










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAjayu Órgano de Difusión Científica del Departamento de Psicología UCBSP
versión On-line ISSN 2077-2161
Ajayu v.7 n.2 La Paz ago. 2009
ARTÍCULO
INFLUENCIA DEL INCREMENTO EN LA MAGNITUD DE RAZÓN FIJA SOBRE EL CONTRAFREELOADING EN ORGANISMOS INFERIORES
EFFECTS OF INCREASED FIXED RATIO MAGNITUDES ON CONTRAFREELOADING IN LABORATORY MICE
Yolanda Ferreira Arza* y J. Daniel Walker M1.
Universidad Católica Boliviana “San Pablo”, Departamento de Psicología, Bolivia
* Directora de la investigación
1 La investigación fue realizada para la obtención de Licenciatura en Psicología. Quiero agradecer a la Lic. Yolanda Ferreira, al Dr. Erick Roth, a la Lic. Carmen Wilde y al Lic. Rubén Belmonte por el apoyo en la elaboración de este trabajo.
Dirigir correspondencia al autor: Departamento de Psicología, Universidad Católica Boliviana “San Pablo”. E-mail: jdaniel.walker@yahoo.com
Resumen.
En un ambiente en constante cambio, los animales deben adaptar su comportamiento para reducir la incertidumbre, incluyendo la conducta del forrajeo. El Modelo de Primacía de Información considera al forrajeo y la exploración como medios para obtener comida e información del ambiente. Este modelo explica un comportamiento denominado Contrafreeloading (CFL). CFL ocurre cuando los animales emiten una respuesta operante para obtener un refuerzo positivo, mientras que la misma recompensa existe libremente e ilimitado. Estudios pasados reportan que niveles de CFL se relacionan inversamente al esfuerzo requerido en la tarea operante. Para estudiar el CFL, se realizó un experimento con ratones de laboratorio (mus musculus) como sujetos experimentales, utilizando cinco magnitudes de razón fija para representar diferentes grados de esfuerzo. Los resultados apoyan estudios previos que CFL disminuye mientras el esfuerzo aumenta. Tres de los cuatros sujetos experimentales emitieron significativamente menos respuestas en RF2 que en RF1 (reforzamiento continuo), un descenso gradual desde RF2 a RF4, y una ausencia de respuestas en RF5. Simultáneamente, alimentarse de la comida libre se relacionó positivamente con el incremento de esfuerzo. Resultados adicionales indican una tendencia correlativa positiva entre esfuerzo y el comienzo de CFL, sugiriendo la necesidad de futuros estudios para determinar tendencias similares.
Palabras Claves: forrajeo, contrafreeloading, ratones, esfuerzo
Abstract.
In an ever changing environment, animals must adapt their behavior so as to reduce uncertainty, including foraging tendencies. The Information Primacy Model considers animal foraging and exploration as a way to gather not only food but also environmental information. This model explains a behavior called Contrafreeloading (CFL). CFL occurs when animals emit an operant response to obtain a positive reinforcer, while the same reward exists freely and unrestricted. CFL levels have been reported to be inversely related to effort required in an operant task. To study CFL, an experiment was conducted with laboratory mice (mus musculus) as test subjects, where five fixed ratio magnitudes represented different degrees of effort. Results support previous studies that CFL diminishes whilst effort increases. Three of the four test subjects presented a significant decline in responding in FR2 from FR1 (continual reinforcement), a gradual decline from FR2 through FR4, and an eventual absence of responding in FR5. Simultaneously, feeding from the free food source was positively related to effort increase. Additional results indicate a positive correlative tendency between effort and the commencement of the CFL behavior, suggesting the necessity for future research to determine similar tendencies.
Key Words: foraging, contrafreeloading, mice, effort
INTRODUCCIÓN
La conducta de forrajeo se observa cuando la homeostasis interna de un organismo se encuentra en falta. En dicha situación, el organismo pone en marcha una serie de conductas que previamente fueron reforzadas al encontrar comida (Skinner, 1969). Lo que manifiestan es un comportamiento óptimo, de acuerdo el tiempo y esfuerzo que implementarán para obtener la mayor ganancia (Bautista, 1997).
Muchos factores (i.e., densidad de alimento, tipo de alimento, esfuerzo) hacen que el ambiente fluctúe de estación a estación (Vaz-Ferreira, 1984), por lo que la incertidumbre es inevitable (Inglis, Langton, Forkman & Lazarus, 2001). Los animales desarrollan esquemas de alimentación para lograr mantener estable la cantidad de ingesta aun con cambios internos (físicos, del mismo animal) o externos (ambientales, como ser diluvios o sequías). El comportamiento será óptimo cuando reconocen estos cambios y determinan qué esquema utilizar (Day, Kyriazakis & Rogers, 1998).
Es así que la conducta de forrajeo no solo le permite al organismo alcanzar las exigencias nutricionales para subsistir (Vera, 2002). Más bien, también brinda al individuo información sobre todas las fuentes de alimentación, incluyendo las fuentes alternas disponibles (Inglis & Langton, 2006).
Como explica el Information Primacy Model, una conducta de forrajeo constante permite al organismo reducir la incertidumbre ambiental (Gilbert-Norton & Gee, 2005; Talling, Inglis, van Driel, Young & Giles, 2002). Mientras el animal adquiere informática ambiental continuamente, renueva su conocimiento de la situación ambiental. Esto permite al organismo responder adecuadamente cuando las condiciones cambian (Inglis, Langton, Forkman & Lazarus, 2001).
Este modelo ha sido utilizado para explicar la conducta denominada contrafreeloading. Considerando que el freeloading hace referencia al hecho de alimentarse de comida disponible libremente, el contrafreeloading (CFL) es el hecho de responder o trabajar para obtener alimento cuando, en el mismo ambiente, existe una fuente ilimitada y sin restricciones del mismo alimento (Bachá, 2002; Bean, Mason, & Bateson, 1999; Coburn & Tarte, 1976; Gilbert-Norton & Gee, 2005; Inglis, Forkman, & Lazarus, 1997; Jensen, 1963; Lindqvist & Jensen, 2003; Lindqvist, Zimmerman, & Jensen, 2006; Robertson & Anderson, 1975; Rutter & Nevin, 1990).
Los etólogos expresan que los comportamientos suelen ser específicos a una especie (Vaz-Ferreira, 1984). Sin embargo, se ha demostrado que el CFL no es una conducta particular de una sola especie, observando dicha conducta en una variedad de especies, como ser ratas (Bachá, 2002; Coburn & Tarte, 1976; Jensen, 1963; Neuringer, 1969; Osborne & Shelby, 1975; Rutter & Nevin, 1990), palomas (Clement, Feltus, Kaiser, & Zentall, 2000; Neuringer, 1969), otras especies de aves (Jensen, 2000; Lindqvist & Jensen, 2003; Lindqvist, Zimmerman, & Jensen, 2006; Schütz & Jensen, 2001), chimpancés (Menzel, 1991), hasta en organismos humanos (Singh, 1970; Tarte, 1981), demostrando su amplitud como conducta inter-especie.
Se enfatiza que tanto la comida obtenida por trabajar como la comida libre son idénticas (Gilbert-Norton & Gee, 2005). El alimento obtenido como reforzador posterior a una conducta operante conlleva esfuerzo mientras que la comida libre no posee ninguno (Lindqvist, Zimmerman, & Jensen, 2006). Sin embargo, para el mantenimiento del CFL a partir de una conducta operante es necesario la presencia de una consecuencia (reforzador positivo), sin el cual el CFL cesará (Bachá, 2002).
Dicha conducta parece contradecir la teoría clásica de forrajeo óptimo (Inglis, Forkman & Lazarus, 1997; Inglis, Langton, Forkman & Lazarus, 2001; Forkman, 1996; McGowan, Newberry, Robbins & Alldredge, 2005) donde un animal busca maximizar la relación entre beneficio o ingesta y el esfuerzo o costo de obtener su alimento. Este principio afirma que un animal debería obtener la máxima cantidad de alimento, gastando la mínima cantidad de energía (Menzel, 1991), ya que evolutivamente sería lógico ahorrar energía cuando existan fuentes de comida libre (Jensen, 2000).
La información que los animales obtienen al emitir conductas de forrajeo quizá no sea necesaria en el momento particular, pero llega a ser información útil a corto plazo, permitiendo al individuo desplazarse a esas fuentes alternas de alimento si es que su fuente principal fuera a desaparecer o reduce su rentabilidad (Inglis, Langton, Forkman & Lazarus, 2001; Menzel, 1991). Por lo que conductas cómo el CFL permiten al animal mantenerse actualizado sobre la información de recursos impredecibles, dándole control sobre ambientes variables (Gilbert-Norton & Gee, 2005).
De acuerdo al Information Primacy Model, realizar el CFL permite la obtención tanto de alimento como de información actual del ambiente a partir del forrajeo y la exploración. El animal se informa sobre la rentabilidad de fuentes inciertas, sub-óptimas o desconocidas de alimento que quizá no frecuente, pero que le servirán si en el futuro su fuente primaria desaparece (Gilbert-Norton & Gee, 2005) y la secundaria se convierta en la principal (Bean, Mason & Bateson, 1999; Inglis, Forkman & Lazarus, 1997).
Sin embargo, el Neophobia-Optimality Model argumenta que el CFL se da por la aversión que el animal siente hacia el contenedor con comida libre por su novedad. Explica que animales expuestos previamente a este contenedor prefieren la comida libre y los que no son expuestos prefieren trabajar por su alimento. Por lo tanto, es el miedo al comedero libre que hace que los animales presenten el CFL (Mitchell, Becnel & Blue, 1981; Mitchell, Fish & Calica, 1981; Mitchell, Scout & Williams, 1973). Sin embargo, el estudio de Carder y Beckman (1975) demostró que ratas expuestas previamente al contenedor y a la comida libre antes de la fase de prueba respondieron de la misma manera que ratas que no fueron expuestas previamente, discrepando con este modelo.
Específicamente, Inglis et al. (1997) postularon que existen factores adicionales que intervienen en la frecuencia del CFL. Entre los que mencionan la naturaleza del forrajeo o tarea, la manipulación del ambiente, la incertidumbre ambiental, el cambio de estímulos, extensión de entrenamiento previo, condiciones de crianza, nivel de privación y el esfuerzo requerido (para más detalle ver: Inglis, Forkman & Lazarus, 1997).
Asimismo, otros autores detallan similitudes y adiciones, donde el CFL es influido por la incertidumbre ambiental (Inglis, Langton, Forkman & Lazarus, 2001; Lindqvist & Jensen, 2003), el tipo de reforzador (Powell, 1974), si existen reforzadores secundarios (Osborne & Shelby, 1975), el valor del reforzador (cualidad o cantidad), la especie del animal (Powell, 1974), si existen otras maneras para obtener información aparte de explorar (inspeccionar visualmente una fuente alimenticia) (Bean, Mason & Bateson, 1999), y la extensión de entrenamiento de la respuesta operante (Powell, 1974; Tarte & Rasmussen, 1979). Sin embargo, algunos estudios demostraron que el CFL se puede presentar sin previo moldeamiento (e.g., Bachá, 2002; Rutter & Nevin, 1990). Las condiciones de albergue y crianza también influyen sobre el CFL, sorprendentemente habiendo demostrado que los animales en ambientes empobrecidos exhiben mayores cantidades de conductas exploratorias, y por ende mayor CFL (Coburn & Tarte, 1976).
Tanto el hambre como el esfuerzo para obtener alimento a través del trabajo son considerados por Inglis et al. (1997) esenciales al caracterizar a esta conducta y postulan tres niveles de la misma. Los autores detallan que existen niveles bajos (niveles elevados de hambre y esfuerzo), intermedios (nivel de hambre es bajo y el de esfuerzo es elevado, o viceversa) y elevados (niveles de hambre y esfuerzo son bajos) de CFL.
De esta manera, el nivel de probabilidad de ocurrencia del CFL aumenta progresivamente mientras más fácil sea la obtención de la fuente de alimentación (también: Carder & Berkowitz, 1970) y menos hambre tenga el animal (también: Bean, Mason & Bateson, 1999; Neuringer, 1969; Tarte & Rasmussen, 1979). Lo que sucede al incrementar el esfuerzo para obtener un reforzador, en lugar de alimentarse de comida libre, es que “equivale a la reducción de energía neto que puede ser obtenido de la fuente de alimento ganado” (Inglis, Langton, Forkman & Lazarus, 2001, p. 549). Sin embargo, algunos estudios reportan que los animales se esfuerzan mucho por obtener alimento como reforzador (Taylor, 1975).
Por tanto, se vio necesario demostrar empíricamente que el nivel de esfuerzo disminuye el nivel de CFL, y si los organismos emiten el CFL con niveles mayores de esfuerzo. Mientras que autores (e.g., Inglis, Forkman & Lazarus, 1997; Inglis, Langton, Forkman & Lazarus, 2001) describen una relación directa entre esfuerzo y CFL, no plantean la tendencia de dicha relación. Es a partir de ello que la presente investigación se elaboró para determinar en qué medida el incremento de la magnitud requerida en un programa de razón fija (RF) influye sobre la conducta de CFL en organismos inferiores.
MÉTODO
Sujetos. Se utilizaron cuatro ratones experimentalmente ingenuos, de la Familia Muridae, Género Mus y Especie músculus (mus músculus) como sujetos experimentales. Las edades de los sujetos fueron al inicio de la investigación oscilaron alrededor de los seis meses. Aunque no se pudo controlar la edad exacta, se aseguró que sus tamaños fueran lo más adecuado para los requerimientos del programa experimental, así como para los aparatos a usarse. Al inicio de la experimentación, el peso ad libitum de los sujetos fluctuó alrededor de los 32 g. Se albergaron a los sujetos en jaulas hogares individuales, con acceso ilimitado de agua, con algodón y aserrín para higiene, y una rueda, en un ambiente bajo un ciclo de 12 horas de luz y 12 horas de oscuridad.
Durante la experimentación los sujetos fueron mantenidos en privación alimenticia a un 80% de su peso ad libitum. De acuerdo a Wilde y Otondo (1990), los pellets fueron hechos en base a harina blanca, harina amarilla, agua, en proporciones iguales, vitaminas y aceite de bacalao.
Ambiente. Se realizó la presente investigación en el Laboratorio de Conducta Animal del Departamento de Psicología ubicado en el Bloque R de la Universidad Católica Boliviana “San Pablo”, en la ciudad de La Paz, Bolivia. En dicho ambiente se almacenaban los espacios experimentales y se albergaban a los sujetos experimentales en sus jaulas hogares debidamente. El ambiente constó de aislamiento sonoro, ventilación y una estufa, asegurando tanto el control de las variables ambientales como la salubridad de los sujetos experimentales.
Variables. La VI de la presente investigación fue la comida libre que se introdujo dentro del espacio experimental en condiciones determinadas. La VD fue el número de respuestas de la conducta de CFL, entendiendo dicho concepto como una conducta de presionar un operando para la obtención de alimento (reforzador positivo) cuando, en el mismo ambiente, existe acceso libre al mismo tipo alimento (VI). La VD fue dividida en categorías y estudiada bajo diferentes niveles de esfuerzo, representado por diferentes magnitudes de RF (entre RF1 a RF5), constituyendo cada vez mayor nivel de esfuerzo.
Aparatos. Se constó de una balanza para pesar a cada sujeto experimental, y para medir el alimento diario. Dentro del área de albergue se mantuvo la temperatura estable entre 24-26º C. El espacio experimental fue de Lafayette Instrument Co., modelo 84222-SS. Dicho espacio tenía una superficie de 20 cm. por 27,5 cm., con una altura de 21 cm. Dentro de dicho espacio, se encontraban tanto el operando, de 5 cm. de largo a una altura de la base de 7 cm., ubicado al centro de la pared principal; un estímulo discriminativo visuales (luz blanca) a 3 cm. por encima del operando; y el comedero de 5 cm. Además, cuando la condición experimental requirió de un comedero libre se utilizó una tapa de plástico con dimensiones casi similares al comedero del espacio mismo. Así, este comedero libre tenía un diámetro de 4,5 cm. y una altura de 2,5 cm. Este Espacio Experimental fue manejado manualmente, donde el acto de encender y apagar el estímulo discriminativo (luz blanca), así como la entrega del pellet (reforzador positivo), fueron manejados por el experimentador directamente.
Procedimiento. Previo el inicio del estudio, se observó el tiempo que los sujetos tardaban en consumir la porción de alimento asignado diariamente. De acuerdo a estas observaciones, se calcularon sesiones diarias de 30 minutos de duración, introduciendo a un sujeto dentro del espacio experimental a la vez.
Etapa 1 (Adaptación). Inicialmente, un sujeto fue colocado en el espacio experimental a la vez durante tres sesiones experimentales sin ninguna estimulación. Estas sirvieron como adaptación, para eliminar conductas emocionales provocadas por estar expuestos a un ambiente nuevo (Wilde & Otondo, 1990) que podrían interferir en el moldeamiento posterior. Los sujetos tuvieron acceso ilimitado al alimento y agua.
Etapa 2 (Privación). Posterior a la tercera sesión, se introdujo a los sujetos en sus jaulas hogares sólo con acceso a un bebedero con agua. Los sujetos fueron expuestos al único día de privación total de 24 horas. A partir del peso ad libitum se calculó el peso experimental a un 80% del peso inicial para mantener a los sujetos privados pero sin arriesgar su salubridad (Martin & Pear, 1999).
Etapa 3 (Moldeamiento al Comedero). Se trabajó una sesión de moldeamiento al comedero, estableciendo una asociación entre el comedero y el sonido de la caída del pellet (reforzador positivo). Por cada acercamiento al comedero, el sujeto recibía un reforzador.
Etapa 4 (Condiciones Experimentales). Antes de comenzar con el trabajo experimental en sí, se asignó de manera aleatoria a uno de los animales para ser el sujeto control. Los tres animales que restaron fueron experimentales, propiamente dicho.
Etapa 5 (Moldeamiento al Operando). Durante la etapa de moldeamiento al operando, se entrenaron a los sujetos a presionar el operando para así recibir alimento, utilizando las técnicas de aproximaciones sucesivas y reforzamiento diferencial. Cuando los sujetos respondieron de acuerdo a la clase de respuesta de presionar el operando con sus dos patas delanteras (RF1) de manera constante, se iniciaron las sesiones de la primera etapa experimental propiamente dicho.
Etapa 6 (RF1). La sexta etapa fue dividida en cuatro fases, de las cuales dos correspondieron a la Línea Base (A) y dos al Tratamiento (B). Fueron trabajadas de manera intercalada (A-B-A-B), tres sesiones por fase. Durante las fases de Línea Base, contingentemente a las respuestas de RF1 los sujetos recibieron un reforzador. Por su parte, para las sesiones de Tratamiento se introdujo la comida libre (VI) en su propio recipiente al espacio experimental. Los sujetos podían consumir la comida libre o emitir RF1 para recibir un reforzador. Se consideraba una conducta de CFL en cuanto los sujetos emitían la conducta operante (RF1) en presencia de la comida libre. En ambas fases se daba por finalizada una sesión cuando se concluyeron los 30 minutos o al acabar la alimentación diaria. Los tres sujetos experimentales fueron expuestos a este procedimiento, mientras que el sujeto control solo trabajó sesiones de Línea Base.
Etapa 7 (RF5). La siguiente etapa fue subdividida en cuatro etapas, y el procedimiento fue llevado a cabo de igual manera a la etapa anterior y tuvo la misma duración. Sin embargo, los reforzadores fueron entregados contingentemente a la clase de respuesta de RF5, donde los sujetos debían presionar el operando con sus dos patas delanteras cinco veces sucesivas sin bajar sus patas del operando.
Etapa 8 (Reorientación Experimental). Dado el hecho de que los tres sujetos experimentales no emitieron respuestas de RF5 en la etapa anterior, menos un sujeto que en una sola sesión experimental emitió seis respuestas, se procedió a una reorientación experimental. En ella, se determinó estudiar todas las magnitudes intermedias entre RF1 y RF5, incluyendo éstas. Además, el sujeto control fue asignado como el cuarto sujeto experimental para así buscar mayor grado de generalización.
Etapa 9 (RF1). El procedimiento de la novena etapa fue manejado de igual manera a la sexta previamente descrita. Se trabajaron las cuatro fases correspondientes a la Línea Base (A) y al Tratamiento (B) de manera intercalada (A-B-A-B), pero en lugar de tres sesiones por fase, solo se trabajaron dos. La clase de respuesta, o criterio conductual, que fue reforzada fue de RF1. Por tanto, contingente a una respuesta sobre el operando, los sujetos recibían un reforzador. Este criterio fue mantenida tanto en condiciones sin la comida libre (A) como durante el Tratamiento (B), siendo las respuestas en las sesiones de Tratamiento consideradas CFL.
Etapa 10-13 (RF2, 3, 4, 5). Siguiendo el mismo procedimiento a la etapa anterior, los únicos cambios que se presentaron en las últimas cuatro etapas (con sus cuatro fases correspondientes a cada etapa) fueron las clases de respuesta. En la magnitud de RF2, los sujetos fueron exigidos a presionar el operando dos veces consecutivas sin bajar las patas delanteras del operando. De igual manera, RF3 fue observada con tres presiones consecutivas, RF4 con cuatro y, finalmente, RF5 se cumplía con cinco presiones consecutivas sobre el operando con las dos patas delanteras sin bajarlas del operando. La etapa decimotercera fue la que dio fin a la presente investigación debida a que el número de respuestas fue nulo.
RESULTADOS
En las Etapas 6 y 7 se observó el nivel de CFL en las magnitudes de RF1 y RF5, respectivamente. Las condiciones de Tratamiento (B) correspondieron a la conducta CFL propiamente dicho. Mientras que los sujetos experimentales presentaron la conducta de CFL en RF1, el número de respuestas en RF5 descendió a cero. Debido a la imposibilidad de analizar una tendencia solo entre las dos magnitudes de RF, se realizó una reorientación experimental para lograrlo.
Los datos que siguen corresponden a las Etapas 9-13. En ellas, se observó el CFL en todas las magnitudes entre RF1 y RF5. En la Figura 1 se observa el promedio de número de respuestas operantes en condiciones de Tratamiento (B) por sujeto. Lo más representativo al observar esta figura es la tendencia de disminución de los promedios de respuestas de los sujetos experimentales Nº 1, 2 y 4 mientras la magnitud de RF aumenta, hasta llegar a cero respuestas en RF5. Adicionalmente, se observó que gradualmente los sujetos experimentales consumieron mayor cantidad de comida libre en lugar de trabajar por ello. Lo dicho demuestra que mientras aumenta la magnitud de RF (esfuerzo), el número de respuestas de CFL disminuye y el consumo alimenticio como comida libre aumenta.
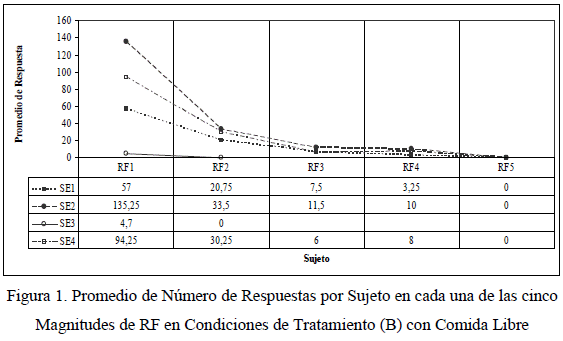
Aunque en la figura anterior se observa una disminución de respuesta relativamente innegable, se acudió a un análisis estadístico para determinar si dicha disminución fue significativa. Se realizó la prueba de análisis de varianza noparamétrico de Kruskal-Wallis, el cual es un método libre de distribución y utiliza las estadísticas de orden de rangos (Kerlinger & Lee, 2002; Lind, Marchal & Mason, 2004; Wackerly, Mendenhall & Shaeffer, 2003):
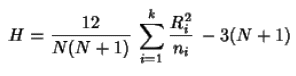
donde µi representa el promedio de la respuesta de la magnitud i. Dicha prueba estadística permitió observar cómo Y (número de respuestas) se comporta en función de la magnitud de RF. Los resultados de la prueba dieron resultados de y = 103,01x-0.527, y R2 = 0,8321. El dato más importante es la significancia, que en este caso es asintótica debido a que es una prueba aproximada por medio de la Chi-cuadrada. Se demostró que a un nivel de significancia de 0.05, existen diferencias significativas entre los distintos tratamientos. Es decir, el comportamiento varía de una magnitud de razón fija a la siguiente, de manera significativa.

Complementariamente a los datos relacionados con el número de respuestas, la Figura 2 es una determinación relacionada al Tiempo de Reacción al Estímulo Discriminativo Inicial (TRALEDi). Este dato se refiere al tiempo (en minutos) que un sujeto tarda en emitir su primera respuesta de la sesión experimental. En cada sesión se obtuvo un dato respecto al TRALEDi, y posteriormente se promediaron los datos detonas las sesiones correspondientes a la condición de Tratamiento (B) en cada magnitud de RF por sujeto. La ausencia de datos relacionados con el tercer sujeto experimental es debido a la falta de respuestas en RF2, y la finalización de su trabajo experimental luego de esta magnitud por lo que no fue pertinente demostrar que su promedio de TRALEDi para RF2 fue de cero. La falta de datos pertenecientes a la magnitud de RF1 es debido a que las primeras respuestas tendían a oscilar entre los primeros minutos de las sesiones experimentales, y se observaron, a lo largo de esas sesiones, periodos de conductas operantes y alimentación de reforzadores positivos, y periodos de alimentación de comida libre.
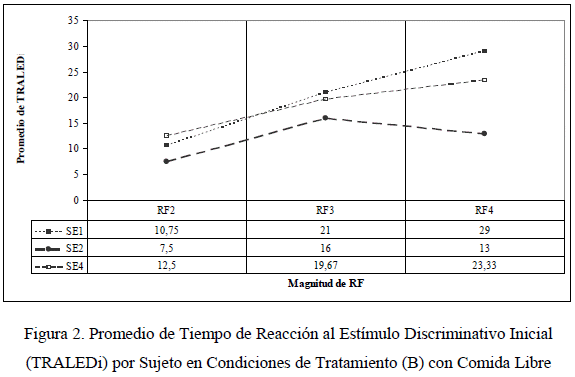
Se aprecia un aumento gradual en los TRALEDi mientras aumenta la magnitud de RF. Aunque la tendencia del segundo sujeto experimental parece desviarse en RF4, vale una aclaración. En dicha magnitud el sujeto respondió de acuerdo al criterio conductual en dos ocasiones. En la primera, emitió RF4 a los 25 minutos, mientras que en la segunda, el TRALEDi fue de 1 minuto, es decir, al primer minuto de la sesión experimental. Si se tomara a la respuesta en el primer minuto como de pura coincidencia, el promedio hubiera sido de 25 minutos, aproximadamente. A no ser así, la figura parece desviarse completamente de la posible tendencia generalizable entre los tres sujetos experimentales. A pesar de ello, se observa que mientras mayor sea la magnitud de RF, mayor la latencia de la primera respuesta (TRALEDi) de CFL.
DISCUSIÓN
La presente investigación tuvo el fin de determinar en qué medida el incremento de la magnitud requerida en un programa de RF influye sobre la conducta de CFL en organismos inferiores (mus músculus). Los resultados demuestran una clara tendencia de disminución de CFL mientras se aumenta la magnitud de RF. Traducido como esfuerzo, estos resultados no sólo confirman estudios previos que el nivel de CFL disminuye mientras mayor el esfuerzo (Carder & Berkowitz, 1970; Inglis, Forkman & Lazarus, 1997; Inglis, Langton, Forkman & Lazarus, 2001), sino demuestran cómo el aumento gradual de esfuerzo influye sobre la disminución de CFL.
Se observaron tendencias similares en tres de los cuatro sujetos experimentales mientras mayor la magnitud de RF (esfuerzo). Mientras el esfuerzo era mínimo, el CFL se mantuvo relativamente elevado. En contraste, cuando se incrementaba el esfuerzo el CFL era menor. Por tanto, se puede adjudicar mayor valor al estudio de Inglis et al. (2001) que los niveles de CFL son menores cuando se exige mayor esfuerzo.
A pesar de ello, no se puede aclarar contundentemente si es que RF5 representa menor o mayor esfuerzo en organismos inferiores (mus músculus) en situaciones sin comida libre. Lo que se puede confirmar es el nivel de esfuerzo bajo el cual el nivel de CFL es nulo en organismos inferiores. Tanto en la primera parte, cómo posterior a la reorientación experimental, los sujetos no emitieron la conducta de CFL en la magnitud de RF5. Se concluye que esta magnitud conlleva demasiado esfuerzo para mantener respuestas de CFL, por lo que los organismos “prefieren” alimentarse exclusivamente de comida libre para así ahorrar energía (Menzel, 1991; Jensen, 2000).
Asimismo, se ha establecido que la tendencia de descenso de los promedios de respuesta de CFL al incrementar gradualmente la magnitud de RF es significativo. Lo que se observó indica que sin importar el promedio de respuestas emitidas durante un programa de reforzamiento continuo (RF1), cuando el programa se vuelve intermitente (RF2-5), el promedio de respuestas se agrupan en una tendencia de descenso muy similar hasta llegar a cero en la magnitud de RF5. Por lo que se concluye que no importa cuanta exposición un organismo haya tenido con respecto a una respuesta operante, cuan consolidada esté dicha conducta, ni cuantas respuestas hayan emitido en un programa continuo, la tendencia de descenso de respuesta del CFL mientras se incrementa la magnitud de RF se asemeja en los organismos inferiores (mus músculus).
Estos resultados rebaten el argumento de Tarte y Rasmussen (1979) que un periodo mayor de entrenamiento sobre el operando daría lugar a la preferencia por la respuesta operante sobre la comida libre. A pesar de periodos extendidos de consolidación (o de entrenamiento), no se observaron cantidades de respuestas representativas de CFL, demostrando que cuando la magnitud (o esfuerzo) de una conducta operante es elevada la ampliación del periodo de consolidación durante condiciones sin comida libre no incrementa la probabilidad que el CFL se presente.
Quizá mayores periodos de entrenamiento sirvan más en programas de reforzamiento continuo que en programas intermitentes.
Adicionalmente a dichos resultados, se observó que mientras la magnitud de RF (nivel de esfuerzo) incrementa gradualmente, el promedio de respuestas disminuye y consecuentemente la cantidad de ingesta de alimento libre (g.) aumenta. Aunque dichas tendencias positiva y negativa relacionadas con el aumento de magnitud podrían asumirse a priori, los resultados claramente permiten concluir sobre la manera que estas tendencias se presentan al escalonar de una magnitud a la siguiente.
Cabe recalcar que existen ramificaciones adicionales inesperadas, pero de interés. Por una parte, mientras que los promedios de respuestas en la condición de Tratamiento (B) disminuyen al incrementar el esfuerzo, los promedios aumentan o se mantienen durante la condición de Línea Base (A). Muchas veces los sujetos en la condición de Tratamiento (B) lograron alimentarse hasta con un doble de la cantidad que lo hacían durante la condición de Línea Base (A). Este incremento puede explicarse según las afirmaciones de Rutter y Nevin (1990) que los animales tratan de mantener una misma cantidad de ingesta aunque hayan cambios ambientales que alteren la accesibilidad a la misma. Así, se puede concluir que para lograr mantener la misma cantidad de alimento ingerido en la condición de Línea Base (A) que en la condición de Tratamiento (B) los organismos emitieron mayores cantidades de respuestas para recibir mayores cantidades de reforzadores positivos (pellets), consecuencia debida a la experiencia con el CFL.
Asimismo, la cantidad total de alimento consumido (g.) en la condición de Tratamiento (B), sumando el alimento consumido como reforzador con la comida libre, tiende a ser mayor que la cantidad de alimento consumido (g.) en la condición de Línea Base (A), en la mayoría de las magnitudes de RF estudiadas. Dicha tendencia fue demostrada inicialmente por Powell (1974).
Otra observación adicional es referida al tiempo de reacción al estímulo discriminativo inicial (TRALEDi). Los resultados indican que cuando el esfuerzo es mínimo (reforzamiento continuo), la primera respuesta de CFL se emite tempranamente. Interesantemente, al incrementar la magnitud de RF (reforzamiento intermitente), el tiempo en que el sujeto emite su primera respuesta operante demora, el cual se extiende gradualmente al incrementar el esfuerzo. Es decir, los organismos emiten el CFL cada vez más tardíamente cuando el esfuerzo aumenta.
A este detalle se puede agregar el hecho que la mayor parte de las respuestas de CFL en total emitidas durante una sesión se presentan en aglomeraciones de trenes de respuesta y no respuestas aisladas. Cuando el sujeto responde sobre el operando lo hace en varias ocasiones seguidas. En global, dichos resultados afinan la postulación de que en un inicio los organismos prefieren la ingesta libre, y que subsiguientemente realizan el CFL (Inglis, Forkman & Lazarus, 1997). Esto se debe a que cuando un organismo se encuentra (por ejemplo, de alimento) recién dará importancia a reducir la incertidumbre ambiental (Inglis, Langton, Forkman & Lazarus, 2001).
A pesar de los alcances y la novedad de algunos resultados, existen ciertas limitaciones en la presente investigación a tomarse en cuenta. Por una parte, Powell (1974) demostró que el valor de reforzador (cualidad y cantidad) influye sobre el CFL. Quizá el tamaño de los pellets utilizados no fue lo suficientemente reforzante como para mantener la conducta operante, y en lugar de trabajar optaron por el alimento libre.
Otro factor que pudo haber influido en los resultados es la calidad de albergue de los sujetos experimentales. El estudio de Coburn y Tarte (1976) indicó que el CFL disminuye en organismos albergados en ambientes favorables e incrementa en organismos albergados en ambientes empobrecidos. Debido a que los sujetos tuvieron acceso ilimitado al bebedero de agua y a una rueda, y fueron expuestos a una interacción con el investigador por lo menos de una hora diariamente, se considera que fueron albergados en un ambiente favorable. Aunque los resultados no puedan afirmar dicha limitación, es razonable considerarlo al analizar los resultados.
Por otra parte, la presente investigación consideró la metodología de estudios previos de trabajar en espacios experimentales controlados y midiendo el CFL como una conducta operante de presionar un operando. Es posible que la conducta operante utilizada de presionar un operando quizá no represente una conducta naturalmente dentro del repertorio conductual de un roedor (mus músculus) relacionada con una conducta de forrajeo. Elegir una conducta operante similar a una conducta que se presenta naturalmente en su predisposición biológica (Domjan, 1999) cuando el organismo busca alimento podría representar un verdadero nivel de CFL.
Finalmente, la duración de cada condición del diseño experimental utilizado (diseño reversible A-B-A-B) quizá conlleve ciertas limitaciones. La duración de cada condición se determinó de antemano, en lugar de aplicar cada condición hasta encontrar una estabilidad conductual.
Ya que el nivel de esfuerzo es relativo a diferentes especies, se recomienda que se replique la experimentación realizada con otras especies, controlando el incremento gradual de las magnitudes de RF para determinar si es que la tendencia observada es similar en otras especies. Cada especie presenta diferentes características comportamentales. Es posible que naturalmente ciertas especies tiendan a holgazanear en lugar de trabajar, por lo que se discrepen algunos resultados cuando se comparan inter-especie. Si se logra replicar dicha tendencia habrá mayor posibilidad de generalizar este resultado a la teoría del CFL.
Además, se recomienda la reformulación del mismo diseño experimental, diseño reversible A-B-A-B, o utilizar otro diseño favorable para estudiar esta tendencia. Se recomienda que la siguiente aplicación se enfoque en un determinante conductual, buscando llegar a una constancia conductual en cada condición antes de considerar pasar a la siguiente condición.
Por otra parte se recomienda estudiar el elemento novedoso observado en la presente investigación, el TRALEDi. Ya que se encontró una tendencia de aumento de este tiempo de reacción mientras mayor el nivel de esfuerzo, sería conveniente considerarlo como un elemento adicional al estudio del CFL.
La teoría y las suposiciones sobre el fenómeno del CFL aun son jóvenes a pesar de de la amplitud experimental. Mediante la aplicación de la presente investigación se ha logrado confirmar algunos conceptos existentes del CFL y establecer otros conceptos que todavía se encuentran incompletos o desconocidos.
REFERENCIAS
Bachá M., G. (2002). Adquisición de una respuesta operante en presencia de una fuente alterna: Contrafreeloading. Revista Mexicana de Análisis de la Conducta, 28(1), 91-104. [ Links ]
Bautista, L. M. (1997). Estrategias de Alimentación: ¿Es posible reducir el comportamiento a tiempo y energía?. Recuperado el 4 de julio de 2007 en http://usuarios.lycos.es/lmbautista_2/EtoloGUIA.html [ Links ]
Bean, D., Mason, G. J., & Bateson, M. (1999). Contrafreeloading in starlings: Testing the information hypothesis. Recuperado el 5 de diciembre de 2006 en http://www.ingentaconnect.com/content/brill/beh/1999/00000136/F0020010/art0000 1;jsessionid=3sd56g8acq1l8.alice [ Links ]
Carder, B., & Beckman, G. C. (1975). Limitations of “Container Neophobia” as an explanation of rats' responding for food in the presence of Free Food. Recuperado el 16 de mayo de 2007 en http://www.sciencedirect.com/science?_ob=ArticleURL&_udi=B7MD5-4DJ4PW153&_user=10&_coverDate=05%2F31%2F1975&_alid=577301789&_rdoc=9&_fmt=summary&_orig=search&_cdi=23221&_sort=d&_docanchor=&view=c&_ct=10& _acct=C000050221&_version=1&_urlVersion=0&_userid=10&md5=28e204ce7285 b013aceb14290db40ea0
Carder, B., & Berkowitz, K. (1970). Rats' Preference for Earned in Comparison with Free Food. Recuperado el 18 de mayo de 2007 en http://www.sciencemag.org/cgi/content/abstract/167/3922/1273 [ Links ]
Clement, T. S., Feltus, J. R., Kaiser, D. H., & Zentall, T. (2000). “Work ethic” in pigeons: Reward value is directly related to the effort or time required to obtain the reward. Psychonomic Bulletin & Review 2000, 7(1), 100-106.
Coburn, J. F., & Tarte, R. D. (1976). The Effect of Rearing Environments on the Contrafreeloading Phenomenon in Rats. Journal of the Experimental Analysis of Behavior, 26(2), 289-294. [ Links ]
Day, J. E. L., Kyriazakis, I., & Rogers, P. J. (1998). Food Choice and Intake: Towards a unifying framework of learning and feeding motivation. Nutrition Research Reviews, 11, 25-43. [ Links ]
Domjan, M. (1999). Principios de Aprendizaje. México: International Thomson. [ Links ]
Forkman, B. (1996). The Foraging Behaviour of Mongolian Gerbils: A behavioural need or a need to know?. Recuperado el 15 de febrero de 2007 en http://cat.inist.fr/?aModele=afficheN&cpsidt=3060638 [ Links ]
Gilbert-Norton, L., & Gee, P. (2005). Captive Birds and Contrafreeloading: The choice to work. Recuperado el 22 de marzo de 2007 en http://www.wcs.org/media/file/ICEEProceedingsFinal.pdf [ Links ]
Inglis, I. R., & Langton, S. (2006). How an Animal's Behavioural Repertoire changes in response to a Changing Environment: A stochastic model. Recuperado el 15 de febrero de 2007 en http://www.ingentaconnect.com/search/article?title=information+primacy+hypothesis&title_type=tka&year_from=1998&year_to=2007&database=1&pageSize=20&index=1 [ Links ]
Inglis, I. R., Forkman, B., & Lazarus, J. (1997). Free food or earned food? A review and fuzzy model of contrafreeloading. Animal Behaviour, 53(6), 1171-1191. [ Links ]
Inglis, I. R., Langton, S., Forkman, B., & Lazarus, J. (2001). An Information Primacy Model of Exploratory and Foraging Behaviour. Animal Behaviour, 62(3), 543-557. [ Links ]
Jensen, G. D. (1963). Preference for bar-pressing over free-loading as a function of number of rewarded presses. En Osborne, S. R., & Shelby, M. (1975). Stimulus Change as a Factor in Response Maintenance with Free Food available. Journal of the Experimental Analysis of Behavior, 24(1), 17–21.
Jensen, P. (2000). Natural Behaviour and Behavioural Needs of Farm Animals. Recuperado el 5 de diciembre de 2006 en http://www.agriculture.de/acms1/conf6/ws5abehav.htm [ Links ]
Kerlinger, F. N., & Lee, H. B. (2002). Investigación del Comportamiento: Métodos de investigación en ciencias sociales. México: McGraw Hill/Interamericana. [ Links ]
Lind, D. A., Marchal, W. G., & Mason, R. D. (2004). Estadística para Administración y Economía. México: Alfaomega Grupo. [ Links ]
Lindqvist, C., & Jensen, P. (2003). Contrafreeloading decreases with Age and Social Isolation in Red Jungle Fowl and White Leghorn Layers. Recuperado el 23 de marzo de 2007 en http://www.bs.izs.it/Referenza/Benessere/Articoli/Quaderno%2054.pdf [ Links ]
Lindqvist, C., Zimmerman P., & Jensen, P. (2006). A note on contrafreeloading in broilers compared to layer chicks. Applied Animal Behaviour Science, 101(1-2), 161–166.
Martin, G., & Pear, J. (1999). Modificación de Conducta: Qué es y cómo aplicarla; Quinta Edición. Madrid: Prentice Hall Iberia. [ Links ]
McGowan, R., Newberry, R., Robbins, C., & Alldredge, R. (2005). Contrafreeloading in Grizzly Bears: Implications for environmental enrichment. Recuperado el 22 de marzo de 2007 en http://www.wcs.org/media/file/ICEEProceedingsFinal.pdf [ Links ]
Menzel, Jr., E. W. (1991). Chimpanzees (Pan Troglodytes): Problem Seeking versus the Bird-in-hand, Least-effort Strategy. Primates, 32(4), 497-508. [ Links ]
Mitchell, D., Becnel, J. R., & Blue, T. (1981). The Neophobia-Optimality Explanation of Contrafreeloading Rats: A Reassessment. Recuperado el 16 de mayo de 2007 en http://www.sciencedirect.com/science?_ob=ArticleURL&_udi=B7MD6-4DJ4NPH2V&_user=10&_coverDate=08%2F31%2F1981&_alid=577286530&_rdoc=9&_fmt=summary&_orig=search&_cdi=23222&_sort=d&_docanchor=&view=c&_ct=9&_ acct=C000050221&_version=1&_urlVersion=0&_userid=10&md5=68ae4a5545b31 384fa45f06abb7bcf33 [ Links ]
Mitchell, D., Fish, R. C., & Calica, D. R. (1981). Rats Respond for Food in the Presence of Free Food: How free is the “free” food?. Recuperado el 16 de mayo de 2007 en http://www.sciencedirect.com/science?_ob=ArticleURL&_udi=B6WMT-4D62P0X 6V&_user=10&_coverDate=05%2F31%2F1982&_alid=577296605&_rdoc=9&_fmt =summary&_orig=search&_cdi=6943&_sort=d&_docanchor=&view=c&_ct=12&_ acct=C000050221&_version=1&_urlVersion=0&_userid=10&md5=e8e87fa9be273 927d234aa393f46e359
Mitchell, D., Scott, D. W., & Williams, K. D. (1973). Container Neophobia and the rat's preference for Earned Food. Recuperado el 16 de mayo de 2007 en http://www.sciencedirect.com/science?_ob=ArticleURL&_udi=B7MD5-4GPX16K 8&_user=10&_coverDate=11%2F30%2F1973&_alid=577313541&_rdoc=4&_fmt= summary&_orig=search&_cdi=23221&_sort=d&_docanchor=&view=c&_ct=5&_ac ct=C000050221&_version=1&_urlVersion=0&_userid=10&md5=52876cf50a3ad58 4d90be4cd9ed11645 [ Links ]
Neuringer, A. J. (1969). Animals Respond for Food in the Presence of Free Food. Recuperado el 25 de mayo de 2007 en http://www.sciencemag.org/cgi/content/abstract/166/3903/399 [ Links ]
Osborne, S. R., & Shelby, M. (1975). Stimulus Change as a Factor in Response Maintenance with Free Food available. Journal of the Experimental Analysis of Behavior, 24(1), 17–21.
Powell, R. W. (1974). Comparative Studies of the referent of Free versus Response-Produced Reinforcers. Animal Learning & Behavior, 2(3), 185-188. [ Links ]
Robertson, L. C., & Anderson, S. C. (1975). The Effects of Differing Type and Magnitude of Reward on the Contrafreeloading Phenomenon in Rats. Animal Learning & Behavior, 3(4), 325-328. [ Links ]
Rutter, S., & Nevin, J. A. (1990). Long-term Contrafreeloading in Rats during Continuous Sessions. Bulletin of the Psychonomic Society, 28(6), 556-558. [ Links ]
Schütz, K. E., & Jensen, P. (2001). Effects of Resource Allocation on Behavioural Strategies: A Comparison of Red Junglefowl (Gallus gallus) and Two Domesticated Breeds of Poultry. Ethology, 107(8), 753-765. [ Links ]
Singh, D. (1970). Preference for Bar Pressing to obtain reward over Freeloading in Rats and Children. En Inglis. I. R., Forkman, B., & Lazarus, J. (1997). Free food or earned food? A review and fuzzy model of contrafreeloading. Animal Behaviour, 53(6), 1171-1191. [ Links ]
Skinner, B. F. (1969). Contingencies of Reinforcement: A Theoretical Analysis. Appleton-Century-Crofts: Nueva York. [ Links ]
Talling, J. C., Inglis, I. R., van Driel, K. S., Young, J., & Giles, S. (2002). Effect of Hunger on Starlings' Preferences for Food Sources associated with Variability or Uncertainty. Recuperado el 15 de febrero de 2007 en http://www.ingentaconnect.com/content/brill/beh/2002/00000139/00000009/art00009 [ Links ]
Tarte, R. D. (1981). Contrafreeloading in Humans. En Inglis. I. R., Forkman, B., & Lazarus, J. (1997). Free food or earned food? A review and fuzzy model of contrafreeloading. Animal Behaviour, 53(6), 1171-1191. [ Links ]
Tarte, R. D., & Rasmussen, C. T. (1979). Training Effects on Contrafreeloading in Rats housed in Operant Chambers. Animal Learning & Behavior, 7 (1), 69-74. [ Links ]
Vaz-Ferreira, R. (1984). Etología: El Estudio Biológico del Comportamiento Animal. Washington, D.C.: Secretaría General de la Organización de los Estados Americanos. [ Links ]
Vera C., J. L. (2002). Estrategias Alimentarias de los primeros Homínidos. En Martinez C., J. & Veá, J. J. (2002). Primates: Evolución, Cultura y Diversidad. México: Centro de Estudios Filosóficos, Políticos y Sociales Vicente Lombardo Toledano. [ Links ]
Wackerly, D. D., Mendenhall III, W., & Shaeffer, R. L. (2003). Estadística Matemática con Aplicaciones. México: Thomson. [ Links ]
Wilde, M. C., & Otondo, M. E. (1990). Manual Básico de Laboratorio: Análisis Experimental de la Conducta. La Paz, Bolivia. Universidad Católica Boliviana. Páginas 12-24. [ Links ]

