











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
Permalink
INTRODUCCIÓN
La Proteína Quinasa Activada por AMP (AMPK), es una enzima metabólica que actúa como un monitor y regulador central del estado energético celular en eucariotas, por tanto, es responsable de la regulación del metabolismo en respuesta al suministro y demanda de energía. Desde una perspectiva funcional, las condiciones de agotamiento de energía (cuando la proporción entre AMP/ATP se incrementa) causan la activación de la AMPK que conduce a la regulación positiva de procesos catabólicos productores de ATP (como la oxidación de ácidos grasos y la glucólisis) y la correspondiente regulación negativa de procesos anabólicos (como la síntesis de colesterol, lípidos y la gluconeogénesis) para así, restaurar los niveles de ATP y recobrar el balance energético celular (Carling et al.,2011; Hardie, 2008; Oakhill et al., 2012; Steinberg and Kemp, 2009). Dada esta función central en el metabolismo, la AMPK surge como un blanco proteico prometedor para el tratamiento de diferentes enfermedades como la Diabetes Mellitus tipo 2 (DM2), el Síndrome Metabólico (SM) y el Cáncer (Hardie, 2013; Viollet et al., 2007). Sin embargo, la complejidad de esta enzima y la falta de comprensión de su modo de acción molecular han dificultado el desarrollo farmacológico de moléculas activadoras de AMPK como enfoque terapéutico en estos trastornos.
ESTRUCTURA
La AMPK es un heterotrímero de 145 kDa con actividad Serina/Treonina quinasa, formada por una Subunidad Catalítica α y dos Subunidades Reguladoras β y γ (Figura 1). La Subunidad α incluye un Dominio Quinasa N-terminal (KD) o Dominio Catalítico y un Dominio de Interacción de Subunidades (SD) que juega un rol estructural mediando las interacciones con las subunidades β y γ. Entre estas regiones, yace un Bucle que forma un Dominio Auto-Inhibidor (AID) compuesto por un grupo de tres α hélices localizado en la región C-terminal de KD. Adicionalmente, el dominio KD contiene un Bucle de Activación donde se encuentra la αThr172 que es susceptible de ser fosforilada en respuesta a la unión de nucleótidos (AMP). Por otro lado, dos Motivos Reguladores de Interacción de subunidades (αRIM1 y αRIM2) se encuentran localizados en AID y SD, respectivamente, los cuales parecen tener un rol crítico en la regulación alostérica de la AMPK por AMP (Chen et al., 2013; Xiao et al., 2011).
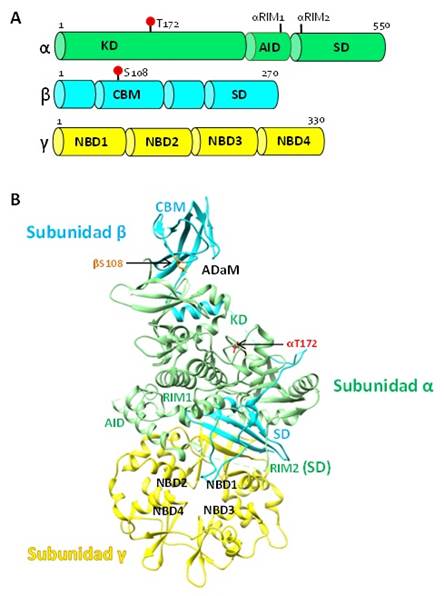
Figura 1 A. Esquema de la AMPK (Extraído y modificado de Chen et al., 2013). B. Estructura trimérica de la AMPK. Subunidad α (verde). Dominio Quinasa (KD) con la αThr172. Dominio de Interacción de Subunidades (SD) con el Motivo Regulador RIM2. Dominio Auto-Inhibidor (AID) con el Motivo Regulador RIM1. Subunidad β (celeste). Módulo de Unión a Carbohidratos (CBM) en N-terminal. Dominio de Interacción de Subunidades (SD) en C-terminal. Subunidad γ (amarillo). Dominios de Unión a Nucleótidos (NBD1-4). El Sitio ADaM entre αKD y βCBM con la βSer108 (PDB: 4ZHX) ((Elaboración propia, Chimera 1.11.2).
La Subunidad β, posee una región N-terminal sin una estructura definida seguida de un Módulo de Unión a Carbohidratos (CBM) formada por hojas plegadas β capaces de interaccionar con el Glucógeno (Polekhina et al., 2005). La región C-terminal actúa como una estructura de andamio con un Dominio de Interacción de Subunidades (SD) propio, que forma una estructura con forma de hoja β extendida entre las Subunidades α y γ constituyendo un núcleo de regulación. Una zona formada por un lóbulo pequeño N-terminal localizado en el KD de la Subunidad α y el CBM de la subunidad β (residuos 76 -156), da origen a una región denominada como el sitio de Unión de Drogas y Metabolitos Alostéricos (ADaM) que parece participar en la activación de la AMPK tras su unión con moléculas pequeñas potencialmente activadoras (Langedorf and Kemp, 2015). El sitio ADaM posee un residuo de βSer108 que generalmente se encuentra fosforilada. Además, la Subunidad β puede sufrir un proceso de α-N-miristoilación en el residuo de βGly2 que facilita su unión reversible con la membrana celular y para la activación de otras quinasas (Oakhill et al., 2010; Steinberg and Kemp, 2009; Warden et al., 2001).
La subunidad γ está constituida por cuatro Motivos en tándem de Cistationina β-Sintasa (CBS1-4) que son módulos estructurales de ~60 aminoácidos (repetidos en tándem) organizados en dos hojas β entrelazadas con dos α hélices circundantes. Un par de módulos CBS forman un dominio “Bateman” el cual posee dos sitios de unión de nucleótidos, uno en cada lado (NBD1/2 y NBD3/4). Los nucleótidos de Adenina como el ATP, ADP y AMP interaccionan dinámicamente con los NBDs de esta subunidad. La activación alostérica de la AMPK por AMP parece involucrar 3 de los 4 NBDs, sin embargo, la unión de AMP con NBD3 puede ser el más importante (Langendorf et al., 2016).
Cada subunidad de la AMPK tiene múltiples isoformas (α1, α2, β1, β2, γ1, γ2, γ3) que se regulan y expresan diferencialmente en todo el organismo. Por ejemplo, las isoformas AMPK-α1 y α2 se distribuyen en el corazón (Kim et al., 2012), mientras que la isoforma AMPK-α2/β2 se expresa casi exclusivamente en el músculo esquelético que es el órgano primario para el almacenamiento y eliminación de la glucosa. La isoforma AMPK-β2 puede dirigir la homeostasis de la glucosa en músculo esquelético por una ruta independiente a la insulina, por tanto, es un importante blanco terapéutico para el tratamiento de enfermedades metabólicas (como la DM2) (Wojtaszewski et al., 2005; Scott et al., 2008).
ACTIVACIÓN ALOSTÉRICA
AMPK detecta el estrés energético traducido en el incremento de AMP. Su activación alostérica involucra la unión de AMP a tres NBDs (NBD1, 3 y 4), lo que desencadena un cambio conformacional y un aumento en la fosforilación del residuo αThr172 por quinasas como LKB1 y Proteína Quinasa 2 dependiente de Calmodulina (CaMKK2). La unión de AMP además, mantiene protegida a la αPThr172 de la acción de fosfatasas, promoviendo un desplazamiento del bucle AID lejos del Dominio Catalítico (αKD). Por el contrario, la unión de ATP promueve la inactivación de AMPK induciendo la defosforilación de α Thr172 (Figura 2). Estudios indican que αRIM2 participa detectando el estado de unión a nucleótidos de la Subunidad γ para la transducción de la señal hacia el Dominio Catalítico (αKD). Otros estudios, señalan que la fosforilación de la βSer108 incrementa la estabilización del sitio ADaM que es importante en la activación de la AMPK. Actualmente, existe un interés importante en el desarrollo de fármacos Activadores de AMPK para su uso terapéutico potencial en el tratamiento de distintas enfermedades metabólicas (DM2, Obesidad, Enfermedades Cardiovasculares, etc) (Hardie and Alessi, 2013).
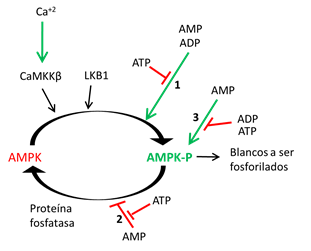
Figura 2 Regulación de la AMPK. La AMPK es activada por el incremento en la proporción AMP o ADP/ATP o el aumento de la concentración de Ca2+. La fosforilación de αThr172 para su activación (AMP-P) es catalizada por al menos dos quinasas: LKB1, la cual parece ser constitutivamente activa y CaMKK2 que solo es activa al incrementarse el Ca2+ intracelular. El aumento de AMP o ADP promueve la activación de la AMPK por tres mecanismos: 1. La unión de AMP/ADP a AMPK produce un cambio conformacional que promueve su fosforilación por quinasas como LKB1. 2. La unión de AMP/ADP ocasiona un cambio conformacional que inhibe su defosforilación por fosfatasas. 3. La unión de AMP (excluyendo el ADP) produce la activación alostérica de la AMPK-P. Todos estos tres efectos son antagonizados por el ATP, permitiendo a la AMPK actuar como un sensor de energía (Extraído y modificado de Hardie y Alessi, 2013).
MOLÉCULAS ACTIVADORAS
La primera droga activadora de la AMPK fue AICAR (ribósido de 5-aminoimidazol-4-carboxamida) (Sullivan et al., 1994; Henin et al., 1995) que administrada en ratones obesos y con resistencia a la Insulina, revirtió muchas de sus anormalidades metabólicas (Song et al., 2002; Buhl et al., 2002). Por otro lado, la Metformina también mostró capacidad activadora de la AMPK in vivo. Estudios con ratones knock-out para la activación de AMPK en el hígado, revelaron que la Metformina perdía su efecto anti-hiperglucémico, sugiriendo que el mayor efecto de la droga ocurre reprimiendo la gluconeogénesis por medio de la activación de la AMPK hepática (Shaw et al., 2005). Otros estudios señalan que algunas saponinas como Damulin A y B, Foenumosida B, Soyasapogenol B y Dioscin aisladas de plantas, disminuyen los factores de transcripción adipogénicos a través de una ruta de señalización que implica la activación de AMPK (Marrelli et al., 2016).
Actualmente, se han identificado pequeñas moléculas activadoras de primera generación como A769662 (Abbott Laboratories), PF-06409577, Salicilato y el Compuesto 991 (Merck Sharp y Dohme Corporation and Metabasis Therapeutics) que se unen al sitio ADaM. Se piensa que la activación de la AMPK por estas moléculas se debe a la estabilización del sitio ADaM debido al incremento en la fosforilación del residuo de βSer108 (Langendorf et al., 2016). Sin embargo, estos activadores de primera generación muestran una selectividad muy fuerte para la isoforma AMPK-β1 (Xiao et al., 2013; Cameron et al., 2016).
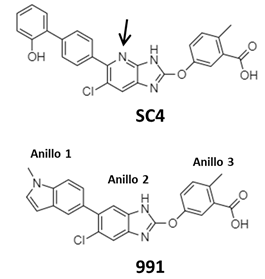
Otro activador alostérico de la AMPK es el ácido 5-(5-hidroxil-isoxazol-3-il)-furan-2-fosfónico (Compuesto 2 o C2) (Metabasis Therapeutics), este fármaco es impermeable para las células, por tanto, se emplea su correspondiente profármaco el di-iso-propil fosfoéster (Compuesto 13 o C13) que estimula preferentemente a la isoforma AMPK-α1 de células eucariotas. AMP y C2 comparten probablemente el mismo sitio de unión y mecanismo de activación debido a que sus efectos alostéricos no son aditivos y la actividad de ambos se pierde en cepas mutantes insensibles a AMP (Kim et al., 2016). Otros estudios pre-clínicos realizados in vivo indican que activadores como PF-739 y MK-8722 mejoran la homeostasis de la glucosa en ratones diabéticos, sin embargo, MK-8722 fue asociado con complicaciones cardiacas tras dosis prolongadas (Myers et al., 2017). Estudios recientes señalan la importancia de la molécula activadora SC4, puesto que tiene una gran selectividad por la isoforma AMPK-β2. Estructuralmente, SC4 comparte con el compuesto 991 el anillo -3(ácido 2-metilbenzoico/ácido o-toluico) y el anillo del núcleo -2(imidazolpirina), pero difiere en la presencia de un átomo de Nitrógeno en la posición 4’ (Figura 3). Debido a su función biológica, SC4 podría servir como modelo para ampliar el repertorio de activadores o agonistas de AMPK-β2 y el desarrollo de fármacos más específicos para tratar enfermedades metabólicas (Ngoei et al., 2018). Sin embargo, la información estructural de cómo estos fármacos activadores se unen y activan a la isoforma AMPK-β2 aún no está bien establecida (Ngoei et al., 2018).
METODOLOGÍA
En este estudio, se realizó un análisis estructural comparativo y descriptivo de diferentes estructuras cristalográficas de la AMPK humana, obtenidas de la base de datos Protein Data Bank (PDB: http://www.rcsb.org/).
PDB 4ZHX: Estructura cristalográfica de la isoforma AMPK-α1 en complejo con el activador A769662 y C2. Esta estructura fue utilizada para el modelamiento de la Figura 1B.
PDB 6B2E: Estructura cristalográfica de la isoforma AMPK-α2β2γ1 humana en complejo con el activador SC4 y AMP.
PDB 6B1U: Estructura cristalográfica de la isoforma AMPK-α2β1γ1 humana en complejo con el activador SC4 y AMP.
PDB 4EAI: Estructura cristalográfica del núcleo de AMPK-β1 (Subunidad γ) humana en complejo con AMP (activo).
PDB 4EAK: Estructura cristalográfica del núcleo de AMPK-β1 (Subunidad γ) humana en complejo con ATP (inactivo).
Todas las estructuras fueron analizadas, evaluadas y modeladas mediante el software Chimera v1.11.2. Las superposiciones y alineamientos estructurales fueron realizados mediante la herramienta Match Maker con una Iteración no mayor a los 2 Å (Pettersen et al., 2004).
RESULTADOS
UNIÓN DE CS4 CON AMPK-β2
El activador sintético SC4 se une en el sitio ADaM de AMPK-β2 formado por la interface entre sus subunidades α y β (dominios α-KD y β2-CBM, respectivamente) (Figura 4A y B). El N4’ forma un puente salino (interacción electrostática) con βD111 y los anillos aromáticos se estabilizan por la interacción con aminoácidos hidrofóbicos como αV11, αL18, αV24, αI46, αF90, y βI115 y βV113 (Figura 4C).
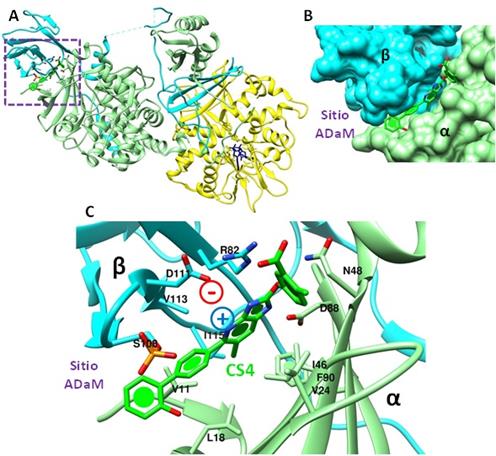
Figura 4 β2-AMPK. A. Estructura heterotrimérica (Subunidad α verde claro, Subunidad β celeste y Subunidad γ amarilla) unida a la molécula activadora CS4 (verde dentro el recuadro morado). B. Densidad electrónica de la superficie de unión de CS4 que corresponde al sitio ADaM formado entre los dominios α-KD (verde) y β2-CBM (celeste). C. Residuos en el sitio ADaM que participan en la interacción entre CS4 y AMPK. N4’ (+) forma una interacción electrostática con βD111 (-). βS108 se encuentra en su estado fosforilado (activo) (PDB: 6B2E) (Elaboración propia, Chimera 1.11.2).
COMPARACIÓN DE ADaM EN AMPK-β1 y β2
En el sitio ADaM de AMPK-β2 se encuentra βD111 que no está conservado entre las diferentes isoformas de AMPK, este aminoácido interacciona específicamente con el Nitrógeno localizado en la posición 4’ en la estructura de CS4 (Véase Figura 3). Por el contrario, en el sitio ADaM de AMPK-β1 se encuentra un βN111 (en lugar de βD111), lo que provoca que su interacción con CS4 sea estructuralmente distinta (Figura 5).
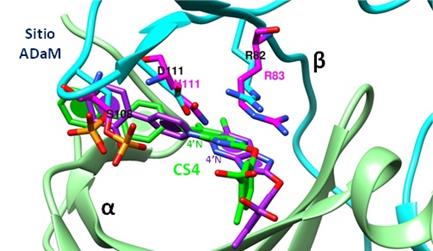
Figura 5 Superposición del sitio ADaM de las isoformas de AMPK-β1 y β2. En la isoforma de AMPK-β2 el residuo βD111 (celeste) interacciona con 4’N de SC4 (verde), mientras que, en su lugar, en la isoforma de AMPK-β1se encuentra el residuo βN111 (magenta) que modifica la configuración de unión de SC4 (morado). El residuo βR82/83 actúa como estabilizador. En ambos casos se observa el cambio en la configuración de los residuos. Por simplicidad se omitió la estructura secundaria de la isoforma de AMPK-β1 (PDB: 6B1U isoforma de AMPK-α2β1γ1 y PDB: 6B2E isoforma de AMPK-α2β2γ1) (Elaboración propia, Chimera 1.11.2).
UNIÓN DE AMP CON NBD1 y 3
El AMP (activador metabólico de AMPK) interacciona con la Subunidad γ (Figura 6A), uniéndose principalmente en los sitios NBD1 y NBD3 e interaccionando con aminoácidos catiónicos (como γR69, γR151, γH150, γK169, γH297, entre otros) que estabilizan la carga negativa de su grupo fosfato (Figura 6B).
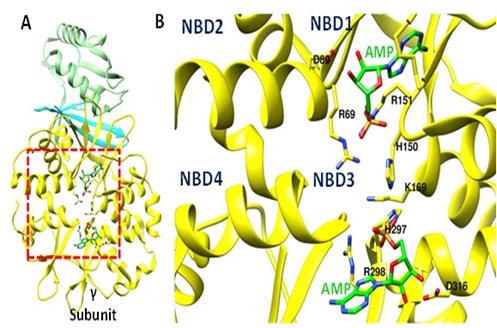
Figura 6 A. Estructura de la Subunidad γ de AMPK-β2 en su estado activo. B. Ampliación del recuadro rojo donde se observa la unión de AMP (verde) en las regiones NBD1 y NBD3. Se indican principalmente los aminoácidos con carga positiva (como γR69, γR151, γH150, γK169, γH297) que estabilizan el grupo fosfato negativo (-3PO4) del AMP. Por simplicidad no se muestra la estructura del AMP unida al sitio NBD4 (PDB: 4EAI) (Elaboración propia, Chimera 1.11.2).
UNIÓN DE ATP CON NBD1 y 3
El ATP (inactivador metabólico de AMPK) compite con el AMP para interaccionar en los sitios NBD1 y NBD3 de la Subunidad γ. La comparación estructural de la disposición de aminoácidos en NBD1 muestra que el ATP produce un cambio conformacional importante en residuos catiónicos como γH150, γR151 y γR69 (Figura 7A y B). Lo mismo ocurre con los residuos γK169 y γH297 de NBD3 (Figura 7C y D).
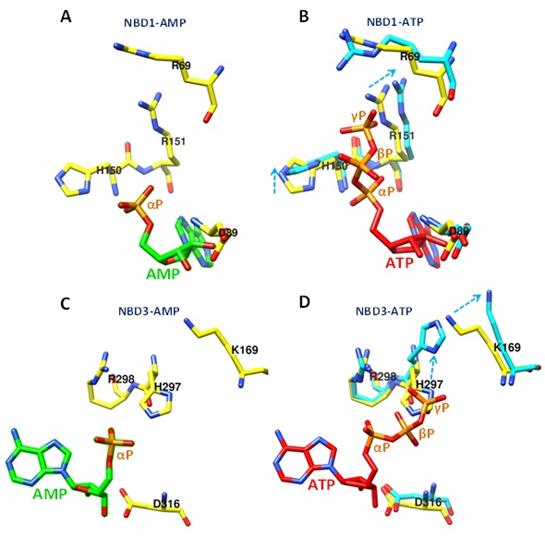
Figura 7 Cambios conformacionales producidos por la unión de ATP en NBD1 y NBD3. La unión de ATP en NBD1 produce el desplazamiento y rotación de los residuos de γH150, γR151 y γR69 (comparación entre A y B). Mientras que en NBD3 produce desplazamientos en los residuos γH297 y γK169 (comparación entre C y D). Los cambios en la posición de los residuos por la unión de ATP se observan en celeste. (AMP verde, ATP rojo). Por simplicidad se omitieron las estructuras secundarias y el resto de aminoácidos. La localización de los dominios NBD1 a 4, fueron realizados de acuerdo a los estudios de Oakhill et al., 2010 (PDB: 4EAI Subunidad γ unida a AMP, PDB: 4EAK Subunidad γ unida a ATP) (Elaboración propia, Chimera 1.11.2).
PAPEL DE NBD3
El cambio conformacional que se produce en la Subunidad γ durante la activación e inactivación de la AMPK, transfiere la señal hacia las otras Subunidades mediante la interacción de γNBD3 con α-RIM2 (Subunidad α) y con un Bucle C-terminal en βSD (Subunidad β), respectivamente (Figura 8A). Ambas (α-RIM2 y el Bucle-βSD), repercuten en la configuración de αAID y la β-Hélice, esta última parece acoplarse al surco pequeño formado por αAID (Figura 8B).
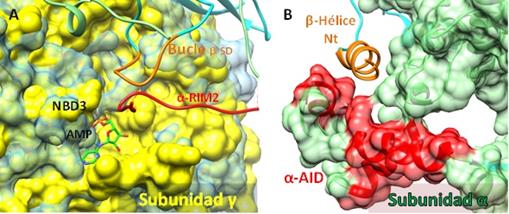
Figura 8 A. Cambios conformacionales de la subunidad γ (densidad electrónica amarilla activa y celeste inactiva). NBD3 interacciona con α-RIM2 (lazo/gancho rojo) y con un Bucle en βSD (anaranjado). B. Una β-Hélice (región N-terminal de la Subunidad β, anaranjado) interacciona con un surco pequeño formado por α-AID (densidad electrónica roja) localizado en la Subunidad α (densidad electrónica verde) (PDB: 6B2E, PDB: 4EAK para la Subunidad γ inactiva) (Elaboración propia, Chimera 1.11.2).
REGIONES INVOLUCRADAS EN LA ACTIVACIÓN DE AMPK-β2
Las principales regiones de la AMPK-β2 se encuentran esquematizadas en la Figura 9. La superposición de las densidades electrónicas de la Subunidad γ de AMPK-β2 señala los cambios conformacionales globales que esta estructura sufre tras su unión con ADP o ATP. Por otro lado, se observan diferentes regiones con forma de bucles o lazos como α-RIM2 perteneciente a la Subunidad α y un Bucle localizado en el Dominio de Interacción de Subunidades (SD) de la Subunidad β, ambos, interaccionan muy próximos al NBD3 de la Subunidad γ. Adicionalmente, se señala el Lóbulo-N que forma parte del Dominio Catalítico (KD) y el α-AID con sus tres hélices (α1-3), ambos pertenecientes a la Subunidad α. Una hélice extra denominada como β-Hélice localizada en la región N-terminal de la Subunidad β, parece interaccionar con α-AID. La estructura de SC4 en el sitio ADaM se encuentra muy próximo al residuo fosforilado βS108. Finalmente, puesto que la estructura corresponde a una AMPK en su estado activo, el residuo αT172 presente en el bucle de activación de la Subunidad α, se encuentra también fosforilado (Figura 9).
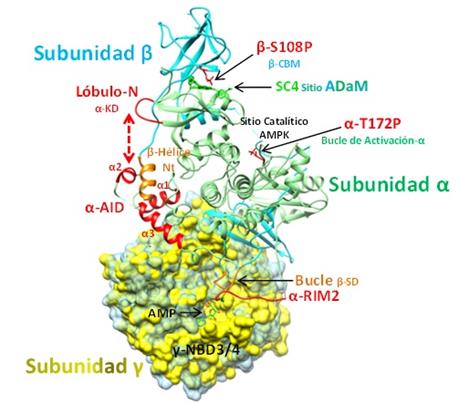
Figura 9 Estructura trimérica de AMPK-β2. Se indican todas las regiones que participan en la dinámica de su activación e inactivación de acuerdo a su unión con moléculas activadoras como SC4 y AMP o inactivadoras como ATP. En la subunidad γ se observa la superposición de su densidad electrónica en estado activo (amarillo) e inactivo (celeste) evidenciando sus cambios conformacionales que tienen repercusión en el resto de la estructura proteica (PDB: 6B2E, PDB: 4EAK para la Subunidad γ inactiva) (Elaboración propia, Chimera 1.11.2).
DISCUSIÓN Y CONCLUSIONES
Debido a que el músculo esquelético se encarga del consumo y almacenamiento de la Glucosa, la acción de moléculas activadoras de la AMPK-β2 es de gran interés clínico para mejorar su homeostasis en sangre (Dasgupta et al., 2012). Estudios recientes indican que el activador SC4 actúa de forma específica sobre AMPK-β2 (que es la isoforma predominante en el músculo esquelético humano) uniéndose al sitio ADaM de manera similar a muchos otros activadores de AMPK (Wojtaszewski et al., 2005; Ngoei et al., 2018). La especificidad de SC4 por AMPK-β2 (EC50 17,2 ± 1,6 nM) se debe principalmente a la interacción electrostática (puente salino) entre el átomo de Nitrógeno 4’ del anillo -2(imidazolpirina) de SC4 y el residuo no conservado β2D111 del sitio ADaM, aunque la sustitución del grupo 2-hidroxifenilo con estructuras basadas en ciclohexeno sustituido con moléculas polares (compuestos MSG010 y MSG011) también mostraron selectividad para AMPK α2β2γ1 (Ovens et al., 2022). Aunque esta interacción podría ser la única responsable de la potencia y especificidad de SC4, el residuo β2R82 del Sitio ADaM también forma una interacción de tipo catiónica-π con el mismo anillo que podría contribuir en la especificidad que tiene este activador. Por tanto, la especificidad del sitio ADaM de la AMPK-β2 por SC4 se debe esencialmente a sus residuos característicos y no así a su estructura global (Gu et al., 2018). A diferencia de otros activadores como A-769662 (que actúan de forma sinérgica con el AMP), estudios realizados en miotubos y hepatocitos de ratón indican que SC4 induce la activación de AMPK-β2 de manera independiente a la proporción celular de AMP/ATP y a los cambios en la fosforilación de αT172 y βS108. Así, SC4 actuaría directamente sobre el sitio ADaM estabilizando la estructura de la AMPK-β2 en una configuración activa (Bultot et al., 2016; Stephenne et al., 2011; Ngoei et al., 2018).
Los resultados obtenidos de la comparación de diferentes estructuras cristalográficas de la AMPK incluyendo las isoformas β1 y β2, sugieren un modelo en la dinámica de su activación en el que participa la Subunidad γ como un sensor de la proporción AMP/ATP. En condiciones de déficit energético (incremento de AMP/ATP), el AMP primero interacciona con NBD1 puesto que este dominio une AMP con una mayor afinidad (x10) en comparación con el ATP. La unión de AMP-NBD1 produce cambios conformacionales que luego son transmitidos al resto de la Subunidad γ, principalmente a NBD3 que es el que tiene menor afinidad por AMP. La unión AMP-NBD3 también podría afectar la interacción de NBD4 por AMP, por consiguiente, es muy probable que la activación alostérica de AMPK dependa en primera instancia de la identidad del nucleótido que ocupa NBD1 (Kurumbail and Calabrese, 2016). De esta manera y en concordancia con lo reportado por Oakhill et al. (2010), se produce una “conversación cruzada” entre los NBDs que conlleva a un cambio conformacional integral de la Subunidad γ. Esta señal de activación a través de cambios conformacionales producidos por la unión de AMP en la Subunidad γ tiene que ser posteriormente transferida al resto de Subunidades (γ y β) de la AMPK.
El segmento regulador con forma de gancho α-RIM2 (365 - 371) de la Subunidad α podría contactar directamente con NBD3 (detectando el estado de unión a AMP o ATP) (Xiao et al., 2011; Chen et al., 2013) y comunicando así, la señal de activación o inactivación al Dominio Catalítico (αKD). Si la señal es de activación, α-RIM2 transmite la señal hacia el Bucle de Activación de αKD (mediante los segmentos regulatorios flexibles presentes en la Subunidad α) protegiendo así el residuo αPT172 de la acción de fosfatasas inactivadoras (Iñiguez y Coutiño, 2008 y Kurumbail and Calabrese, 2016). Adicionalmente, la señal de activación detectada por α-RIM2, también es transmitida hacia la región trihelicoidal α-AID que experimenta una rotación importante de tal manera que se aleja del Lóbulo-N localizado en el Dominio Catalítico (α-KD) de la AMPK y se acerca a la Subunidad γ (Figura 9). De esta forma, el Lóbulo-N puede mantenerse en una configuración extendida permitiendo que la AMPK se mantenga activa.
El Bucle-βSD también interacciona con la región NBD3 de la Subunidad γ, enviando la señal de activación por AMP hacia una β-Hélice corta (en la región N-terminal de la Subunidad β) que comprende los residuos βP60 a βS70. Esta β-Hélice se ajusta a un surco poco profundo y abierto formado por α-AID. La interacción de la β-Hélice parece mantener a α-AID en una configuración alejada del Dominio Catalítico (α-KD) de la AMPK. Por el contrario, cuando la AMPK se encuentra en estado inactivo, α-AID se acerca e interacciona con el Lóbulo-N promoviendo la apertura del Dominio Catalítico haciéndola más susceptible a la acción de fosfatasas inactivadoras.
NBD3 es el sitio de unión a nucleótidos más débil en la Subunidad γ, sin embargo, es extremadamente sensible a los cambios en los niveles de nucleótidos y esto permite que AMPK responda rápidamente a las variaciones del estado energético de la célula. Por el contrario, moléculas activadoras como SC4 interaccionan con AMPK en el sitio AdaM independientemente del estado de fosforilación de T172, S108 y del estado de unión de los NBDs en la Subunidad γ. Actúan directamente promoviendo un cambio conformacional activo y estabilizando esta configuración con un Dominio Catalítico activo. El estudio de este modelo de activación es muy importante para poder diseñar, extraer e identificar compuestos activos de fuentes naturales que tengan propiedades de especificidad y potencia similar a SC4 u otros activadores sintéticos, para poder emplearlos como tratamientos alternativos de enfermedades metabólicas como la Obesidad y DM2 que afectan enormemente a nuestra población.

