostática (Min et al., 2020).
CONCLUSIÓN
Las MCs tienen el potencial contaminar fuentes de agua como ríos y lagunas, causando daños a la salud humana y a las plantas agrícolas afectando la cadena alimentaria, estas MCs debido a su estructura química cíclica, tienen la capacidad de tolerar distintos factores fisicoquímicos y biológicos, es por ellos que ver alternativas para su degradación es gran importancia. Entre las alternativas estudiadas la degradación bacteriana promete ser la más confiable, por una degradación total de estas cianotoxinas, llegando a péptidos y aminoácidos por un complejo multiproteico modulado por los genes mlr ABCD observadas en bacterias, destacando el género Sphingopyxis por su capacidad de metabolizar MC-LR hasta CO 2 . La degradación biológica es amigable con el ecosistema a diferencia de la fotocatálisis, que aun presenta problemas con rentabilidad y generar sub-productos con probabilidad de toxicidad, aunque los avances en este campo continua viendo alternativas, como utilizar catalizadores como el óxido de titanio, que en distintas matrices podría optimizar la producción de ROS y generar una oxidación completa, siendo así que podría llegar ser un método degradación de importancia en las plantas de tratamiento. Finalmente entre los distintos métodos para la detección de MCs, el de mayor compromiso son los inmunoensayos, debido a su versatilidad de acoplarse a diferentes métodos sean tradiciones o novedosos, mejorando distintos parámetros analíticos, aun así los métodos cromatográficos continúan siendo los más confiables pero debido a sus grandes costos los hacen inviables.
Es innegable la importancia de las MCs en la actualidad y es por ellos que se debe continuar con su investigación, dilucidando rutas de biosíntesis y degradación para comprender su comportamiento bajo distintos panoramas fisicoquímicos y biológicos, además de detectar oportunamente estos metabolitos y evitar su descontrol en distintos cuerpos agua debido a que pueden repercutir en él, medio ambiente, en la economía y la salud general.

