











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
Permalink
Introducción
En México y el mundo, el queroseno, es un derivado del petróleo, una mezcla insoluble de hidrocarburos alifáticos y aromáticos tóxicos ambientales1, como el benceno, n-hexano, tolueno, xileno, al igual que otros aromáticos policíclicos como el naftaleno y n-propilbenceno, susceptibles al ataque microbiano de acuerdo con el tipo de metabolismo que poseen, así como de las condiciones ambientales que existen en la naturaleza o en el laboratorio1-3. Cuando el queroseno impacta ambientes naturales como el suelo, induce la selección de una amplia diversidad de microorganismos, que pueden ser procariotas con géneros y especies de bacterias comunes en ese ambiente. Además de los eucariotas heterotróficos, aerobios como los hongos filamentosos y algunos géneros de levaduras que tienen la capacidad de biodegradar el queroseno al utilizarlo como única fuente de carbono y energía4-6. Según Prince et al.7 se conocen más de 150 géneros y especies de bacterias que degradan queroseno del tipo: Rhodococcus aetherivorans y R. wratislaviensis8, Streptomyces spp.9, Pseudomonas aeruginosa10,11Vibrio, Brevibacterium, Achromobacter, Mycobacterium y Bacillus spp.12-14. En contraste, una menor diversidad de hongos filamentosos que se han reportado15,16 cuyos ejemplos más representativos son: Aspergillus sp., Penicillum sp, y Trichoderma asperellum que biodegradan hidrocarburos aromáticos17-19. Mientras que existe interesante información relacionada con géneros y especies de levaduras del ambiente que han sido menos investigadas para la eliminación de hidrocarburos alifáticos y aromáticos cuando los utilizan como única fuente de carbono y energía20,21. Así como para la síntesis de proteína de origen unicelular como alimento para humanos y animales22,23. Un ejemplo, en la naturaleza es el género Candida24,25 en la naturaleza, al igual que Cryptococcus, Pichia, Yarrowia26-28 así como: Meyerozyma, Rhodotorula, Wickerhamia y Rhodosporidium6. Por lo anterior existe la evidencia de que el género de levadura Saccharomyces tiene potencial para explotarse en la síntesis de proteína de origen unicelular para la elaboración de alimento de excelente calidad nutricional de bajo costo22. Así como para la recuperación de ambientes impactados por mezclas de hidrocarburos, como el queroseno mediante bioaumentación proceso en el cual Saccharomyces sería fundamental para lograrlo27. Al respecto se ha reportado que especies de Saccharomyces tienen la capacidad bioquímica para la utilización de fracciones hidrocarburos de 12 a 16 carbonos análogas al queroseno para mineralizarlos hasta CO2 y agua28. Por lo anterior el objetivo de este trabajo fue analizar la capacidad de Saccharomyces exiguus para usar queroseno como única fuente de carbono y energía.
Materiales y métodos
Origen de Saccharomyces exiguus. En esta investigación partió de la premisa de que S. exiguus al igual que otros géneros y especies de levaduras son parte de la microbiota que existe en el suelo de pozos petroleros de PEMEX en Altamira, Tamaulipas, México, y del mundo26. El suelo contaminado por derivados del petróleo se analizó en el Laboratorio de Microbiología Industrial y Suelo de la Facultad de Ciencias Biológicas (FCB), Universidad Autónoma de Nuevo León (UANL) y de Microbiología Ambiental del Instituto de Investigaciones Químico Biológicas (IIQB) de la Universidad Michoacana de San Nicolás de Hidalgo (UMSNH). Para ello 1.0 g de suelo se sembró en 50 mL de un medio mineral con la siguiente composición química (g(L-1): queroseno al 5 % (v/v) esterilizado por filtración con una membrana Millipore 0.2 µm, NH4Cl 2.0, NaCl 4.0, MgSO4 0.5, K2HPO4 0.5, KH2PO4 1.0, a pH ajustado a 5.5, que se incubó a 30 °C/15 días en agitación a 200 rpm, al aparecer la turbidez se sembró en agar queroseno al 5 % que se incubo a 30 °C/15 días con el crecimiento de colonias de S. exiguus, que se resembraron en agar Sabouraud (AS) con la siguiente composición química (g(L-1): polipeptona 10.0, glucosa 40.0, agar 18.0, un pH ajustado a 5.6 que se incubó a 30 °C/40 h, entonces estas colonias se suspendieron en solución salina (NaCl 0.85 %) mezclado con el detergente La CoronaMR 0.01 % (SSD): la levadura se agitó a 200 ppm/2 h, después se centrifugó, se suspendió en SSD al 0.85 % e inocularon en matraces nefelométricos: en donde como control absoluto (CA) se empleó un matraz solo con SSD al 0.85 % ajustado a 0 en una absorbancia de 440 unidades Klett (UK) en un fotocolorímetro Klett Sumerson con filtro rojo (650 nm): de la suspensión de S. exiguus se tomaron 5 mL e inocularon en 50 mL queroseno liquido mineral 5 %, con NH4Cl al 1.2 % y 50 ppm de extracto de levadura a pH de 5.56.
Cinética de crecimiento de S. exiguus en queroseno. La levadura S. exiguus se reprodujo en queroseno al 5 % con NH4Cl al 1.2 % y liquido de remojo de maíz (LRM) al 0.025 % (producto de Maíz, Guadalajara, Jal. México), o bien con extracto de malta 50 ppm (Merck), y/o extracto de levadura 50 ppm (Merck). Entonces S. exiguus se incubó a 30 °C/4 días/200 rpm por triplicado. En esta fase se emplearon diferentes variables para medir el crecimiento de S. exiguus en queroseno: i) densidad óptica en un fotocolorímetro Klett Sumerson a 650 nm cada 24 h, como blanco queroseno mineral al 5 % sin S. exiguus, ii) peso seco con membrana Millipore de 0.2 µm y 13 mm de diámetro (Millipore Corporation, USA), que se tararon en un horno a 110 °C, para filtrar 0.4 mL de la suspensión celular de S. exiguus, nuevamente se tararon, la diferencia en el peso de la membrana con células y peso de la membrana sin células se expresó en g/L para graficar en gramos (g) de S. exiguus en relación al tiempo, iii) concentración de proteína: por el método de Lowry como medición indirecta del crecimiento de S. exiguus con el reactivo Folin-Ciocalteu, para ello se tomaron 0.4 mL de sobrenadante del queroseno al 5 % liquido mineral a intervalos por 24 h, se centrifugó a 2500 rpm/15 min, se midió en un espectrofotómetro Modelo/620 Coleman Junior II (Beckman Instruments, Inc.). Los datos experimentales se validaron con el programa estadístico ANOVA/Tukey HSD P<0.05 % con Statgraphics Centurion29, iv) la identificación de S. exiguus y otras análogas de géneros fue de acuerdo al manual de Lodder30 y de Phaff et al.31, basados en las características morfológicas y fisiológicas indicadas en la Tabla 2 con el empleo de los siguientes medios de cultivo, agar extracto de malta (AEM) (g/L): extracto de malta 3.0, extracto de levadura 3.0, peptona 5.0, glucosa 10.0, agar 18.0, a pH 5.0, caldo malta levadura (CML) (g/L): extracto de malta 3.0, extracto de levadura 3.0, peptona 5.0, glucosa 10.0 y pH a 5.0, caldo Gorodkowa (g/L): glucosa 2.5, NaCl 5.0, extracto de carne 10.0 y pH de 6.5, agar acetato de Fowell (g/L): acetato de sodio 0.5, agar 2.0 a pH 6.5, etanol de Starkey modificado (g/L): etanol 0.05, K2HPO4 0.025, MgSO4*H2O 0.025 a pH 6.5, Wickerham* (g/L): extracto de levadura 4.5, peptona 7.5 y pH a 6.5, y Wickerham** (g/L): KH2PO4 0.15, MgSO4*H2O 0.5, NaCl 0.1, CaCl2 0.1, dextrosa 10.0, NH4Cl 0.5 y pH de 6.5 como referencia de identificación de las levaduras aisladas se usaron cepas de levaduras obtenidas comercialmente32.
Análisis cromatográfico del crecimiento de S. exiguus en queroseno. El queroseno remanente que S. exiguus no utilizo como fuente de carbono y energía, se separó del medio mineral por centrifugación, el cual se analizó en un cromatógrafo de gases Beckman GC 72-5. Para demostrar y cuantificar los hidrocarburos alifáticos y aromáticos del queroseno que fueron consumidos por S. exiguus. Para ello se utilizó una mezcla de estándares: C10, C12, C13, C14, C16 (Varían) se analizaron por la elución de los picos de la cromatografía de gases antes y después del crecimiento de S. exiguus en el queroseno33.
Resultados

Figura 1 Medición indirecta de la capacidad de S. exiguus para utilizar el queroseno al 5 %, como fuente de carbono y energía NH4 Cl al 1.2 % con diversos estimuladores o factores de crecimiento
En la Figura 1, se observa el crecimiento de S. exiguus en queroseno al 5 % como única fuente de carbono y energía enriquecido con 50 ppm de extracto de levadura: en la que se indica que la fase de adaptación inició antes de las 8 h, mientras que la logarítmica comenzó las primeras 12 h para concluir a las 48 h, de ahí la estacionaria hasta las 120 h. Análogo cuando S. exiguus creció en el queroseno al 5 % con el NH4Cl al 1.2 % como fuente mineral de nitrógeno, enriquecido con 50 ppm de extracto de levadura.
En la Figura 2, se muestra el crecimiento de S. exiguus en queroseno al 5 %, sin el NH4Cl al 1.2 %, enriquecido con 50 ppm de extracto de levadura, ahí se registró la mayor cantidad de proteína celular de 39 μg/mL en las primeras 24 h, con un incremento de hasta 190 μg/mL a las 120 h. Mientras que cuando S. exiguus se le dio como fuente de nitrógeno mineral en la forma de NH4Cl al 1.2 %, que enriquecido con 50 ppm de extracto de levadura se registraron 36 μg/mL de proteína en las 24 h y 153 μg/mL a las 120 h evidencia indirecta de que S. exiguus utilizo el queroseno como fuente de carbono y energía.

Figura 2 Concentración de proteína de S. exiguus cuando utilizo del queroseno al 5 %, como fuente de carbono y energía NH 4 Cl al 1.2 % con diferentes factores o estimuladores de crecimiento
En la Figura 3, se muestra la medición indirecta del crecimiento mediante el peso seco de S. exiguus en queroseno al 5 % cuando lo utilizo como fuente de carbono y energía, sin el NH4Cl al 1.2 % pero enriquecido con 50 ppm de extracto de levadura, ahí se registraron 1.35 g/L en las primeras 48 h con un aumento de hasta 1.45 g/L a las 120 h, lo que indica que el extracto de levadura también es una fuente de nitrógeno orgánico, limitada en disponibilidad para S. exiguus, por la baja concentración que se agregó al medio mineral.
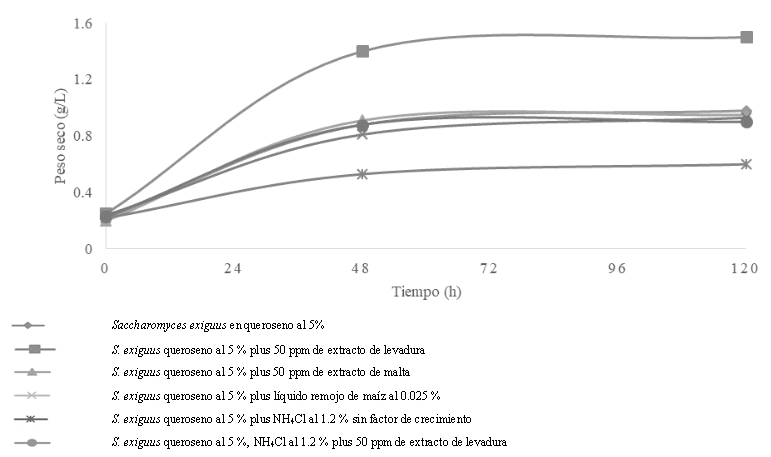
Figura 3 Peso seco de S. exiguus al utilizar el queroseno al 5 %, como fuente de carbono y energía con el NH 4 Cl al 1.2 % y diversos factores o estimuladores de crecimiento
En la Tabla 1, se muestra el crecimiento de S. exiguus en queroseno al 5 % cuando lo utilizo como fuente de carbono y energía con el NH4Cl al 1.2 %, enriquecido con el extracto de levadura, donde se registró el mayor rendimiento o producción celular de hasta 1.15 g/L con una máxima velocidad de crecimiento de 0.081 h-1, con un menor tiempo de duplicación de 8.5 h, que causo una disminución de pH hasta 3.5. Lo anterior que mostro que la composición química del medio mineral con el queroseno al 5 %, el NH4Cl, el extracto de levadura como factor de crecimiento fueron la mejor condición nutricional para la biodegradación del queroseno.
Tabla 1 Crecimiento de S. exiguus al utilizar queroseno al 5%, como fuente de carbono y energía NH 4 Cl con y sin factor o estimulador de crecimiento
| Saccharomyces exiguus en queroseno al 5%. | X (g/L) | µexp (h-1) | t.d (h) | pH final |
|---|---|---|---|---|
| NH4Cl al 1.2 % sin factor de crecimiento | 0.73c** | 0.049b | 14c | 4.3a |
| NH4Cl al 1.2 % + extracto de levadura 50 ppm | 1.15a | 0.081a | 8.5d | 3.5b |
| NH4Cl al 1.2 % + extracto de malta 50 ppm | 0.87b | 0.047b | 14.5c | 4.0a |
| NH4Cl al 1.2 % + líquido de remojo de maíz al 0.025 % | 0.72c | 0.031c | 22.0b | 3.8a |
| Sin NH4Cl al 1.2 %, ni factor de crecimiento | 0.28e | 0.022d | 30a | 3.9a |
| Sin NH4Cl al 1.2 % con extracto de levadura 50 ppm | 0.63d | 0.038c | 18c | 3.5b |
Condiciones: Inoculo 10 % (v/v), agitación 200 rpm, 30 °C, pH inicial 5.5, X = rendimiento celular, µ exp = velocidad de crecimiento, t.d. = tiempo de duplicación,
*n= 3. **valores con diferente letra indican diferencia estadística según ANOVA/Tukey (P ≤ 0.05).
En la Figura 4, se describe el perfil cromatográfico del queroseno al 5 % sin inocular con S. exiguus usado como control absoluto, donde se registró el mayor número de picos de elución que indican los hidrocarburos alifáticos del tipo iso-alcanos, según el tiempo de retención similares a los n-alcanos de los estándares empleados como referencia34,35.
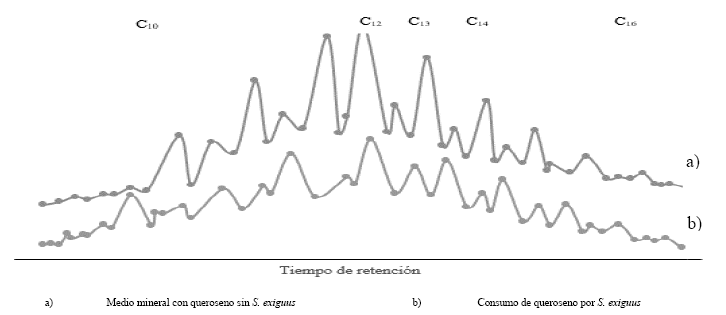
Figura 4 Análisis de la utilización por S. exiguus de los hidrocarburos del queroseno al 5 %, como fuente de carbono y energía, el NH 4 Cl al 1.2 % y el extracto de levadura a 30°C/200 rpm
En la Tabla 2, se muestran las características morfológicas y fisiológicas de los géneros y especies de levaduras aisladas de diferentes pozos petroleros de PEMEX de la ciudad Altamira, Tam., México, que utilizan el queroseno como única fuente de carbono y energía. Las que de acuerdo con las características morfológicas y fisiológicas de los géneros levaduras en los medios de cultivo empleados para la identificación como: AS, caldo malta levadura (CML), Gorodkowa, agar acetato de Fowell, Starkey´s etanol, Wickerham y las descritas en el Manual de Lodder30 y de Phaff et al.31, en relación al género Aureobasidium, con células ovales de gemación polar, sin ascosporas, con micelio septado, blastosporas y clamidiosporas, que forma un tipo de pellet en cultivos líquidos, no fermentó la glucosa, aunque en anaerobiosis redujo los NO3 - a NO2 -.
Tabla 2 Características morfológicas y bioquímicas de los géneros de las levaduras que utilizan el queroseno como fuente de carbono y energía
| Características morfológicas y fisiológicas | Aureobasidium sp. | Rhodotorula sp. | S. exiguus |
|---|---|---|---|
| Tipo de gemación | Polar | Multilateral | Multilateral |
| Crecimiento en agar extracto de malta-extracto de levadura (AEML) | Abundante | Abundante | Abundante |
| Formación de ascas | - | - | 1 a 4 por asca |
| Fermentación de glucosa | + | - | - |
| Asimilación de NO3 (nitratos) | - | - | - |
| Síntesis de pigmentos | Verde oliváceo a negro | Naranja | - |
| Formación de película y/o pellet | +* | + | - |
Ausencia de crecimiento (-), crecimiento (+), *produce una película o pellet.
Discusión
La Figura 1 muestra que S. exiguus creció sin problemas a pesar de que la concentración del queroseno fue de un 5 %, sin el NH4Cl al 1.2 %, pero enriquecido con el extracto de levadura que contiene vitaminas del complejo B, en concentración suficiente para que S. exiguus haya utilizado los hidrocarburos alifáticos y aromáticos del queroseno como única fuente de carbono y energía por la vía aerobia, dependiente del O2, con las enzimas de la cadena respiratoria que son dependientes de la concentración de vitaminas del complejo B del extracto de levadura32,36,37. Al respecto se reporta que diferentes géneros y especies de levaduras. Candida albicans tienen la capacidad de metabolizar hidrocarburos aromáticos policíclicos similares a los del queroseno. En tanto que S. exiguus tuvo un pobre crecimiento en queroseno al 5 % con el NH4Cl al 1.2 % como fuente de mineral de nitrógeno, en ausencia de las vitaminas del complejo B, factor de crecimiento, en forma del extracto de levadura, que le impidió a S. exiguus activar las enzimas para metabolizar los hidrocarburos alifáticos del queroseno33. La Figura 2 muestra el aumento en la concentración de proteína de S. exiguus cuando utiliza el queroseno al 5 % como única fuente de carbono y energía en el medio de cultivo, favorecido con la fuente inorgánica de nitrógeno como el NH4Cl al 1.2 %, y enriquecido con el extracto de levadura con las vitaminas del complejo B, activaron las enzimas necesarias para la oxidación de los hidrocarburos alifáticos del queroseno por S. exiguus por la vía de beta oxidación ruta de bioquímica de degradación para el queroseno23,34. En contraste con el crecimiento de S. exiguus en el queroseno al 5 % con el NH4Cl al 1.2 %, sin el factor de crecimiento, ahí se registró una concentración de proteína con valores que fluctuaron entre 25-56 μg/mL al final de la fase estacionaria a las 120 h, lo que indica que la ausencia del extracto de levadura ralentizo e inhibió capacidad de S. exiguus para utilizar los hidrocarburos alifáticos del queroseno como fuente de carbono y energía33,37. En la Figura 3 se muestra el peso seco de S. exiguus cuando utilizo los hidrocarburos alifáticos y aromáticos del queroseno como única fuente de carbono y energía, donde el extracto de levadura que agrego como factor de crecimiento enriqueció el medio de cultivo mineral con vitaminas del complejo B, que activo las enzimas que se requieren para la oxidación de los hidrocarburos del queroseno, detectado por el aumento en la síntesis de proteína unicelular de S. exiguus como medida indirecta de la utilización del queroseno como fuente de carbono y energía. En tanto cuando S. exiguus creció en el queroseno al 5 %, pero ahora enriquecido con el extracto de malta, o bien con el LRM, como inductores of factores de crecimiento, se registró un peso de 0.8 g/L a las 24 h, lo que indica que la composición química del extracto de malta o el LRM no contenían los inductores de enzimas, necesarias para que S. exiguus pudiese utilizar los hidrocarburos de alifáticos del queroseno como única fuente de carbono y energía, en consecuencia hubo una evidente inhibición del crecimiento de S. exiguus, pues el peso seco detectado al final fue similar al registrado en el queroseno al 5 % usado como CA sin inocular con S. exiguus23,35,38. En la Tabla 1, se muestra la crecimiento de S. exiguus en queroseno al 5 %, al usarlo como única fuente de carbono y energía, en donde se demostró que requiere de NH4Cl al 1.2 %, para la síntesis de proteína y de los ácidos nucleicos, mientras que el extracto de levadura proporcionó las vitaminas del complejo B, para la activación de enzimas asociadas con la biodegradación hidrocarburos alifáticos como aromáticos del queroseno26,39,40. Los valores numéricos del crecimiento o rendimiento de S. exiguus en el queroseno cuando lo metabolizo como única fuente de carbono y energía, con el NH4Cl al 1.2 % plus el extracto de levadura como factor del crecimiento, fueron estadísticamente diferentes comparados con los valores del rendimiento o crecimiento celular de 0.73 g/L, que alcanzó la máxima velocidad de crecimiento con 0.049 h-1, en contraste con el prolongado de 14 h del tiempo de duplicación de S. exiguus en queroseno al 5 % como fuente de carbono y energía sin NH4Cl ni ningún factor de crecimiento, que se usó como CA. Lo anterior demostró la utilización del queroseno como único fuente de carbono y energía por S. exiguus con el NHCl4 como fuente de nitrógeno mineral, y que fue estimulado con el extracto de levadura, una fuente de vitaminas del complejo B, que facilitaron la reproducción de S. exiguus, medido indirectamente por la síntesis de proteína, o bien para el biotratamiento de ambientes impactados por queroseno conocida como bioaumentación una técnica de eliminación biológica dirigida de algún tipo de contaminante (s), mediante un microorganismo especifico como S. exiguus, que en ambientes impactados por queroseno podría eliminarlo33,34. Además, se observó el crecimiento de Aureobasidium sp. y Rhodotorula sp. en queroseno como única fuente de carbono y energía, pero sin la capacidad para consumirlo tan eficazmente como lo hace S. exiguus23-25.
La Figura 4 muestra el cromatograma que demuestra que cuando S. exiguus utilizo el queroseno al 5 % como fuente de carbono orgánico y energía se probó que tiene la capacidad de oxidar los hidrocarburos alifáticos de esta mezcla que contiene compuestos de entre 12 a 16 carbonos, basado en la desaparición de los picos de elución, de esos hidrocarburos33-35,38. Al respecto Okpokwasili & Amanchukwu39, reportaron que el género Candida posee los genes para utilizar mezclas de hidrocarburos similares al que existen en el queroseno40,41 por lo que estos géneros y especies de levaduras son una opción biológica adecuada para: i) la síntesis de proteína unicelular de alto valor nutricional y/o ii) la bioaumentación o eliminación “ex situ” de ambientes impactados por queroseno42. Mientras que Zinjarde & Pant43 reportaron la efectividad de las especies de Candida para biodegradarlo como: C. parapsilosis, C. guilliermondii, C. tropicalis al igual que Yarrowia lipolytica que mostró la mayor capacidad para eliminar la fracción alifática del queroseno hasta en un 78 %.
En la Tabla 2 se presentan las principales características morfológicas y bioquímicas de los géneros de las levaduras que usan los hidrocarburos del queroseno como fuente de carbono y energía, que se observaron en los cultivos jóvenes de levaduras de 24 h que tuvieron colonias brillantes convexas, con pigmentos olivácea. Al microscopio se observaron células que se dividen por gemación polar, en cultivos de 3-4 días las colonias se oscurecieron al formar un micelio en la periferia44. El género Rhodotorula se aisló del suelo impactado por queroseno de Altamira, Tam, al microscopio se observaron células ovales con gemación multilateral, sin micelio o pseudomicelio, en caldo no genero ninguna película, en medio de cultivo solido se observaron colonias cónicas, con un pigmento intracelular blanco de consistencia cremosa, de forma convexa. Rhodotorula no fermento la glucosa, aunque asimiló los NO3 -/45, mientras que el género Saccharomyces especie exiguus mostró al microscopio células ovales con gemación multilateral, de 1 a 4 ascosporas, en medio de cultivo líquido no generó ninguna película en la superficie, no asimiló los NO3 -, pero si fermentó la glucosa y la galactosa44,45.
Se concluye en esta investigación que S. exiguus es capaz de usar como fuente de carbono y energía hasta el 51 % de los hidrocarburos alifáticos del queroseno. Por lo anterior es posible reproducirla como una forma de proteína de origen unicelular (single cell protein) y eliminador de hidrocarburos del queroseno por bioaumentación para la recuperación de aguas y suelo impactados por derivados del petróleo similares o diferentes al queroseno.

