









 texto em
texto em  Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO  uBio
uBio 
 Permalink
PermalinkJournal of the Selva Andina Biosphere
versão impressa ISSN 2308-3867versão On-line ISSN 2308-3859
J. Selva Andina Biosph. vol.10 no.2 La Paz 2022 Epub 01-Nov-2022
https://doi.org/10.36610/j.jsab.2022.100200068
Comunicación Corta
Variation of herbivory damage on potato (Solanum tuberosum ssp. andigena) leaves in different degrees of proximity to native vegetation in Combaya, La Paz

1Universidad Mayor de San Andrés. Institute of Ecology. Botanical Garden of La Paz. Casilla 10077-Correo Central. La Paz, Plurinational State of Bolivia.
Las tasas de herbivoría elevadas están relacionadas con la aparición de nuevas plagas afectando a las plantas de interés comercial, como la papa (Solanum tuberosum). Si bien el interés de los agricultores es conocer a los principales depredadores de tubérculos, también es necesario entender los patrones de herbivoría en otras partes de la planta. En sistemas agrícolas la estructura del paisaje está determinada por los parches de vegetación, las parcelas de cultivo y el desplazamiento de la fauna asociada. Este estudio evaluó el daño por herbivoría en hojas de papa en función de la cercanía a parches de vegetación nativa en el Municipio de Combaya (Departamento La Paz, Bolivia). Se midió el porcentaje de daño foliar, el nivel de daño, el área foliar y la riqueza de herbívoros en cinco cultivos de papa. Los resultados revelaron que el porcentaje de herbivoría en cultivos cerca de la vegetación es significativamente superior al de los cultivos alejados (ANOVA P<0.00023729, F=9.3046). Los cultivos cerca de la vegetación mostraron en promedio 8.266±0.859 % de herbivoría mientras que los cultivos alejados 5.909±0.585 %. En total se capturaron 5 morfo-especies de herbívoros asociados a los cultivos de papa: 3 en cultivos cercanos y 2 en los alejados de la vegetación. La vegetación nativa juega un rol importante porque permitiendo que la fauna aproveche los recursos de todo el paisaje.
Palabras clave: Paisaje; herbivoría; plagas; tubérculos; vegetación; cultivos
High herbivory rates are related to the emergence of new pests and are a problem for plants of commercial interest, such as potato (Solanum tuberosum). While the interest of farmers is to know the main tuber predators, it is also necessary to understand herbivory patterns in other parts of the plant. In agricultural systems, landscape structure is determinated by vegetation patches, crops and the movement of associated fauna. This study evaluated herbivory damage on potato leaves as a function of proximity to patches of native vegetation in the municipality of Combaya (La Paz, Bolivia). The percentage of leaf damage, damage level, leaf area and herbivore richness were measured in five potato crops. The results showed that close vegetation crops have significantly higher herbivory than far away ones (ANOVA P<0.00023729, F=9.3046). Close vegetation crops showed 8.266±0.859 % herbivory, while are away ones 5.909±0.585 %. In total, five morpho-species of herbivores were found associated with potato crops: 3 in crops near and 2 in crops far from the vegetation. The native vegetation plays an important role because it allows the fauna to take advantage of the resources of the entire landscape.
Keywords: Landscape; herbivory; plagues; tubers; vegetation; crops
Introduction
High herbivory rates cause two types of effects, i) at the physiological level, it is common for the plant to benefit from commensurate growth and increased production of defense chemicals1,2 and ii) at the ecological level, it is related to events such as the appearance of pests2. Although plants have defense mechanisms against herbivores3, some factors, such as nutrient and water limitation, alter their ability to respond to predation4,5 and in cultivated areas commercial plants are put at risk.
Crop pest problems are related to the structure of the landscape matrix6, as patches of native vegetation interact with crops by displacing associated fauna7,8. On the one hand, specialist fauna prefers to stay in native vegetation, while generalist fauna does not mind expanding into disturbed areas, such as arable areas9. The variety of resources and the lack of competition make agricultural systems diversify areas for different animal groups, including herbivores10. Therefore, the distance between vegetation and crops is a factor that can determine the composition of herbivores in crops and at the same time increase herbivory rates.
In most plants, herbivory rates are dependent on leaf area (LA) and leaf age11. It is understood that small and young leaves are more susceptible to predation, although the reasons are diverse, and there is no consensus among authors12. We know that the isoflavonoids and glycoalkaloids present in potato leaves are harmful to some herbivores3 and some studies indicate that the concentrations of these substances fluctuate with the age and size of the leaves12. They occur in lower concentrations when leaves are very young11. Similarly, small leaves have lower concentrations of tannins making them more susceptible to predation by herbivores11.
In potato crops in Bolivia, problems were reported with herbivores such as the brown cicada (Russelli-ana solanicola), green cicada (Empoasca spp.), and nematodes of the genera Nacobbus spp. and Globodera spp.13 that mainly attack tubers. Whereas records of herbivores attacking leaves are scarce, an example is the black leaf beetle (Epicauta spp.) which was a pest in lowland areas of the country14,15. Although the main concern of farmers is to avoid the proliferation of pests in tubers, leaf herbivory also affects the vitality of the plant16, altering the patterns of photosynthesis, reproduction, and the integrity of the individuals16. Previous studies have shown that there is a negative relationship between tuber predation and foliar herbivory17. However, leaf herbivory has been scarcely studied in potatoes and the herbivores that affect it are not clearly known.
The potato (S. tuberosum spp. andigena) is a cultivated species of the Solanaceae family with a wide distribution in South America, which has been recorded between 1500 and 4300 m altitude18, and the same authors place Bolivia and Peru among the countries with the most cultivable varieties of this species19. The center origin and domestication of potato is mentioned as the center of origin and domestication of the potato in the highlands of Perú19,20, from where it spread, and diversified its cultivation and consumption. In Bolivia, the first potato domestication sites were established around Lake Titicaca19,21,22. More than 500 potato varieties have been recorded in Bolivia, and the Alq'a imilla variety is widely used in the Altiplano areas of La Paz due to its adaptation to altitudes above 3000 m23.
Under these concepts, the objectives of this work were: i) to determine the variation of damage by foliar herbivory in potato crops in relation to their proximity to remnants of native vegetation, ii) to establish if there is a relationship between herbivory damage and leaf size and iii) to identify the herbivores present in potato crops.
Materials and methods
Study Area. This study was conducted between October 2017 and March 2018 in the municipality of Combaya, located in the Larecaja province, northeast of the department of La Paz, Bolivia. It is located at 3275 m altitude with an average annual temperature of 16° C and an average rainfall of 750-1200 mm/year. The natural vegetation is shrubby and herbaceous, with the families Asteraceae, Loasaceae, Calceolariceae, Scrophulariacea, Lamiaceae, Oro-banchaceae, and Fabaceae (Figure 1). Agriculture is practiced in the area with potatoes, vegetables, maize, green peas, and green beans.

Figure 1 Vegetation in Combaya municipality, a) Genus Alonsoa, b) Genus Baccharis c) Genus Achyrocline, d) Genus Calceolaria
Distance between crops and vegetation remnants. The distance between crops and vegetation remnants was measured over a range of 100-1000 m (Figure 2). We considered the degree of proximity to native vegetation as follows: i) crops close to vegetation are those located less than 200 m and ii) crops far from vegetation are those located more than 800 m24. For each crop we took into account the following data: number of plants/m2, total plot area, slope, and altitude. These data were taken using a Garmin GPS (Table 1).
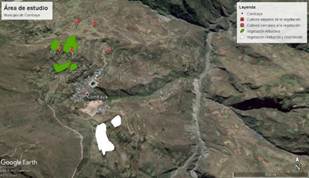
Figure 2 Study area in Combaya municipality. The remnants of shrubby vegetation are marked in green and the remnants of graminoids are marked in white. The crops P1, P2 and P3 were considered distant from the vegetation and P4 and P5, as near ones

Crop sampling. In each crop an "X" sampling was followed, marking the first 3 meters from the first plant to differentiate the edge from the center of the plot (Figure 3). On each plant, a branch was collected at random and 3 leaves were selected for evaluation. The leaves from each branch were stored and labeled in zip lock bags and transported to the offices of the La Paz Botanical Garden. A system was then set up with a universal stand 15 cm from the sample and a 2 cm mark as a reference for leaf size. All samples were photographed from the underside and evaluated with Image J software.
Table 1 Crop data set
| Crop | Distance to vegetation patches (m) | Area (m2) | Coordinates | Altitude (m.a.s.l.) | #Plants/m2 |
|---|---|---|---|---|---|
| Near 1 | 180 | 142 | 15°48’08.81’’ 68°45’25.72’’ | 3376 | 10 |
| Near 2 | 150 | 245 | 15°48’08.71’’ 68°45’21.88’’ | 3360 | 9 |
| Distant 1 | 841 | 577 | 15°48’06.06’’ 68°45’10.80’’ | 3219 | 10 |
| Distant 2 | 800 | 2.159 | 15°47’57.05’’ 68°45’16.37’’ | 3228 | 12 |
| Distant 3 | 720 | 494 | 15°47’56.12’’ 68°45’22.62’’ | 3224 | 11 |
Estimation of leaf damage. For each leaf collected, the leaf area in mm2 LA and the percentage of damaged LA were evaluated. To calculate the percentage of the area lost, the following formula was used.
The lost area is the sum of the partial areas of each leaf space with some kind of damage. LA = total leaf area, all measured in mm225.
Holes inside the leaf, incomplete margins, necrosis marks, galls, and fungal stains were considered damaged LA. The level of damage was categorized as follows26:
Relationship between herbivory damage and leaf size. To determine if there is a relationship between herbivory damage and leaf size, the LA was compared with the percentage of herbivory2.
Herbivore sampling in potato crops. Twenty-five pitfall traps with 70 % alcohol were randomly set in each potato crop. The traps remained active for 24 h, after which they were closed. Direct captures were also made from the plants27 and invertebrates were identified by morphospecies with the help of researchers from the Bolivian Fauna Collection. For the classification of invertebrates, we took into account mouthparts, types of antennae, and types of wing venation28. Individuals were described using the 30x magnification of an Olympus stereo-microscope, and a scale of 0.5 cm was used to estimate the size of the individuals.
Statistical analysis. A total of 698 leaves were analyzed within the 5 cultures. Three replicates were used for the distant crops, each with 151, 111, and 103 leaves evaluated. On the other hand, for the crops close to the vegetation, we took two replicates, each with 173 and 160 leaves. To evaluate the percentage of herbivory, a two-factor ANOVA test was used. As the original data did not meet the assumption of normality, a log-natural rate (ln) transformation was performed. To compare herbivory damage with leaf size, a simple linear regression was used, considering herbivory percentage as the dependent variable and leaf size as the independent variable. All analyses were performed in R software29.
Results
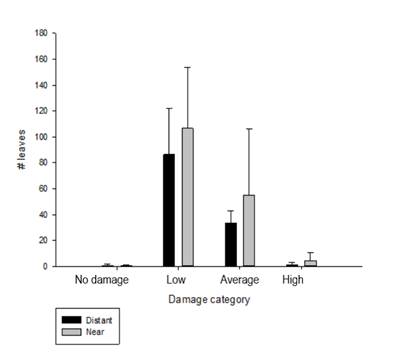
Leaf herbivory on potato. 67 % of the total number of leaves studied (472 leaves) have less than 5 % of their surfaces damaged (Figure 4). 30 % (211 leaves) have up to 20 % of their surfaces affected. Finally, leaves without any damage are 0.42 % (3 leaves), and leave with high degrees of herbivory are 2 % (12 leaves) (Figure 4).
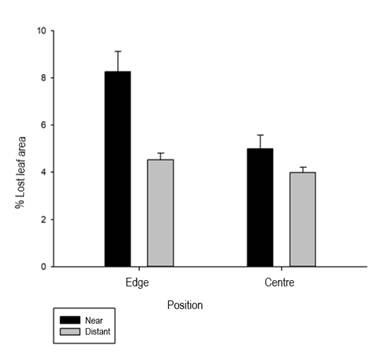
A significant difference was observed between crops near and far from the vegetation patches (ANOVA test, P<0.00023729, F = 9.3046) (Table 2). Crops located less than 200 m from the vegetation suffered more damage than crops more than 800 m away (Figure 5, Table 2).
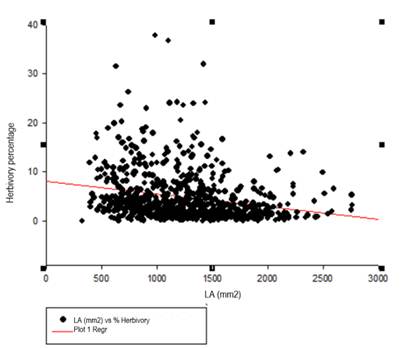
Relationship between leaf size and herbivory damage. According to Figure 6, we notice that smaller leaves are more damaged. However, simple linear regression analysis shows no correlation between the two variables (R=0.22) (Table 3).
Herbivore richness. Five herbivore morph species were captured: green cicada (family Cicadellidae), brown cicada (family Cicadellidae), green grasshopper (family Acrididae), slug (order Gasteropoda), and weevil (family Curculionidae) (Figure 7, Table 4).
Discussion
The percentage of LA lost is higher in crops near native vegetation (8.266±0.859 %), than in remote ones (5.909±0.585 %) (Figures 4 and 5). This is because these crops are more exposed to herbivores moving from the vegetation patches6. Similarly, we know that many herbivores are opportunistic and readily move into crops when they are present in the landscape30, increasing the percentage of herbivory in crops near vegetation. This background leads us to believe that the more heterogeneous landscapes are, the more habitats they provide for various invertebrate groups, and potato crops are exploitable resources for herbivory.
Tabla 3 Regresión lineal
| Df | SS | MS | F | P | |
|---|---|---|---|---|---|
| Regression | 1 | 979.152 | 979.152 | 38.328 | <.001 |
| Residuals | 696 | 17780.494 | 25.547 | ||
| Total | 696 | 18759.647 | 26.915 |
R = .228, R2 = .0522
Table 4 The four herbivores morpho-species present in the studied crops
| Family/Class | Morfo-specie | Near vegetation | Distant vegetation |
|---|---|---|---|
| Cicadellideae verde | Cigarra verde | X | X |
| Cicadellidae café | Cigarra café | X | X |
| Gasteropoda | Babosa | X | X |
| Acrididae | Saltamontes verde | X | _ |
| Curculionidae | Gorgojo | X | _ |

Figure 7 Herbivores in potato crops. a) Grasshoppers, Family Acrididae. b) and c) Cicadas, Family Cicadellidae. d) Slug, Gastropod
The percentage of LA lost in crops close to the vegetation can be explained by the richness of herbivores recorded: 5 morpho-species in nearby crops (green cicada, brown cicada, slug, green grasshopper, weevil) and 3 distant crops (green cicada, brown cicada, and slug) (Figure 7, Table 4). Although this is not a relevant difference, there are records of crop damage by populations of leafhoppers (Acrididae), weevils (Curculionidae), and cicadas (Cicadellidae)14,15,31. Mainly because they are generalist insects that adapt easily to different environmental conditions, they can move between crops and vegetation patches9. Also, several weevil species have been known to cause damage to potato crops14,32. The best known is Premnotrypes sp. which has a preference for tubers14,32. As for species that prey on leaves and other parts of the plant, we have potato moths, potato leafminers, shoot fly, potato flea moths, etc11. In this regard, the present study recorded 5 morpho-species of herbivores (Figure 7, Table 4) that have not been recorded before in potato crops.
By linear regression analysis, we note that leaf size is not related to the percentage of herbivory (Table 3) in the crops studied. However, there is a tendency for smaller leaves to suffer more herbivory damage (Figure 6). These leaves have less surface area and therefore damage is more visible, but it is also known that tannins and isoflavonoids fluctuate with the size and age of the leaves, being present in higher concentrations in larger leaves11,12,33. This phenomenon was observed in perennial plants34 and would be worth exploring further in Andean crops.
It is concluded that the closer the crops are to the vegetation patches, the higher the herbivory rates in potato crops because the native vegetation harbors fauna with the ability to move. In this study, 5 herbivore morpho-species were recorded, all of which were found in crops close to native vegetation, while only 3 were found in remote crops. It is clear that the closer to the vegetation, the greater the richness of herbivores and the higher the percentage of herbivory. However, further experimental studies are needed to better understand herbivore preferences for different habitat types in agroecosystems.
REFERENCES
1. Garcia LC, Eubanks MD. Overcompensation for insect herbivory: a review and meta-analysis of the evidence. Ecology 2019;100(3):e02585. DOI: https://doi.org/10.1002/ecy.2585 [ Links ]
2. Ito K, Sakai S. Optimal defense strategy against herbivory in plants: conditions selecting for induced defense, constitutive defense, and no-defense. J Theor Biol 2009;260(3):453-9. DOI: https://doi.org/10.1016/j.jtbi.2009.07.002 [ Links ]
3. Bernal JS, Medina RF. Agriculture sows pests: how crop domestication, host shifts, and agricultural intensification can create insect pests from herbivores. Curr Opin Insect Sci 2018;26:76-81. DOI: https://doi.org/10.1016/j.cois.2018.01.008 [ Links ]
4. Friedman M. Analysis of biologically active compounds in potatoes (Solanum tuberosum), tomatoes (Lycopersicon esculentum), and jimson weed (Datura stramonium) seeds. J Chromatogr A 2004;1054(1-2):143-55. DOI: https://doi.org/10.1016/j.chroma.2004 [ Links ]
5. Cornelissen T. Climate change and its effects on terrestrial insects and herbivory patterns. Neotrop Entomol 2011;40(2):155-63. DOI: https://doi.org/10.1590/s1519-566x2011000200001 [ Links ]
6. Schowalter TD. Herbivory. In: Schowalter, editor. Insect Ecology: An Ecosystem Approach. Ámsterdam: Elsevier BV; 2022. p. 567-622. DOI: https://doi.org/10.1016/B978-0-323-85673-7.00006-X [ Links ]
7. Fahring L. Effects of habitat fragmentation on biodiversity. Annu Rev Ecol Evol Syst 2003;34: 487-515. DOI: https://doi.org/10.1146/annurevecolsys.34.011802.132419 [ Links ]
8. Mazzi D, Dorn S. Movement of insect pests in agricultural landscapes. Ann Appl Biol 2012;160(2): 97-113. DOI: https://doi.org/10.1111/j.1744-7348.2012.00533.x [ Links ]
9. Aligner A, Raymond L, Deconchat M, Menozzi P, Monteil C, Sarthou JP, et al. The effect of semi-natural habitats on aphids and their natural enemies across spatial and temporal scales. Biol Control 2014;77:76-82. DOI: https://doi.org/10.1016/j.biocontrol.2014.06.006 [ Links ]
10. Yamamoto S, Uchida K. A generalist herbivore requires a wide array of plant species to maintain its populations. Biol Conserv 2018;228:167-74. DOI: https://doi.org/10.1016/j.biocon.2018.10.018 [ Links ]
11. Coley P. Effects of leaf age and plant life history patterns on herbivory. Nature 1980;284:545-6. DOI: https://doi.org/10.1038/284545a0 [ Links ]
12. Medinaceli A, Miranda-Avilés F, Flores-Saldaña NP, Gutierrez-Calucho E. Herbivoría en relación al tamaño de la planta y a las diferencias de exposición de Pilea sp. (Urticaceae) en la Estación Biológica Tunquini, Cotapata, La Paz - Bolivia. Ecol Boliv 2004;39(2):4-8. [ Links ]
13. Nicholls Estrada CI. Control biológico de insectos: un enfoque agroecológico [Internet]. Medellín: Editorial Universidad de Antioquia; 2008 [citado 2 de mayo de 2021]. 294 p. Recuperado a partir de: https://archive.foodfirst.org/wp-content/uploads/2016/01/Control-biologico-de-insectos-un-enfoque-agroecolgico.pdf [ Links ]
14. Franco J, Ramos J, Oros R, Main G, Ortuño N. Pérdidas económicas causadas por Nacobbus aberrans y Globodera spp. en el Cultivo de la Papa en Bolivia. Rev Latinoam Papa 1999;11:40-66. DOI: https://doi.org/10.37066/ralap.v11i1.95 [ Links ]
15. Aguilera J, Almanza J, Álvarez V, Barea O, Bejarano C, Bonifacio A, et al. Compendio de enfermedades, insectos, nematodos y factores abióticos que afectan el cultivo de papa en Bolivia. Gandarillas A, Ortuño N, editors [Internet]. Cochabamba: Fundación PROINPA; 2009. [citado 2 de mayo de 2020]. 182 p. Recuperado a partir de: https://www.researchgate.net/publication/345385397_COmpendio_de_las_enfermedades_insectos_nematodos_y_factores_abioticos_que_afectan_en_el_cultivo_de_la_papa_en_Bolivia [ Links ]
16. Granados-Sánchez D, Ruiz-Puga P, Barrera-Escorcia H. Ecología de la herbivoría. Rev Chapingo Ser Cienc For Ambient 2008;14(1):51-63. [ Links ]
17. Horgan FG, Quiring DT, Lagnaoui A, Pelletier Y. Trade-off between foliage and tuber resistance to Phthorimaea operculella in wild potatoes. Entomol Exp Appl 2009;131(9):130-7. DOI: https://doi.org/10.1111/j.1570-7458.2009.00842.x [ Links ]
18. Valoy M, Ordano M, Bernacki F, Palacio FX, López-Acosta JC, Varela O. Patrones de herbivoría en Vassobia breviflora (Solanaceae): variación en el daño foliar y selección natural mediada por herbívoros. Rev Biol Trop 2018;66(4):1683-700. DOI: http://doi.org/10.15517/rbt.v66i4.31869 [ Links ]
19. Tapia ME, Fries AM. Guía de campo de los cultivos andinos [Internet]. Lima: Asociación Nacional de Productores Ecológicos del Perú; 2007 [citado 22 de octubre de 2020]. 222 p. Recuperado a partir de: https://www.fao.org/3/ai185s/ai185s.pdf [ Links ]
20. Morales Garzón FJ. Sociedades precolombinas asociadas a la domesticación y cultivo de la papa (Solanum tuberosum) en Sudamérica. Rev Latinoam Papa 2007;14(1):1-9. DOI: https://doi.org/10.37066/ralap.v14i1.139 [ Links ]
21. Rodríguez LE. Origen y evolución de la papa cultivada: una revisión. Agron Colomb 2010;28(1):9-17. [ Links ]
22. Bonavia D. La papa: apuntes sobre sus orígenes y su domesticación. J Soc Am 1993;79:173-87. DOI: https://doi.org/10.4000/BOOKS.IFEA.5680 [ Links ]
23. Bradshaw JE, Ramsay G. Potato origin and production. In: Singh J, Kaur L, editors. Advances in potato chemistry and technology. New York: Academic Press is an imprint of Elsevier; 2009. p. 1-26. DOI: https://doi.org/10.1016/B978-0-12-374349-7.00001-5 [ Links ]
24. Tscharntke T, Karp DS, Chaplin-Kramer R, Batáry P, DeClerck F, Gratton C, et al. When natural habitat fails to enhance biological pest control - five hypotheses. Biol Conserv 2016;204(Pt B):449-58. DOI: https://doi.org/10.1016/j.biocon.2016.10.001 [ Links ]
25. Ugarte ML, Iriarte V. Papas Bolivianas: Catalogo de cien variedades nativas [Internet]. Cochabamba: Fundación PROINPA; 2006 [citado 22 de octubre de 2020]. 113 p. Recuperado a partir de: https://es.scribd.com/document/407918901/Catalogo-de-cien-variedades-nativas-de-papas-bolivianas-pdf [ Links ]
26. Pérez-Harguindeguy N, Díaz S, Garnier E, Lavorel S, Poorter H, Jaureguiberry P, et al. New handbook for standardised measurement of plant functional traits worldwide. Aust J Bot 2013;61 (3):167-234. DOI: https://doi.org/10.1071/BT122 25 [ Links ]
27. Christie FJ, Hochuli DF. Elevated levels of herbivory in urban landscapes: are declines in tree health more than an edge effect. Ecol Soc 2005;10(1):10. DOI: https://doi.org/10.5751/ES-00704-100110 [ Links ]
28. Delvare G, Aberlenec HP, Michel B, Figueroa A. Los insectos de áfrica y de América tropical claves para la identificación de las principales familias [Internet]. Montpellier: Campus International de Baillarguet - CSIRO; 2002 [citado 22 de octubre de 2020]. 257 p. Recuperado a partir de: http://aberlentomo.fr/02_delvaberl_1989_pdf/delvaberl_espana.pdf [ Links ]
29. Llisterri J. R, A language and environment for statistical computing [Internet]. The R Foundation. 2021 [citado 3 de octubre de 2020]. Recuperado a partir de: https://joaquimllisterri.cat/phonetics/fon_R/R.html [ Links ]
30. Fernández García I, Favila Castillo ME, López Iborra G. Composición, riqueza y abundancia de coleópteros (coleóptera) asociados a bosques semideciduos y vegetaciones ruderales de la Sierra del Rosario, Cuba. Boletín de la Sociedad Entomológica Aragonesa 2014;54:329-39. [ Links ]
31. Andow D. Vegetational diversity and arthropod population response. Annu Rev Entomol 1991; 36(1):561-86. DOI: https://doi.org/10.1146/annurev.en.36.010191.003021 [ Links ]
32. Pérez W, Forbes G. Guía de identificación de plagas que afectan a la papa en la zona andina [Internet]. Lima: Centro Internacional de la Papa; 2011 [citado 2 de octubre de 2020]. 44 p. Recuperado a partir de: https://www.fao.org/3/as407s/as407s.pdf [ Links ]
33. Maoela MA, Esler KJ, Roets F, Jacobs SM. Physiological responses to folivory and phytopathogens in a riparian tree, Brabejum stellatifolium, native to the fynbos biome of South Africa. Afr J Ecol 2018;56(3):477-87. DOI: https://doi.org/10.1111/aje.12481 [ Links ]
34. Kursar TA, Coley PD. Convergence in defense syndromes of young leaves in tropical rainforests. Biochem Syst Ecol 2003;31(8):929-49. DOI: https://doi.org/10.1016/S0305-1978(03)00087-5 [ Links ]
Source of financing Project "phytochemical research on leaves and fruits of potatoes for products with added value of Combaya-La Paz" of the institute of physical research Undergraduate grant from the institute of ecology.
Acknowledgments This study was carried out thanks to the funding projects "Phytochemical research on leaves and fruits of potatoes for products with added value in Combaya-La Paz" of the Physical Research Institute and to the undergraduate scholarship granted by the Ecology Institute. Gabinete's work was carried out with the support of the La Paz Botanical Garden. Thanks to Juan Carlos Peñaranda, Camila Ramallo and Sofia Lottersberger, for their support in data collection in the field. Also, to Miguel Limachi, associate researcher of the Bolivian Fauna Collection for his support in the identification of soil macrospecies.
Ethical considerations All field data was collected with the permission of the corresponding authorities of the municipality of combaya, as well as with the knowledge of the inhabitants and owners of the potato plots.
Authors' contributionEmili Antonia Jimenez; data collection in the field, organization, systematization and analysis of data, writing of the article. Esther Valenzuela Celis; review of data and article.
Editor's Note: Journal of the Selva Andina Biosphere (JSAB). All statements expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, editors and reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Received: April 01, 2021; Revised: September 01, 2022; Accepted: October 01, 2022
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons