











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkJournal of the Selva Andina Research Society
versión On-line ISSN 2072-9294
J. Selva Andina Res. Soc. vol.10 no.2 La Paz 2019
Articulo Original
Respuesta de Zea mays a Burkholderia spp endófita de Zea mays var mexicana (teocintle)
Responds of Zea mays to Burkholderia spp endophyte from Zea mays var mexicana (teocinte)
García-Reyna Miriam Judith1, Santoyo-Pizano Gustavo1, Hernández-Mendoza José Luis2, Ignacio-De la Cruz Juan Luis1, Sánchez-Yáñez, Juan Manuel1*
1Universidad Michoacana de San Nicolás de Hidalgo. Francisco J. Mujica S/N, Col. Felicitas del Rio C.P. 58000, Morelia, Mich., México. Laboratorio de Microbiología Ambiental, Instituto de Investigaciones Químico Biológicas, Ed-B3 CU.
2Centro de Biotecnología Genómica (CBG), Boulevard del Maestro S/N esq. Elias Pina, Col. Narciso Mendoza, C.P. 88710, Apartado Postal No. 152, Cd. Reynosa, Tamaulipas, México. Teléfonos: 01 (899) 924 3627 o (55) 57 29 6000, Ext. 87726.
*Dirección de contacto: Juan Manuel Sánchez-Yáñez
Universidad Michoacana de San Nicolás de Hidalgo. Francisco J. Mujica S/N, Col. Felicitas del Rio C.P. 58000, Morelia, Mich., México. Laboratorio de Microbiología Ambiental, Instituto de Investigaciones Químico Biológicas, Ed-B3 CU.
E-mail address: syanez@umich.mx
Historial del artículo.
Recibido febrero 2019.
Devuelto mayo 2019
Aceptado junio, 2019.
Disponible en línea, agosto 2019.
Resumen
Zea mays var mexicana o teocintle es una gramínea, considerada como uno de los principales ancestros de Zea mays. El teocintle como otras gramíneas, se asocia con géneros de bacterias endófitas promotoras del crecimiento vegetal (BEPCV) en el interior de los tejidos vegetales. Z. mays es una gramínea doméstica cultivada normalmente bajo el sistema de producción intensiva, con relativa elevadas dosis de fertilizante nitrogenado (FENI), una posible alternativa de solución para minimizar este problema, es la inoculación de la semilla de Z. mays con géneros de BEPCV del teocintle, que colonizan las raíces, los tallos, las hojas y promueven un sano crecimiento a dosis reducida hasta en un 50% FENI como la urea, que equivale a disminuir de 280 kg/ha-1 a 140 kg/ha-1, de lo recomendado para la región del norte de Michoacán y sur de Guanajuato, México. Los objetivos de este trabajo fueron: i) determinar la densidad de Burkholderia spp BEPCV de teocintle, ii) analizar el efecto de la inoculación de Burkholderia spp de BEPCV en el crecimiento de Z. mays, a dosis reducida de FENI en invernadero, iii) identificar Burkholderia spp BEPCV de teocintle. Por principio se determinó la densidad de la población de Burkholderia spp BEPCV en tejidos (órganos), semillas, tallo-hojas y raíces del teocintle, mediante la siembra de estos, en agar Pseudomonas cepacia ácido azelaico triptamina (PCAT), posteriormente a la incubación, las colonias de Burkholderia spp se seleccionaron por la abundancia en: hojas, tallo, raíces y/o semillas, Estos aislados de Burkholderia spp, se codificaron con las claves T41 y T42 e inocularon en Z. mays sembrado en arena estéril, en donde como única variable-respuesta se determinó el peso seco total comparado el Z. mays cultivado con la dosis FENI recomendada, para esa región, se concluyó con la caracterizaron bioquímica. Los resultados muestran que en Burkholderia spp BEPCV de teocintle, se registró una densidad de 3.4 Log UFC/g de peso fresco (GPF) en la raíz, de 3.0 Log UFC/GPF en la semilla y un valor similar en el tallo-hojas de teocintle. La respuesta de Z. mays a Burkholderia T41 causó un incremento de un 47% en el peso seco de la raíz al amacollamiento, y de un 57% a la extensión del tallo, comparados con el Z. mays sin inocular con el nivel máximo de FENI (urea) 280 kg/ha-1. El perfil de identificación bioquímica de Burkholderia spp apoya una relación genética con B. vietnamiensis. Se concluye que el teocintle es una fuente de BEPCV para la producción sustentable Z. mays a dosis reducida de FENI.
Palabras clave: Teocintle, bacteria endófita, fijación biológica N2, fertilizante nitrogenado, Z. mays.
Abstract
Zea mays var mexicana or teocinte is a grass, considered one of the main ancestors of Zea mays. Teocinte, like other grasses, is associated with genera of endophytic plant growth promoters (EPGPB) in the interior of plant tissues. Z. mays is a domestic grass normally cultivated under the intensive production system, with relatively high doses of nitrogen fertilizer (NIFE), a possible alternative solution to minimize this problem, is the inoculation of the seed of Z. mays with genera of EPGPB teocinte, which colonize the roots, stems, leaves and promote a healthy growth at a reduced dose up to 50% NIFE such as urea, which is equivalent to decrease from 280 kg/ha-1to 140 kg/ha-1, of what is recommended for the región of northern Michoacán and southern Guanajuato, México. The objectives of this work were: i) to determine the density of Burkholderia spp BEPCV of teocinte, ii) to analyze the effect of the inoculation of Burkholderia spp EPGPB on the growth of Z. mays, at reduced dose of NIFE in greenhouse, iii) identify Burkholderia spp BEPCV teocinte. The population density of Burkholdería spp EPGPB was determined in tissues (organs), seeds, stem-leaves and roots of teocinte by sowing them on Pseudomonas cepacia azelaic acid tryptamine (PCAT), after incubation. Burkholdería spp colonies were selected for abundance in: leaves, stem, roots and/or seeds. These isolates of Burkholdería spp, were coded with the keys T41 and T42 and inoculated in Z. mays sown in sterile sand, where as the only variable-response the total dry weight was determined compared to the Z. mays cultivated with the recommended NIFE dose, for that región, it was concluded with the biochemical characterization. The results show that in Burkholdería spp BEPCV of teocinte, a density of 3.4 Log CFU/g of fresh weight (GPF) was registered in the root, of 3.0 Log CFU/GPF in the seed and a similar valué in the stem-leaves of teocinte The response of Z. mays to Burkholdería T41 caused an increase of 47% in the dry weight of the root to the tillering, and of 57% to the extensión of the stem, compared with the Z. mays without inoculating with the máximum level of NIFE (urea) 280 kg/ha-1. The biochemical identification profile of Burkholdería spp supports a genetic relationship with B. víetnamíensís. It is concluded that teocinte is a source of EPGPB for sustainable production Z. mays at a reduced dose of NIFE.
Key words: Teocinte, entophytic bacteria, biological fixing N2, N fertilizer, Z. mays.
Introducción
La gramínea Zea mays (maíz) es importante en México y el mundo por los múltiples usos en especial en la industria alimenticia, con un promedio de Z. mays solo a finales del 2000 de 22x106 ton/año1, el 50% se destina a consumo humano, de ahí la principal forma de este grano básico, es el sistema intensi vo, caracterizado por aplicar elevada dosis de ferti lizante nitrogenado (FENI) en función de la varieda d, tipo de suelo, así como de plaguicidas y agua.2-4
Una solución alternativa para racionar y optimizar el FENI en Z. mays, es la inoculación de semilla a la siembra con géneros y especies de bacterias promotoras del crecimiento vegetal (BPCV)4,5, para acelerar la germinación, colonizar las raíces e inducir un crecimiento sano, a través de la conversión de los exudados tanto de la semilla como del sistema de raíz, una amplia variedad de sustancias promotoras de crecimiento vegetal (SUPOCEVE) como el ácido-3-indol acético (AIA), que estimula la proliferación de pelos radicales, giberelinas y citocininas que mejoran la absorción radical para optimizar cualesquier dosis reducida de FENI.6 En consecuencia, la selección de géneros y especies de bacteria endófitas promotoras de crecimiento vegetal (BEPCV) que sean competitivas y efectivas en la reducción de la dosis de FENI para la producción sustentable del Z. mays, es posible a partir de gramíneas silvestres genéticas relacionadas con Z. mays como Zea mays var mexicana o teocintle.7-9 Las BEPCV inoculadas en semillas de Z. mays podrían minimizar el impacto negativo de los factores bióticos, que les impiden apoyar la producción sustentable de Z. mays, uno de los más limitantes es la competencia con la población microbiana nativa, el cual se suma a la depredación por protozoarios y/o la lisis por bacteriófagos que reducen la actividad de las BEPCV para ejercer el efecto positivo en la semilla y planta. Mi entras que factores abióticos, como los drásticos cambios de temperatura, la disponibilidad de agua, o carencia de suficientes de compuestos orgánicos e inorgánicos de N (nitrógeno), P (fosforo) y K (potasio), que evitan la supervivencia y/o crecimiento de géneros y especies de BEPCV antes de colonizar raíces de as diversas variedades de Z. mays cultivadas en el mundo. Un factor crítico limitante de la inoculación de Z. mays con BEPCV es el origen, en general si provienen de otra gramínea doméstica, no son competitivas en la colonización de la rizosfera de Z. mays.8,10,11 Una alternativa poco explorada, es el empleo de géneros y especies de BEPCV de teocintle, una gramínea con estrecha relación genética con el Z. mays11-14, que crece en suelos pobre en sales de N, P y K, de escasa humedad que causan stress por sequía, con capacidad de tolerar el ataque de fitopatógenos aéreos y radicales.15-17 Por lo anterior se supone una respuesta positiva de Z. mays a Burkholderia spp (BEPCV) de teocintle a dosis reducida del FENI recomendado para las diferentes regiones de producción, e incluso sin FENI.11,18-20
Las especies del género Burkholderia, endófitas tienen la ventaja de realizar la actividad benéfica desde el interior de los tejidos vegetales en comparación con las BPCV aisladas de plantas domésticas que ejercen la acción positiva desde el exterior del sistema radical.21,22 Con base a la diversidad de mecanismos específicos de las BEPCV en plantas, se supone que las aisladas de teocintle pueden ser una mejor opción a los inoculantes de BPCV para la reducción y optimización del FENI, en la producción sustentable de un mayor diversidad de variedades de Z. mays9,16,18,19 Lo anterior se apoya en la información reportada en literatura sobre 54 géneros y 129 especies de BPCV de valor comercial.16.23-29
En contrate investigación señalan otros géneros y especies de BEPCV como: Pseudomonas sp, Bacillus sp y Enterobacter sp.30,31 aisladas de Ananas comosus32,33 y de Burkholderia brasiliensis recuperada de Gossypium herbaceum. Mientras que la respuesta de Oryza sativa a B. vietnamiensis por SUPOCEVE a una dosis reducida de FENI34,35, causo un aumento 49% en el peso radical comparado con O. sativa sin inocular a la dosis de FENI recomendada para la región. En tanto que a respuesta de O. sativa a Burkholderia spp sin FENI, en cuyo caso en el sistema de conducción: floema y xilema16,34,36,37, por la fijación biológica del N2 incremento hasta en un 50% el N total. Otro ejemplo fue la respuesta positiva de Saccharum officinarum a B. brasiliensis que en mejoró en un 25% el rendimiento18,33 comparada con la respuesta S. officinarum cultivada con la dosis de FENI recomendada.
En general la investigación sobre la respuesta positiva de plantas domésticas a Burkholderia spp endófita es amplía22,23,27-29,38, pero mínima en gramíneas silvestres como el teocintle, como tampoco en relación a la respuesta favorable de Z. mays a Burkholderia endófita a dosis reducida de FENI hasta un 50% de urea, de acuerdo a lo recomendado en el norte de Michoacán y sur del bajío guanajuatense para el cultivo de Z. mays. Por lo anterior los objetivos de este trabajo fueron: i) aislar BEPCV de teocintle para Z. mays: ii) analizar la respuesta de Z. mays a Burkholderia spp con y sin FENI en invernadero y campo ii) Identificar las BEPCV de teocintle.
Materiales y métodos
Origen Zea mays var. mexicana (teocintle). El teocintle se colectó de los municipios Santa Ana Maya y Alvaro Obregón de donde es nativo con suelos franco-arcillosos, pobres en materia orgánica, en N, en PO43- soluble, de pH 6.8 ubicados en el noreste del estado de Michoacán a 35 km al noreste de More lia, Mich, México. En la figura 1 se describe el esquema de aislamiento, respuesta de Z. mays a los aislados de BEPCV de teocintle e la identificación bioquímica.
Determinación de la densidad de Burkholderia spp endófita de teocintle. Los aislados de Burkholderia spp se detectaron en semilla, en las raíces y en el tallo-hojas, en la etapa 6, de la escala de Feekes ó de la extensión del tallo39 y en la etapa 11.4 ó del llenado del grano, para ello los tejidos u órganos del teocintle se enjuagaron externamente con agua destilada y desinfectaron con un detergente comercial al 1.0% (p/v), en el caso de las raíces, se eliminaron los residuos del suelo, en tanto que cada semilla, tejido u órgano de teocinte se desinfectó con NaClO (hipoclorito de sodio) al 3% (p/v) por 15 min, se enjugaron 6 veces con agua estéril, luego se lavaron con tween 20 al 2% (v/v) por 5 min, posteriormente se enjuagaron 6 veces con agua estéril, finalmente se desinfectaron con etanol al 70% por 5 min y enjuagaron 3 veces en agua estéril, la efectividad de la desinfección de las semillas y tejidos del teocintle, se comprobó en agar nutritivo (Bioxon) e incubación a 28-30 °C/24 h.

Posteriormente de cada semilla o órgano de teocintle se pesó un gramo y trituró por separado en un mortero con NaCl al 0.85% (solución salina) estéril, se realizó una dilución desde 10-1 hasta 10-6 para conocer la densidad de la población de Burkholderia spp endófita, de la última dilución se tomó 0.1 mL para sembrar en caldo Watanabe mineral sin N, empleado para la prueba indirecta de fijación biológica de N2, con la siguiente composición química: solución A (g/L): ZnSO4 7H2O 0.43, MnSO4 H2O 1.30, NaMoO4 2H2O 0.75, H3BO3 2.8, CuSO4 5H2O 0.026, CoSO4 7H2O 0.07; solución B (g/L): MnSO4 7H2O 2.00, CaCl2 2H2O 2.00, FeSO4 7H2O 0.44, EDTA 0.04. Solución C (g/L): K2HPO4 60, KH2PO4 60, se mezclaron 50 mL de la solución B, 15 mL de la solución C 15 mL y 20 mL de la solución A por separado se agregaron (g/L): glucosa 5.0, sacarosa 4.5, manitol 3.5, almidón 4.0, el pH se ajustó a 6.8-7.0, el medio de cultivo esterilizó a 121 °C/15 min. Para el aislamiento de Burkholderia spp endófita se empleó el agar Pseudomonas cepacia ácido ázelaico triptamina (PCAT) (g/L): Mg SO4 0.1, ácido ázelaico 0.20, triptamina 0.2 g (disuelto en 2 mL de alcohol absoluto): K2HPO4 4, KH2PO4 4, extracto de levadura 0.02, el pH se ajustó a 5.7, las cajas de PCAT con la BEPCV se incubaron a 28 °C/72 h, en tanto que las de Watanabe se incubaron por 28 °C/48 h, los aislados endófitos de teocintle que crecieron, se resembraron en agar nutritivo para asegurar determinar la pureza.22


Identificación bioquímica de Burkholderia spp o BEPCV de raíces del teocintie. Los aislados de Burkholderia spp del teocintie se identificaron, mediante perfil bioquímico Tabla 1, como se describe en la literatura33, con el sistema API-50CH (BioMerieux, Francia), Burkholderia spp endófita del teocintie, se compararon con las cepas de referencia: B. cepacia ATCC 2546 y B. vietnamiensis TVV75, donadas por el Dr. J. Balandreu y el Dr. Tran Van Van, del Depto. de Ecología Microbiana de Suelos de la Universidad Claude Bernard, Lyon, Francia, además de estas pruebas bioquímicas en la Tabla 2 que se relacionan con este género, referente a la capacidad de utilizar diversas fuentes de carbono (C).
Medición indirecta de la capacidad de Burkholderia spp para la fijación biológica de N2. Los aislados de Burkholderia spp endófita de teocintie se suspendieron en NaCl 0.85% para inocular matraces erlenmeyer de 500 mL con 250 mL de caldo Watanabe, estos matraces se incubaron de 28-30 °C, después se tomaron alícuotas a intervalos de 24, 48, 72 y 96 h, para que por el método de Lowry cuantificar la cantidad de proteína en el medio de cultivo producto indirecto del crecimiento de Burkholderia spp al fijar N220
Inoculación de Z. mays con Burkholderia spp en invernadero. En este ensayo se usaron semillas de Z. mays "raza jala", donadas por el Laboratorio de Química de Productos Naturales perteneciente a la Facultad de Químico-Farmacobiología de la Universidad Michoacana de San Nicolás de Hidalgo.
Las semillas se pasteurizaron sumergidas en agua a 70 °C/5 min, para eliminar las bacterias endófitas que contienen según lo reporta en la literatura27-29, luego las semillas se colocaron en hielo por 5 min, se desinfectaron acorde con el procedimiento señalado en el inciso b), para aislar las bacterias endófitas de tejidos y órganos vegetales de teocintle.
Para el experimento del efecto de Burkholderia spp en Z. mays se sembraron 5 semillas por maceta de plástico con capacidad de 4 kg, en arena húmeda esterilizada a 121 °C/60min por 3 días, cuando las semillas germinaron se inocularon 3 mL de Burkholderia spp de teocintle a una densidad de 1x1O3 log UFC/semilla, después de germinar se dejaron 2 plantas de Z. mays por maceta, la superficie se cubrió con aserrín esterilizado a 121 °C/2 h, las macetas de Z. mays se mantuvieron en invernadero, se alimentaron 3 veces por semana con la solución mineral Thorton para plantas, esterilizada a 121 °C/15 min, cuya composición química fue (g/L): urea 10.0 (dosis reducida al 50%), K2HPO4 2.0, MgSO4 1.0, NaCl 0.1, FeSO4 0.1 y 10.0 mL solución de oligoelementos (g/L): H3BO3 2.86, ZnSO47H2O 0.22, MgCl2 7H2O 1.81, K2MnO40.09 y pH6.5-6.8.40
Diseño experimental y esquema de fertilización nitrogenada en Z. mays. En este ensayo Z. mays se inoculo con Burkholderia spp (aisladas como BEPCV de teocintle) con los siguientes tratamientos: control relativo (C) = Z. mays sin inocular con 280 Kg N/ha-1o la dosis 100% de FENI como urea recomendada para la región, control absoluto (0) = Z. mays sin inocular con Burkholderia spp y sin FENI; tratamiento B1 (0) = Z. mays con Burkholderia spp T41, sin FENI; tratamiento Bl (1/2) = Z mays inoculado con Burkholderia spp T41 y 50% de la dosis del FENI ó 140 kg/ha-1, tratamiento B2 (0) = Z. mays con burkholderia spp T42, sin FENI; tratamiento B2 (1/2) = Z. mays inoculado con Burkholderia spp T42 y 50% del FENI. El diseño se estableció con 20 repeticiones por tratamiento, la distribución de Z. mays en el invernadero fue un diseño de bloques al azar, se realizaron 2 muéstreos: el primero al amacollamiento según la escala Feekes39, el segundo a la extensión de tallo, en cada muestreo se separaron las raíces y el tallo para determinar su peso seco a 80 °C/24h.
Análisis estadístico. Los resultados se sometieron a la prueba de Tukey, con base en un diseño de 6 tratamientos y 20 repeticiones, para evaluar la respuesta de Z. mays a Burkholderia spp se determinó el peso seco, mientras que las diferencias en la respuesta de Z. mays por tratamiento se analizaron por comparación de medias, mediante la prueba de diferencia mínima significativa (DMS) a un nivel de significación de PO.05. Los cálculos se realizaron con el programa para computadora Statgrat versión 5.049
Resultados
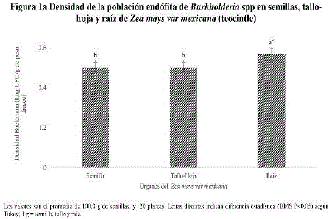
En la figura la, se muestra la densidad de la población endófita de Burkholderia spp o BEPCV de las raíces del teocintle, donde se detectó en un orden de magnitud de 3.4 log UFC/GPF (gramo de peso fresco), este valor numérico fue estadísticamente diferente comparado con los 3.0 log UFC/GPF de Burkholderia spp en el tallo-hoja y en semilla del teocintle.
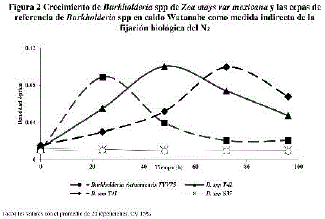
En la figura 2, se presenta el crecimiento de Burkholderia spp T41, T42 y S37 o BEPCV de teocintle que se aislaron en agar Watanabe sin ninguna forma de N combinado, en donde se registró que B. vietnamiensis TVV75 creció en 24 h, mientras que Burkholderia sp T41 lo hizo en 48 h y Burkholderia sp T42 en 72 h, comparado con las cepas de referencia: B. cepacia ATCC 25416, B. vietnamiensis TVV 75.
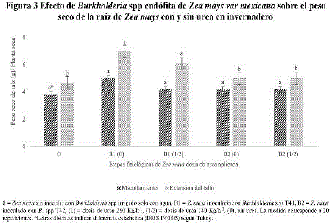
En la figura 3, se muestra la respuesta Z. mays Burkholderia spp T41 sin FENI, en donde se registró un aumento de un 47% en el peso seco radical (PSR) al amacollamiento, comparada con el PSR del Z. mays sin inocular con Burkholderia spp alimentada con el 50% del FENI (urea 140 kg/ha-1) y de Z. mays con el FENI al 100% (urea 280 kg/ha-1), referido como el control relativo. En tanto que Z. mays respondió positivamente a Burkholderia sp T42 sin FENI, ya que registro un incrementó del 11% el peso seco del tallo (PST), comparado con Z. mays alimentado con el FENI al 50% (140 kg/ha-1 de urea) que alcanzo 14% del PST, ambos valores numéricos fueron estadísticamente similares a Z. mays sin inocular y alimentado con el FENI al 100% (280 kg/ha-1de urea).
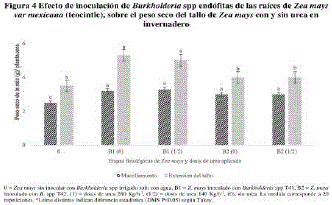
En la figura 4, se muestra la respuesta positiva de Z. mays a Burkholderia sp T41 (BEPCV del sin FENI, donde registró un 57% del PST, comparado con el Z. mays sin inocular alimentado con el FENI al 50% (140 kg/ha'de urea) que alcanzo un 39% del PST, valor numérico sin diferencia estadística al registrado en Z. mays, sin inocular con el FENI al 100% (280 kg/ha-1 de urea). En tanto que Z. mays tuvo un respuesta benéfica a Burkholderia sp T42 sin FENI con un aumento del 18% del PST, valor numérico sin diferencia estadística comparado con el PST de Z. mays con el FENI al 50% (140 kg/ha de urea), sin diferencia estadística con el PST del Z. mays no inoculado con las BEPCV, alimentado con el FENI al 100% (280 kg/ha-1de urea).
En la tabla 1, se muestra el perfil bioquímico de los aislados de BEPCV del teocintle, basado en la literatura que reporta que Burkholderia spp este género se describe como un bacilo Gram negativo, móvil, catalasa y oxidasa positivo, no produce ácido sulfhídrico (H2S), ni triptofanasa, que usa la sintetiza citocromo oxidasa, que tiene un patrón oxidativo en la utilización de azúcares, como fuentes de C y energía, pero no libera ácido de glucosa, pero sí como el citrato de sodio, algunas especies tienen la capacidad genética de fijar N2, por lo que crecen medio de cultivo mineral sin ninguna forma de N combinado inorgánico u orgánico. En la Tabla 1 también se presentan las diferencias bioquímicas básicas entre Burkholderia spp de teocintle y el género Pseudomonas spp22,41, con patrones bioquímicos que apoyan la estrecha relación genética entre ambos géneros, y las diferencias que señalan como 2 géneros distintos, en consecuencia los aislados BEPCV de teocintle fueron clasificados como miembros de especies de Burkholderia.22,34,38
La Tabla 2, se muestra el patrón de asimilación de 50 carbohidratos de los BEPOCV de teocintle denominadas: T421 y T42 en comparación con las cepas de referencia B. cepacia ATCC 25416 y B. vietnamiensis TVV 75. El perfil de asimilación de los aislados de BEPCV fue: acetato, D-![]() -alanina, succinato, 2-ceto-gluconato, celobiosa, D-arabinosa, D-arabitol, D-fructuosa, D-glucosa, D-lisosa, D-manosa, D-tagatosa, D-dulcitol, D-xilosa, esculina, galactosa, glicerol, gluconato, inositol, D-arabinosa, D-fucosa, manitol, N-acetil glucosamina, ribosa, sacarosa, sorbitol, pero no
-alanina, succinato, 2-ceto-gluconato, celobiosa, D-arabinosa, D-arabitol, D-fructuosa, D-glucosa, D-lisosa, D-manosa, D-tagatosa, D-dulcitol, D-xilosa, esculina, galactosa, glicerol, gluconato, inositol, D-arabinosa, D-fucosa, manitol, N-acetil glucosamina, ribosa, sacarosa, sorbitol, pero no ![]() -metil D-glucósido,
-metil D-glucósido, ![]() -metil D-manosida, almidón, (
-metil D-manosida, almidón, (![]() -metil xilosa, ramnosa, melicitosa, melibiosa, maltosa, lactosa, erytritol, L-sorbosa. En tanto que tanto B. vietnamiensis TVV75 como Burkholderia sp T42, usaron (3-gentobiosasola y D-fucosa, mientras que B. cepacia y Burkholderia sp T41 y T42 utilizaron, arabutina, en contraste con B. cepacia que uso D-arabitol: mientras que B. vietnamiensis fue capaz de usar furanosa, contrario Burkholderia sp T41 que utilizaron salicilina, diferente a Burkholderia sp T41 y T42 aislados que usaron glucógeno e inulina como fuente de C y energía. Las diferencias en el patrón bioquímico de utilización o asimilación de azucares y ácidos grasos podrían emplearse en el diseño de medios de cultivo selectivos y diferenciales para cada una de las especies de Burkholderia tanto endófitos de plantas como los de los ambientes en la naturaleza, pues este género es cosmopolita. Las BEPCV relacionados con Burkholderia spp de teocintle T41 y T42, mostraron un patrón de asimilación de azúcares y sus derivados, similar al reportado para otras especies de Burkholderia22, en especial B. vietnamiensis.34
-metil xilosa, ramnosa, melicitosa, melibiosa, maltosa, lactosa, erytritol, L-sorbosa. En tanto que tanto B. vietnamiensis TVV75 como Burkholderia sp T42, usaron (3-gentobiosasola y D-fucosa, mientras que B. cepacia y Burkholderia sp T41 y T42 utilizaron, arabutina, en contraste con B. cepacia que uso D-arabitol: mientras que B. vietnamiensis fue capaz de usar furanosa, contrario Burkholderia sp T41 que utilizaron salicilina, diferente a Burkholderia sp T41 y T42 aislados que usaron glucógeno e inulina como fuente de C y energía. Las diferencias en el patrón bioquímico de utilización o asimilación de azucares y ácidos grasos podrían emplearse en el diseño de medios de cultivo selectivos y diferenciales para cada una de las especies de Burkholderia tanto endófitos de plantas como los de los ambientes en la naturaleza, pues este género es cosmopolita. Las BEPCV relacionados con Burkholderia spp de teocintle T41 y T42, mostraron un patrón de asimilación de azúcares y sus derivados, similar al reportado para otras especies de Burkholderia22, en especial B. vietnamiensis.34
Discusión
En la figura 1 a, se muestra que Burkholderia sp es una endófita del teocintle, distribuida a través del sistema de conducción en todos órganos que incluye la semilla, en especial en las raíces, como se reporta para otros géneros y especies de endófitos en una amplia variedad de vegetales domésticos y silvestres diferentes al teocintle 30,31, lo que confirma los señalado en la literatura sobre Burkholderia spp distribuida en plantas de cualquier ambiente.9,16,18,20,26
El crecimiento B. vietnamiensis TVV 75 y de Burkholderia spp T41 y T42 se presenta en figura 2 en caldo Watanabe, lo que demuestra que son capaces de fijar N2 9,11,42 Mientras que la cepa de referencia B. cepacia ATCC 25416 al igual que Burkholderia sp S37 aunque es una BEPCV de teocintle no fijan N2 en concordancia con la literatura que reporta que especies relacionadas con B. cepacia pueden sintetizar SUPOCEVE aunque no fijen N222, a diferencia B. vietnamiensis TVV75 una endófita que fija N2 libremente como en asociación con O. sativa.9,34,35,43 Los aislados de Burkholderia sp T41 y T42 revelaron un perfil bioquímico diferente B. vietnamiensis, lo que apoya se trata de otra especie capaz de fijar N2.
En la figura 3 se muestra la respuesta positiva de Z. mays a Burkholderia spp a la dosis 50 % de FENI (140 kg/ha-1 de urea), fue debido a que cuando colonizó la raíz de Z. mays, trasformó los exudados radicales en SUPOCEVE, lo que indujo una mayor proliferación de los pelos radiculares que mejoraron y optimizaron la absorción radical del FENI a pesar de reducir la dosis hasta un 50%.11,16 Mientras que la respuesta de Z. mays a Burkholderia sin FENI, se explica con base a que tiene la capacidad de fijar N2, actividad dependiente de la concentración del FENI disponible5,16, la fijación de N2 una propiedad genética que se ha reportado en B. vietnamiensis, pero los endófitos son relativamente menos conocidos que los géneros de BPCV fijadoras de N2 de la rizosfera9,11,28 de gramíneas y que estimulan el crecimiento vegetal a dosis reguladas del FENI.19,36,43,44
En la figura 4 se muestra la respuesta de Z. mays a Burkholderia spp sin FENI, y que apoya que este género fijar N2 en interior de las raíces de Z. mays9 de forma análoga a lo reportado en O. sativa inoculado con B. vietnamiensis sin FENI.16,28 La respuesta positiva de Z. mays a Burkholderia spp fue similar a lo reportado en la literatua20 con S. officinarum inoculado con Azotobacter diazotrophicus que aumento en un 50% el contenido de N, sin FENI mediante fijación de N2 y la síntesis de SUPOCEVE.42,43,45,46 Respuesta similar a la S. officinarum sin inocular alimentada con el FENI al 100%. La respuesta favorable de Z. mays a Burkholderia spp, indica que es probablemente por la capacidad de sintetizar la nitrogenasa para fijar el N2, cuya actividad que no se inhibir por la concentración del FENI en el suelo o por el aplicado, contrario a lo señalado en la literatura sobre S. officinarum y A. diazotrophicus en que a dosis moderadas de FENI, se inhibió la actividad de fijación biológica de N2, por un efecto feedback, dependiente de la concentración del N mineral en el suelo.29,47,48
En la relación a la identificación de los aislados de BEPCV de teocintle el género y especies de Burkholderia fueron las principalmente reconocidas, el perfil bioquímico sugiere que algunos están asociados genéticamente con B. vietnamiensis, primero por la capacidad de fijar N2 y porque también sintetizan SUPOCEVE algunas de las modifican el patrón de crecimiento del sistema radical de Z. mays, por compuestos volátiles diferentes a las fithormonas conocidas, y que actualmente reporta que son parte de los mecanismos de promoción para mejorar el crecimiento vegetal a dosis reguladas de FENI.43,45,46 Mientras que otra explicación al efecto positivo de Burkholderia spp (BEPCV), en Z. mays fue dependiente de la conversión de compuestos orgánicos, derivados de metabolismo vegetal durante la fotosíntesis en SUPOCEVE, este clase de fithormonas, tiene que ver con el género y especies de BPCV o BEPCV, así como de la especie vegetal específica con la que se asocian, tanto a nivel endófita o radical y que es regulado por la concentración de FENI o N disponible en el suelo.18,23,44,45,48
Con base en lo anterior se concluye que en los órganos o tejidos del teocintle: raíces, tallo-hojas y semillas en ese orden de densidad existe Burkholderia spp. Mientras que estos aislados de BEPCV fueron benéficas para el crecimiento de Z. mays, en función de la concentración del FENI aplicado, aunque también se detectó que algunos aislados de Burkholderia fijan N2 en ausencia de FENI sin N disponible.
El perfil bioquímico de los aislados de BEPCV de teocintle, apoya que algunas de estas especies están relacionadas con B. vietnamiensis por la capacidad de fijar N2, aunque también porque sintetizan SUPOCEVE, ambos mecanismos hacen posible que este grupo de BEPCV sean consideradas útiles en la producción sustentable de Z. mays.
Conflictos de intereses
Los participantes en esta investigación declaramos que no existe ningún problema en la relación con la información aquí divulgada con las instituciones participantes así como quienes la financiaron total o parcialmente.
Agradecimientos
Al Proyecto 2.7 (2019) de la CIC-UMSNH, a BIONUTRA S A de CV Maravatio, Mich, México, a Beatriz Noriega-Gamboa por la escritura.
Aspectos éticos
La aprobación de la investigación por el Comité de Ética, de la Universidad Michoacana de San Nicolás de Hidalgo - México, siguió las pautas establecidas para publicaciones en la divulgación de la ciencia generada.
Literatura citada
1. Eubanks MW. The mysterious origin of maize. Econ Bot 2001;55(4):492-514. [ Links ]
2. Ferrera Cerrato R. Efecto de rizosfera. En: Ferrera Cerrato R, Pérez Moreno J, editores. Agromicrobiología, elemento útil en la agricultura sustentable. Montecillos, Estado de México: Colegio de postgraduados en Ciencias Agrícolas, 1995. p. 36-50. [ Links ]
3. Cárdenas Navarro R, Sánchez Yáñez JM, Farías Rodríguez R, Peña Cabriales JJ. Los aportes de nitrógeno en la agricultura. Rev Chapingo Ser Hortic 2004; 10(2): 173-8. http://dx.doi.org/10.51_54/r.rchsh.2002.07.039 [ Links ]
4. Armenta Bojórquez AD, García Gutiérrez C, Camacho Báez JR Apodaca Sánchez MA, Gerardo Montaya L, Nava Parez E. Biofertilizantes en el desarrollo agrícola de México. Ra Ximbai 2010;6(1):51-6. DOI: http://www.dx.doi.org/10.35197/rx.06.01.2010.07.aa [ Links ]
5. Dobbelaere S, Croonenborghs A, Thys A, Ptacek D, Vanderleyden J, Dutto P, et al. Responses of agronomically important crops to inoculation with Azospirillum. Aust J Plant Physiol 2001; 28(9):871-9. DOI: https://dx.doi.org/10.1071/PP01074 [ Links ]
6. Askary M. Mostajeran A, Amooaghaei R, Mosta jeran M. Influence of the co-inoculation Azospiri llum barasilense and Rhizobium melilotus plus 2,4-D on grain yield and N,P,K content of Triticum aestivum (cv Baccros and Mahdavi) Am Eurasian J Agrie Environ Sci 2009;5(3):296-307. [ Links ]
7. Bashan Y, Levanony H. Current status of Azospirillum inoculation technology: Azospirillum as achallenge for agriculture. Can J Microbiol 1990; 36(9):591-608. DOI: https://dx.doi.org/10.1139/m90-105 [ Links ]
8. Bashan Y, Holguin G, Ferrera Cerrato R. Interacciones entre plantas y microorganismos benéficos. Terra 1996; 14(2): 159-92. [ Links ]
9. Franche C, Lindstrom K, Elmerich C. Nitrogen-fixing bacteria associated with leguminous and non-leguminous plants. Plant Soil 2009;321(1-2):35-59. DOI: https://dx.doi.org/10.1007/s11104-008-9833-8 [ Links ]
10. Dos Reis Júnior FB, Massena Reis V, Urquiaga S, Dóbereiner J. Influence of nitrogen fertilisation on the population of diazotrophic bacteria Herbaspirillum spp. and Acetobacter diazotrophicus in sugar cane (Saccharum spp.). Plant Soil 2000;219(1-3): 153-9. https://dx.doi.org/10.1023/A:1004732500983 [ Links ]
11. Avis TJ, Gravel V, Antoun H, Tweddell RJ. Multifaceted beneficial effects of rhizosphere microorganisms on plant health and productivity. Soil Biol Biochemy 2008;40(7): 1733-40. DOI: https://dx.doi.org/10.1016/j.soilbio.2008.02.013 [ Links ]
12. Wilkes HG. Maiz and its wild relatives. Science 1972; 177(4054):1071-7. [ Links ]
13. Wilkes HG. Hybridization of maize and teosinte in México and Guatemala and improvement of maize. Econ Bot 1997;31:254-93. DOI: http://dx.doi.org/10.1007/BF02866877 [ Links ]
14. Galinat WC. A reconstruction of a possible role of crucial observations leading to a rapid domestic transformation of wild teosinte into the first maize. Econ Bot 2001;55(4):571-4. [ Links ]
15. Wilkes HG. Teosinte: the closest relative of maize revised. Maydica 1985;30:209-23. [ Links ]
16. Ahemad M, Kibret M. Mechanisms and applications of plant growth promoting rhizobacteria: Current perspective. J King Saud Univ Sci 2014;26(1):1-20. DOI: https://dx.doi.org/10._1016/j.jksus.2013.05.001 [ Links ]
17. Sánchez G JJ, Ordaz SL. El teocintle en México. Distribución y situación actual de las poblaciones. Systematic and ecogeographic studies on crop genepools: 2. IBPGR, Rome; 1987. [ Links ]
18. Nejad P, Johnson PA. Endophytic bacteria induce growth promotion and wilt disease suppression in oilseed rape andtomato. Biol Control 2000; 18(3) :208-15. DOI: https://dx.doi.org/10.1006/bcon.2000.0837 [ Links ]
19. Sturz AV, Nowak J. Endophytic communities of rhizobacteria and the strategies required to create yield enhancing associations with crops. Appl Soil Ecol 2000;15(2): 183-90. DOI: https://dx.doi.org/10.1016/S0929-1393(00)00094-9 [ Links ]
20. Sevilla M, De Oliveira A, de Baldani I, Kennedy C. Contributions of the endophyte Acetobacter diazotrophicus to sugarcane nutrition: a preliminary study. Symbiosis 1998;25:181-91. [ Links ]
21. Magalhães Cruz L, Maltempi de Souza E, Baler Weber O, Ivo Baldani J, Döbereiner J, de Oliveira Pedrosa F. 16S Ribosomal DNA characterization of nitrogen-fixing bacteria isolated from banana (Musa spp.) and pineapple (Ananas comosus (L.) Merril). Appl Environ Microbiol 2001;67(5):2375-9. DOI: http://dx.doi.org/10._1128/AEM.67.5.2375-2379.2001 [ Links ]
22. Castellanos Morales V, Villegas J, Cárdenas Navarro R, Farías Rodríguez R, Sánchez Yáñez JM. Diversity of Burkholderia cepacia associated to teocinte. Phyton 2005;54:27-38. [ Links ]
23. Gardner JM, Feldman AW, Zablotowicz RM. Identity and behavior of xylem-residing bacteria in rough lemon roots of Florida citrus trees. Appl Environ Microbiol 1982;43(6): 1335-42. [ Links ]
24. McInroy JA, Kloepper JW. Survey of indigenous bacterial endophytes from cortón and sweet corn. Plant Soil 1995a; 173(2):337-42. DOI: https://dx.doi.org/10.1007/BF00011472 [ Links ]
25. McInroy JA, Kloepper JW. Population dynamics of endophytic bacteria in field-grown sweet corn and cotton. Can J Microbiol 1995b;41(10):895-901. DOI: https://dx.doi.org/10.1139/m95-123 [ Links ]
26. Sturz AV. The role of endophytic bacteria during seed piece decay and potato tuberization. Plant Soil 1995;75(2):257-63. DOI: https://doi.org/10.1007/BF00011362 [ Links ]
27. Halmann J, Kloepper J.W, Rodríguez Kábana R, Sikora RA. Endophytic rizobacteria as antagonists of Meloidogyne incógnita on cucumber. Phytopatology 1997a;85:1136. [ Links ]
28. Hallmann J, Quadt Hallmann A, Mahaffee WF, Kloepper JW. Bacterial endophytes in agricultural crops. Can J Microbiol 1997b;43(10): 895-914. DOI: https://dx.doi.org/10.1139/m97-131 [ Links ]
29. Hallmann J, Kloepper JW, Rodríguez Kábana R. (1997c). Application of the Scholander pressure bomb to studies on endophytic bacteria of plants. Can J Microbiol 1997c;43(5):411-6. DOI: https://dx.doi.org/10.1139/m97-058 [ Links ]
30. Sharrock KR, Parkes SL, Jack HK, Rees George J, Hawthorne BT. Involvement of bacterial endophytes in storage rots of buttercup squash (Cucúrbita máxima D. hybrid 'Delica'). N Z J Crop Hortic Sci 1991;19(2): 157-65. DOI: https://dx.doi.org/10.1080/01140671.1991.10421794 [ Links ]
31. Benhamou N, Kloepper JW, Quadt Hallman A, Tuzun S. Induction of defense-related ultrastructural modifications in pea root tissues inoculated with endophytic bacteria. Plant Physiol 1996; 112 (3):919-29. DOI: http://dx.doi.org/10.1104/pp.112.3.919 [ Links ]
32. Weber OB, Baldani VLD, Teixeira KRS, Kirchhof G, Baldani JI, Dobereiner J. Isolation and characterization of diazotrophic bacteria from banana and pineapple plants. Plant Soil 1999;210(1): 103-13. DOI: https://dx.doi.org/10.1023/A:1004623523179 [ Links ]
33. Mattos KA, Jones C, Heise N, Previato JO, Mendonça Previato L. Structure of an acidic exopoly-saccharide produced by the diazotrophic endophytic bacterium Burkholderia brasiliensis. Eur J biochem2001;268(11):3174-9. DOI: http://dx.doi.org/10.1046/j.1432-1327.2001.02196.x [ Links ]
34. Gillis M, Van Van T, Bardin R, Goor M, Hebbar P, Willems A, Fernandez MP. Polyphasic taxonomy in the genus Burkholderia leading to an emended description of the genus and proposition of Burkholderia vietnamiensis sp. nov. for N2-fixing isolates from rice in Vietnam. Int J Syst Evol Microbiol 1995;45(2):274-89. DOI: http://dx.doi.org/10.1099/00207713-45-2-274 [ Links ]
35. TrânVan V, Berge O, Ngô Kê S, Balandreau J, Heulin T. Repeated beneficial effects of rice inoculation with a strain of Burkholderia vietnamiensis on early and late yield components in low fertility sulphate acid soils of Vietnam. Plant Soil 2000;218(1-2):273-84. DOI: https://dx.doi.org/10.1023/A:1014986916913 [ Links ]
36. Minerdi D, Fani R, Gallo R, Boarino A, Bonfante P. Nitrogen fixation genes in an endosymbiotic Burkholderia strain. Appl Environ Microbiol 2001;67(2):725-32. DOI: http://dx.doi.org/10.1128/AEM.67.2.725-732.2001 [ Links ]
37. Mundt JO, Hinkle NF. Bacteria within ovules and seeds. Appl Environ Microbiol 1976;32:694-8. [ Links ]
38. Baldani JI, Caruso L, Baldani VLD, Goi SR, Dobereiner J. Recent advances in BNF with non-legume plants. Soil Biol Biochem 1997;29(5-6): 911-92. https://dx.doi.org/10.1016/S0038-0717(96)00218-0 [ Links ]
39. Large EC. Growth stages in cereals illustration of the Feekes scale. Plant Pathol 1954;3(4): 128-9. DOI: https://doi.org/10.1111/j.1365-3059.1954.tb00716.x [ Links ]
40. García-González MM, Peña C JJ, Moreno MP, Sánchez Yáñez JM. Inoculación de maíz con bacterias fijadoras de N2. Terra latinoam 1995;13:71-80. [ Links ]
41. Valdivia-Urdiales B, Sánchez-Yánez JM, Pena Cervantes E, Fernández Brondo JM. Efecto de la inoculación con Glomus spp. y Pseudomonas putida en Trigo. Rev Latinoam Microbiol 1999;41 (4): 231-38. [ Links ]
42. Ortiz Castro R, Valencia Cantero E, López Bucio J. The beneficial role of rhizosphere microorganisms in plant health and productivity: improving root development and nutrient acquisition. Acta Hort 2013;1009:241-50. DOI: http://dx.doi.org/10.17660/ActaHortic.2013.1009.29 [ Links ]
43. Hernández A, Rives N, Caballero A, Hernández AN, Heydrich M. Caracterización de rizobacterias asociadas al cultivo de maíz en la producción de metabolitos del tipo AIA, sideróforos y ácido salicílico. Re Colomb Biotecnol 2004;6(1):6-13. [ Links ]
44. Bottini R, Cassán F, Piccoli P. Gibberellin production by bacteria and its involvement in plant growth promotion and yield increase. Appl Microbiol Biotecnol 2004;65(5):497-503. DOI: http://dx.doi.org/10.1007/s00253-004-1696-1 [ Links ]
45. Gutiérrez Luna FM, López Bucio J, Altamirano Hernández J, Valencia Cantero E, Reyes de la Cruz HR, Macías Rodríguez L. Plant growth-promoting rhizobacteria modulate root-system architecture in Arabidopsis thaliana through volatile organic compound emission. Symbiosis 2010;51(1):75-83. DOI: https://dx.doi.org/10.1007/s13199-010-0066-2 [ Links ]
46. Orozco Mosqueda MC, Velázquez Becerra C, Macías Rodríguez LI, Santoyo G, Flores Cortez I, Alfaro Cuevas R, et al. Arthrobacter agilis UMCV2 induces acquisition in Mendicago truncatula (Strategy I plant) in vitro vía dimethylhex-adecylamine emission. Plant Soil 2013;362(1-2): 51-66. DOI: https://doi.org/10.1007/s11104-012-1263-y [ Links ]
47. Sturz AV, Christie BR, Nowak J. Bacterial endophytes: potential role in developing sustainable systems of crop production. Crit Rev Plant Sci 2000;19(1):1-30. DOI: https://dx.doi.org/10.1080/07352680091139169 [ Links ]
48. Berendsen RL, Pieterse CM, Bakker PA. The rhizosphere microbiome and plant health. Trends Plant Sci 2012;17(8):487-6. DOI: http://dx.doi.org/10.1016/j.tplants.2012.04.001 [ Links ]
49. Walpole, E R, Meyers R and Meyers LS. Probabilidad y Estadística para Ingeniería y Ciencias. Ed. Pearson 8ed. México. [ Links ]

