











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
Permalink
La empatía integra un conjunto de constructos que suponen principalmente procesos que vinculan cognitiva y emocionalmente a un individuo con respecto a otros similares. Darwin (1871), advirtiendo la consistencia de la preocupación del individuo por su congénere y del comportamiento de ayuda sugirió que la conducta empática prosocial es una tendencia natural cuyo propósito es la preservación de la especie humana. Mehrabian & Epstein (1972), afirmaron que la empatía es una respuesta emocional vicaria que se experimenta a través de diferentes mecanismos ante las experiencias emocionales ajenas. Años más tarde, Davis (1980), definió el constructo con un mayor grado de complejidad, trascendiendo el planteamiento inicialmente dual (cognitivo-emocional) con que fuera definido años atrás y proponiendo una concepción multidimensional del término, que definía la empatía como “un conjunto de constructos que incluyen los procesos de ponerse en el lugar del otro y respuestas tanto afectivas como no afectivas que resultan de esos procesos” (pp.1-17).
El enfoque conceptual con el que se definió empatía la presupuso indiscutiblemente como un atributo esencialmente humano, capaz de facilitar actitudes altruistas entre miembros de nuestra especie (Hoffman, 1981). La condición humana de la empatía suponía que sus componentes definitorios referían procesos típicamente humanos tales como “percibir”, “interpretar”, “sentir”, “emocionarse”, “apenarse”, etc., que no podían fácilmente atribuirse a otras especies. De ahí que la expresión “empatía animal” era una expresión emergente de un modelo construido para entender un fenómeno humano, que se aplicaba inapropiadamente para explicar por qué un animal menor, ayudaba espontáneamente a otro cuando éste se encontraba en situación de necesidad (Gamboa, et al 2017; Gamboa & Reynoso, 2016; Silva & de Sousa, 2011). Así, no es raro encontrar en la literatura atribuciones cognitivas y/o emocionales en las descripciones del comportamiento de ayuda en especies no humanas. Por ejemplo, Mejía et al., (2009), han sugerido, que en el caso de los primates, roedores y humanos, entre otras especies, la expresión empática debe iniciarse con una conexión cognitiva que informa acerca de una situación emocional, seguida de la preocupación por el otro y de una respuesta motora que involucre un compromiso con la solución actual o potencial del organismo que sufre. En estudios previos realizados por Ben-Ami Bartal, et al, 2011, 2014, 2016; Horner et al, 2011; Mason 2014; Monteau-Arias et al., (2019) Roth & Flores (2019), con roedores y otros animales, se demostró que el alivio de un compañero en situación de necesidad, requería de un procesamiento afectivo y de una conexión social y cognitiva.
El estudio de la intercurrencia de los factores afectivos en la empatía orilló a explorar, el efecto de los bloqueadores emocionales (farmacológicos como las benzodiacepinas) en la manifestación de la conducta de prestar ayuda al prójimo. Entonces, si un animal medicado con un tranquilizante que reduce la activación emocional, ayuda menos o no ayuda a un congénere, se presume que ello puede ser prueba de la importancia de la emocionalidad en la explicación del proceso. Por otro lado, la falta de acceso a los “estados mentales” de los animales menores ha producido una creciente exploración de la influencia del rol del cerebro y el sistema nervioso en su conjunto, sobre el fenómeno de la empatía, y las observaciones reportadas mantienen analogías con los efectos encontrados en humanos. Así, desde una perspectiva neuropsicológica, (Rameson & Lieberman, 2009; Panksepp & Lahvis, 2011), se ha propuesto que la amígdala, intercomunica el sistema de la expresión somática de las emociones (hipotálamo y núcleos del tallo cerebral), que constituye el sistema de los sentimientos como el miedo (cortezas cingulada, parahipocampal y frontal).
Lanteaume et al., 2007 señaló que cuando se estimula eléctricamente la actividad de la amígdala a través de experimentos, en los humanos se produce miedo y aprehensión. Kalin & Shelton (1989), por su parte, afirmaron que los animales, sin la amígdala, generan paralización y miedo, muchos de ellos comparten algunas características de la anatomía y fisiología del sistema límbico humano, presentando también conductas emocionales específicas ante estímulos sensoriales evocados (Filippetti, López & Richaud, 2012).
La “empatía” en roedores por el dolor puede persistir por mucho tiempo; en este fenómeno parecerían estar involucrados el lóbulo prefrontal medial y la circunvolución cingulada anterior, la corteza marginal anterior y la corteza marginal inferior. La persistencia de la reacción emocional ante el dolor del otro, sugiere la existencia un bucle de red neuronal relacionado con la empatía del dolor en el sistema nervioso central (Chen Jun et al., 2015). Según Mejía et al., (2009) y Moya-Albiol, et al (2010), cuando un estímulo externo provoca una emoción en los animales y en el hombre, varios núcleos cerebrales se comunican entre sí, los cuales finalmente activan la contracción y relajación de ciertos músculos faciales.
Se ha descrito también (Langford, 2006) que el dolor en los ratones se comunica a través de diferentes expresiones corporales tales como ojos cerrados, mejillas contraída, frotado de nariz, piloerección y otros. El autor señala que para los animales menores y los seres humanos, el lenguaje corporal es un medio de la expresión emocional que fortalece la comunicación entre individuos. Los ratones comunican estas expresiones, para buscar ayuda de parte de sus congéneres. Por lo tanto, hay suficiente evidencia para pensar que los animales reciben información de los estados emocionales de sus congéneres y reaccionan en consecuencia.
Ciertos roedores pueden transferir su malestar a sus compañeros de jaula durante el proceso de socialización, reduciendo de esta manera, su malestar. No obstante, esto no parecería ocurrir entre animales extraños. Se confirmó que una rata, al cabo de 30 minutos de interacción social con otra compartiendo la misma jaula, el umbral disminuía y la respuesta al dolor mejoraba notablemente (Li et al., 2014). Se realizó un estudio con ratas, que aprendieron a presionar una palanca para obtener comida. Sin embargo, estas dejaban de responder al operando si su acción se combinaba con una descarga eléctrica a otra rata vecina visible (Langford, 2006).
Inhibición de la respuesta emocional.
La suposición de la existencia de un vínculo entre el funcionamiento cerebral y la respuesta empática, ha llevado a postular que es posible inhibir dicho comportamiento actuando sobre las estructuras cerebrales que soportan las emociones. Según la Organización Mundial de la Salud (2004), los psicofármacos, que también se conocen con el nombre de sustancias psicoactivas, pueden modificar el funcionamiento del sistema nervioso central (SNC), y en consecuencia el estado psicológico o los procesos comportamentales de un organismo vivo.
Las benzodiacepinas son inhibidores químicos, compuestas por un anillos de benceno y heterocíclicos capaces de abrir los canales de cloruro activado por el gamma aminobutírico (GABA) que tienen capacidades inhibitorias del SNC. Este neurotransmisor juega un papel determinante en la integración central de las respuestas de estrés, inhibiéndolas en el cerebro. Su uso prolongado disminuye la actividad de partes importantes del cerebro, causando un déficit cognitivo y efectos comportamentales asociados (Armon y Neel, 2013). El clonazepam forma parte de la familia de los ansiolíticos, que también se encuentran incluidos dentro del amplio grupo de las benzodiacepinas. Su principal efecto es la disminución de la actividad eléctrica en el SNC, actuando como sedante, ansiolítico, anti convulsionante y miorrelajante. Este fármaco actúa como estabilizador del estado de ánimo, siendo capaz de provocar en el ser humano, el olvido temporal de ciertos episodios con una gran carga emocional (Rovira, 2018, Rentería-Espejo, 2018).
El clonazepam, una benzodiacepina, se absorbe rápidamente distribuyéndose ampliamente por todos los tejidos del organismo (Aparicio-Medina & Paredes-Vanegas, 2015). En los niños, los efectos farmacológicos del Clonazepam se registran al cabo de los 20-60 minutos, a partir de su administración y permanecen de 6 a 8 horas. En el caso de los adultos, los efectos se instalan por más de 12 horas. La sustancia se metaboliza principalmente en el hígado y es eliminado a través de la orina, al cabo de 22 a 33 horas en los niños, y entre 19 y 50 horas en los adultos (Aronson, 2016).
Según Talegón y Delgado (2009), los animalesmetabolizaneste fármaco más rápidamente que los sereshumanos; su metabolismo se produce mediante el hígado y aseguran que por lo general las benzodiacepinas son seguras y eficaces. Sin embargo, pueden causar efectos secundarios como: sedación, ataxia, relajación muscular, aumento del apetito, excitación paradójica y déficit de memoria. Moutteau-Arias et al., (2019), administraron clonazepan a ratas Wistar, confirmando su efecto inhibidor de la respuesta empática de ayuda y reportaron reducciones del comportamiento motor en dichos animales. Otros autores (Chavez-Marín, 2019 y Rentería-Espejo, 2018), encontraron que el clonazepam aplicado en ratones CD1, ejerce una función protectora contra la ansiedad inducida y afecta el proceso de aprendizaje.
Por lo tanto, si bien las benzodiacepinas son sustancias cuyo empleo puede ser medicinal y por lo tanto necesario, es posible que su ingesta altere también, como subproducto, estados fisiológicos necesarios para el funcionamiento del organismo en el plano individual y social. Esto se aplicaría para el ser humano y también para otras especies.
El propósito del presente estudio es identificar la influencia controlada de la administración del clonazepam sobre la respuesta emocional y sus efectos sobre la conducta de ayudar que presentan los ratones Mus musculus, ante la presencia de un cogénere en necesidad. El supuesto subyacente es que el clonazepam inhibe la actividad emocional a nivel central y con ello reduce las respuestas comportamentales asociadas al rescate de otro ratón en sufrimiento.
MÉTODO
Participantes
Los sujetos experimentales fueron 8 ratones Mus musculus, miembros de una misma camada, todos machos, con pesos entre los 25 a 30 gramos y una edad aproximada de seis meses. Todos los roedores fueron divididos aleatoriamente, según su función, en dos grupos: los roedores cautivos, que deberían ser liberados, y los roedores libres, cuya función consistía en liberar a sus congéneres cautivos. Adicionalmente, todos los sujetos fueron aleatoriamente pareados de manera que cada par esté conformado por un roedor que estará en libertad y otro que sería puesto en cautiverio. En cada pareja se determinó también al azar, cuales de los ratones libres recibirían o no la benzodiacepina (clonazepam). Todos los roedores tuvieron acceso libre a agua (con aditamentos desintoxicantes) y comida balanceada.
Entorno experimental y materiales
El dispositivo de prueba empleado en el presente estudio, fue réplica de una cámara experimental utilizada un estudio similar realizado en la Facultad de Ciencias Médicas de la Universidad de Asunción en Paraguay (Mountteau-Arias et al., 2019). Dicho compartimiento experimental, construido con material acrílico, contaba con dos ambientes separados por una pared del mismo material, cubierta por una tapa. Uno de ellos contenía un área inundable con un pequeño espacio libre de agua donde se situaba el roedor cautivo, y en el otro, un espacio libre de amenaza, se situaba al ratón libre. Ambos compartimientos estaban comunicados por una abertura bloqueada por una puerta que al deslizarse por acción del ratón libre (activando automáticamente un servomecanismo mediante un operando), permitía la salida del ratón cautivo de su encierro (ver Figura 1).
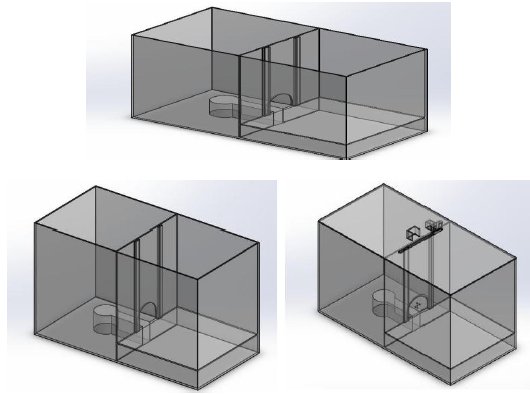
Software. La cámara experimental contaba también con un dispositivo electrónico, similar a otros existentes (ver Drai & Golani, 2001), que permitía registrar la conducta exploratoria del roedor libre, mediante un algoritmo que permitía grabar y mapear térmicamente el movimiento continuo del animal y su ubicación en el espacio donde se encontraba alojado, especialmente alrededor del mecanismo de apertura de la cámara experimental (ver Figura 2).

Figura 2 División del espacio experimental en cuadrantes para facilitar la ubicación del animal en el espacio libre de restricción (arriba), y vista de los animales con la ayuda del dispositivo electrónico para medir su posición (medio y abajo).
Variables
Variable independiente. El procedimiento tuvo diez días de duración, siguiendo las recomendaciones de Aparicio-Medina & Paredes-Vanegas (2015). Durante este tiempo, se administró el medicamento en dosis sesiónales que combinaban 3,125 ml de Clonazepam y 96,675 ml de suero fisiológico. La sustancia fue administrada a los roedores mediante vía oral, a través de una cánula bucal, vigilando que todos los sujetos experimentales en condición de libertad, reciban con la precisión necesaria, la misma dosis. Los sujetos libres de la condición control, no recibieron la variable independiente. La administración del fármaco en el grupo de roedores libres (experimentales), fue realizada por una veterinaria y una bióloga, especialistas en el manejo de roedores.
Variables dependientes. En la presente investigación se cuantificaron dos variables dependientes: a) la latencia de liberación del ratón cautivo por el ratón en libertad, definida por el tiempo en segundos entre el inicio de la prueba y la activación del mecanismo que abría la puerta); y b) la activación emocional del roedor libre, definida como la densidad de visitas de dicho roedor al cuadrante 5.
Procedimiento
Se llevaron a cabo veinte sesiones experimentales, con un tiempo de duración de 30 minutos cada una. Diez sesiones transcurrieron sin la administración del fármaco (Línea Base), mientras que en las diez restantes (condición Experimental) sí se administró el Clonazepam.
Condición de Línea de Base (LB). En la condición de LB, se colocó al roedor libre en el área seca y al ratón cautivo en el área inundada. Seguidamente, se iniciaba el registro de la latencia de la apertura del dispositivo de liberación, mediante el software respectivo. Los roedores libres debían deslizar la palanca de la puerta para liberar a su congénere, cesar con ello su condición de cautiverio y detener el contador que marcaba la latencia de la operación. A continuación, una vez liberado el ratón en cautiverio, se permitía la interacción de ambos animales por espacio de un minuto, antes de dar inicio a una segunda ronda de liberación, repitiéndose invariablemente el mismo procedimiento.
Condición experimental. Durante esta condición, se siguió el mismo procedimiento realizado en LB, también durante 10 días. Sin embargo, a diferencia de la condición de LB, el protocolo incluyó la administración del fármaco en la dosis anteriormente mencionada. Antes de la iniciación de la sesión, el ratón libre recibía el Clonazepam por vía oral. Se administró el medicamento 40 minutos antes de iniciar cada sesión, considerando que en este lapso, la sustancia habría ejercido el efecto esperado en un organismo de las características de un Mus musculus (Aronson, 2016).
Análisis estadístico
La presente investigación experimental se ejecutó mediante un diseño de ANOVA de medidas repetidas intrasujeto (latencias de liberación y visita/presencia en el cuadrante 5) en 10 sesiones sucesivas y dos factores intersujeto (con y sin ingesta de la sustancia).
RESULTADOS
Los datos recabados de la presente investigación fueron analizados tanto mediante la comparación de medias como con la prueba de análisis de varianza lineal mixto (ANOVA MIXED), con la ayuda de los programas IBM-SPSS v27.0 y Excel v2020.
Efectos del Clonazepam en la latencia de liberación
En la Tabla 1, se muestran las latencias de apertura de la puerta expresadas en segundos para cada una de las sesiones diarias, así como sus valores promedio para ambas condiciones. Se utilizaron 4 roedores para el grupo en condición LB y otros 4 roedores para el grupo de CE.
Tabla 1 Latencias (en segundos) de la conducta de liberación de los RL, en las condiciones de LB y tratamiento experimental.
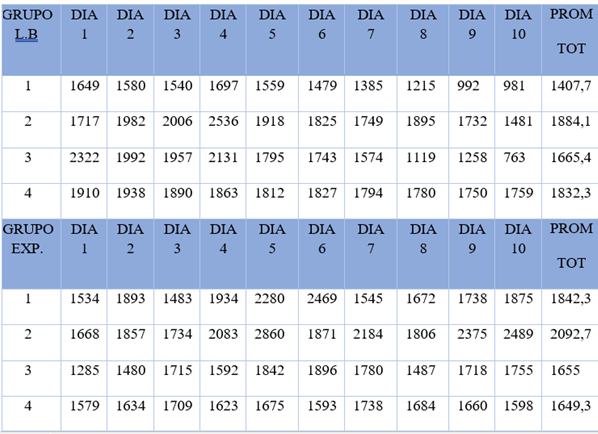
Nótese que el promedio obtenido de los 4 sujetos durante Línea Base (M LB = 1697,2 segs.) es menor al empleado por los ratones experimentales (M exp=1809,7 segs.). No obstante, la diferencia entre ambos grupos no es estadísticamente significativa (t = -,752, p >,05), lo que querría decir que la dosis de clonazepam utilizada no pareció haber influido en el aumento de la latencia en la apertura del dispositivo que liberaba a los ratones cautivos. La siguiente figura, muestra las diferencias entre los promedios de latencia de los ratones por sesión, con y sin la sustancia, aunque tales variaciones son estadísticamente irrelevantes
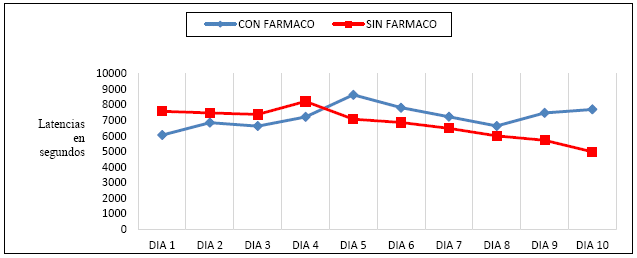
Figura 3 Tendencias de la latencia de liberación o de apertura de la puerta, observadas en los ratones controles y experimentales, a lo largo del experimento.
Activación emocional en los ratones libres
La literatura reciente (Ben-Ami Bartal et al., 2011), señala que la respuesta prosocial manifestada por diferentes especies es generalmente antecedida por una respuesta emocional suscitada por la percepción de una situación de necesidad imperiosa manifestada por un congénere, conocida como “preocupación empática”. En el presente estudio, la respuesta de activación emocional fue definida como porcentaje de visitas del animal al cuadrante 5 (contiguo a la puerta de acceso a la zona de encierro del animal cautivo) (Roth y Flores, 2019). La Tabla 2 muestra dicha proporción de visitas del RL a dicho cuadrante en las condiciones de LB y experimental respectivamente, registradas con el dispositivo electrónico.
Tabla 2 Activación emocional (porcentaje de visitas al cuadrante 5) presentada por los RL, en las condiciones de LB y Experimental.
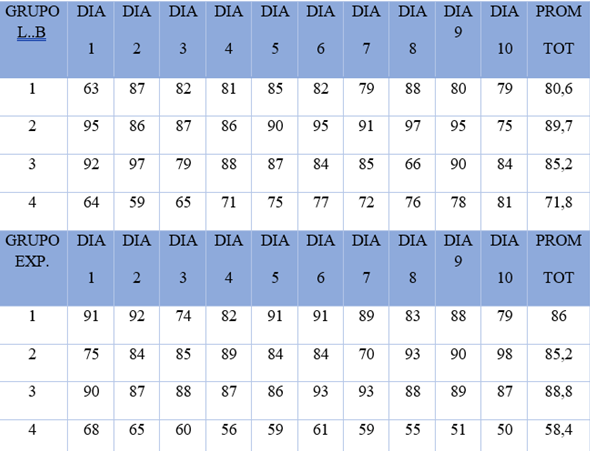
La Tabla 2 y la figura 4, resumen los porcentajes de tiempo que cada uno de los RL pasaba en el cuadrante 5, en ambas condiciones. Adviértase que durante la condición de
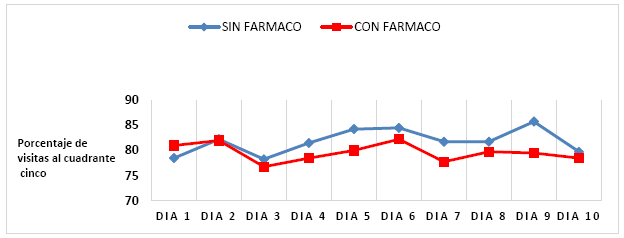
Figura 4 Tendencia porcentual de visitas al cuadrante 5 de los animales del estudio, observada en los ratones controles y experimentales, a lo largo del experimento
Línea base (sin fármaco), el porcentaje de visitas al cuadrante 5 fue de 32,7%, ligeramente más alto que en la fase experimental.

A continuación, se ensayó el ANOVA para medidas repetidas sobre ambas variables. La información obtenida presenta en primera instancia, las pruebas de efectos intrasujeto tanto para Activación Emocional como para Latencia de Liberación. Debido a la confirmación de la imposibilidad de obtener el valor de la Esfericidad de Mauchly, se verificó el indicador Huynh-Feldt para determinar la significación de las medidas repetidas. Adviértase que las medidas de Activación Emocional arrojaron una F = 424,194, p < 0,001 y una η 2 = 0,993, indicando diferencias significativas entre las medidas obtenidas a lo largo de las 10 sesiones, mostrando un paulatino decremento de la activación emocional por efecto del clonazepam. En el caso de la variable Latencia de Liberación, en cambio, las medidas intrasujeto no mostraron diferencias significativas (F = 0,647, p > 0,05, η 2 = 0,177) atribuibles a la sustancia, indicando que la latencia en los diferentes ensayos sucesivos permanecieron invariantes a pesar del clonazepam. Asimismo, tampoco se observaron interacciones entre las variables Activación Emocional y Latencia de Liberación (F = 0,766, p > 0,05, η 2 = 0,203). Las comparaciones por parejas para ambas variables se presentan en la Tabla 4, y confirman las diferencias significativas entre las condiciones estudiadas para la variable Activación Emocional pero no para la variable Latencia de Liberación.
Tabla 4 Comparaciones por parejas con Bonferroni que describen las relaciones entre las variables Activación emocional y latencia de liberación.
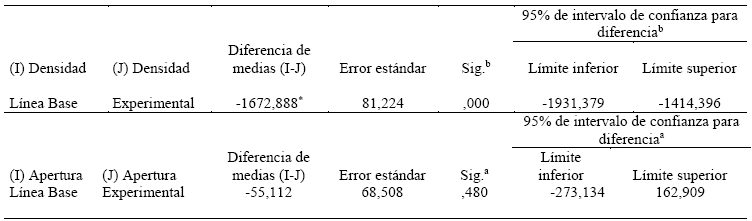
Se basa en medias marginales estimadas
*. La diferencia de medias es significativa en el nivel .05.
b. Ajuste para varias comparaciones: Bonferroni.
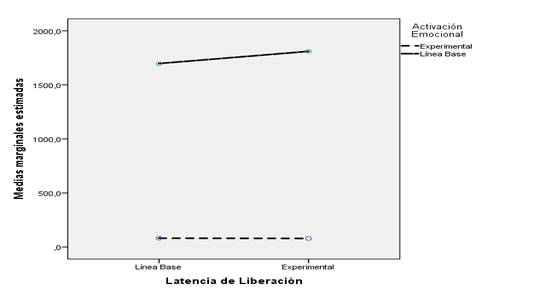
Las relaciones descritas estadísticamente son representadas en la Figura 5. Nótese que para la variable Activación Emocional, las diferencias entre las condiciones de línea base y experimental, son notables. Por el contrario, las diferencias entre las mismas condiciones de la variable Latencia de Liberación, son irrelevantes.
DISCUSIÓN
La evidencia empírica (Church, 1959; Rice & Gainer, 1962; Ben-Ami Bartal, et al, 2011, 2014, 2016; de Waal, 2012; Mojil, 2012; Horner et al, 2011; Mason 2014) permite afirmar que los roedores y una amplia variedad de especies exhiben conducta de ayuda hacia sus congéneres y otros animales diferentes. No obstante, esta descripción se realiza por analogía del comportamiento humano y a partir de inferencias de procesos mentales y emocionales derivados de la auto-observación. Ciertamente, cualquier observador es capaz de advertir que ciertos animales menores, al igual que la especie humana, muestran frecuentemente conductas de ayuda, no obstante la descripción y explicación parsimoniosas, especialmente en el caso de la conducta animal, aconseja cautela en el uso de la analogía.
El presente estudio confirmó la evidencia existente acerca de la capacidad de ciertos roedores de mostrarse reactivos a las urgencias de un compañero percibido en circunstancias de necesidad inducida por confinamiento y amenaza. En el presente caso, dichas “urgencias” fueron transmitidas al animal observador quien pareció manifestar un comportamiento inusual, definido por los autores como activación emocional y medidas a través de la densidad de sus respuestas en las proximidades de la puerta que impedía abandonar la situación aversiva. En tales circunstancias, el animal libre aprendió rápidamente a destrabar el dispositivo que impedía la salida del ratón cautivo y acabar con ello, su respuesta emocional y presumiblemente también la del compañero.
La literatura muestra también que la conducta de ayuda o prosocial es una consecuencia clara de la experiencia emocional suscitada por la constatación del sufrimiento del otro, al extremo de que ciertos autores (Batson, 2009) la considera una condición necesaria para explicar la conducta altruista. El propósito del presente estudio fue verificar si en el caso de los roedores Mus musculus, esta relación se cumplía en los mismos términos que en nuestra especie. Por lo tanto, se manipuló la activación emocional del ratón en libertad, neutralizándola con una dosis de clonazepan y con ello se pretendía reducir la conducta de liberación (Moutteau-Arias et al., 2019).
Estos autores encontraron que la benzodiacepina tenía efectos claros sobre el comportamiento de ayuda en ratas, si se bloqueaba el “procesamiento afectivo” del animal. Los resultados obtenidos en la presente investigación no resultaron ser tan concluyentes. En efecto, se observó una reducción significativa de la conducta emocional causada por la medicación, aunque ello no supuso un decremento en la conducta de ayuda o liberación. Aún bajo los efectos del clonazepan, los ratones no mostraron variaciones en la latencia de apertura de la puerta.
Este resultado puede prestarse a varias consideraciones. En primer lugar, la modalidad de administración del medicamento vía cánula oral podría afectar a la eficacia del medicamento debido al tiempo de absorción del mismo a través del tracto digestivo y explicar parcialmente el resultado. En segundo lugar, si bien la dosis de clonazepan administrada mostró reducir la activación del animal, es posible que haya sido insuficiente para bloquear la conducta de liberación. En tercer lugar, no debe descartarse la posibilidad de que en este tipo de roedores, la conexión entre la respuesta emocional y la conducta de ayuda no sea todo lo robusta que se espera en analogía a la conducta humana. De aquí se desprende la posibilidad de la existencia de otros mecanismos de activación de la ayuda, basados por ejemplo, en la interacción social de los individuos facilitada por la convivencia (cohabitación) y la familiaridad (Roth & Flores, 2019), que fueron restringidos en la prueba experimental utilizada.
Ciertamente, la liberación del congénere puede ser negativamente reforzada al reducir la respuesta emocional concomitante y positivamente, al darse paso a la interacción social que sigue a la liberación. No debe descartarse que la conducta de ayuda esté biológicamente sustentada en la fuerza de la propia supervivencia, en cuyo caso debería prestarse atención al aporte de los múltiples procesos neurofisiológicos y químicos de los organismos cuya función es la perpetuación de la especie (de Waal, 2012; Mason, 2014; Jeon y Cols., 2010; Langford y Cols., 2010). El presente estudio fue realizado exclusivamente con ratones machos. Esta circunstancia limita sin lugar a dudas, los alcances de la investigación y deja interrogantes sobre la influencia sexual en el comportamiento de ayuda en la especie estudiada. Finalmente, la investigación futura deberá ofrecer mayores elementos de juicio acerca del comportamiento animal prescindiendo en lo posible de modelos generados en la investigación humana.

