









 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO  uBio
uBio 
 Permalink
PermalinkJournal of the Selva Andina Research Society
versão On-line ISSN 2072-9294
J. Selva Andina Res. Soc. vol.6 no.2 La Paz 2015
Artículo Original
Correlación fenotípica y genotípica-molecular de la resistencia a enfermedades en variedades mejoradas de papa (Solanum tuberosum L.)
Phenotypic and molecular-genotypic correlation of diseases resistance in improved varieties potato (Solanum tuberosum L.)
Baldelomar Mariela1, Gabriel Julio2* Veramendi Silene2, Terán Ariana1, Plata Giovanna2
1Biología, Universidad Mayor de San Simón, Final Sucre frente al parque La Torre, Cochabamba, Bolivia, Telf. (591) 4-4256747.
2Fundación PROINPA, Casilla 4285, Av. Meneces s/n Km 4 (zona El Paso), Cochabamba, Bolivia. Telf. (591) 4-4319595.
*Dirección de contacto: Fundación-PROINPA
Fundación para la Promoción e Investigación de Productos Andinos.
P.O. Box 4285 Cochabamba, Bolivia. Tel: +591-4-4319595, Fax: +591-4-4319500.
Julio Gabriel
E-mail address : j.gabriel@proinpa.org
Historial del artículo.
Recibido febrero, 2015.
Devuelto mayo 2015
Aceptado julio, 2015.
Disponible en línea, agosto, 2015.
Resumen
En dos campañas consecutivas 2010-2011 y 2011-2012 en invernaderos de la Fundación PROINPA fueron sembrados 18 variedades mejoradas de papa y dos variedad testigo (Waycha y Desirée) con el objetivo de estudiar: i) La resistencia a tizón (Phytophthora infestans), verruga (Synchytrium endobioticum) y virus PVX y ii) Validar los marcadores moleculares GP94, RB, CP60 y NL25 para co-localizar con los genes mayores de resistencia de las enfermedades mencionadas. Los resultados mostraron diferencias notables entre variedades para AUDPCrel (tizón), severidad (verruga) y severidad y absorvancia (PVX). Se observó diferentes niveles de resistencia en las variedades, encontrándose variedades con alta resistencia y moderada resistencia. Los análisis moleculares fueron altamente notables al Pr<0.01 de probabilidad para tres de los marcadores utilizados. Se observó que el alelo para el marcador GP94 se co-localizó con el gen de resistencia Rpi-phu1 a tizón a una distancia de 350 pb, observándose el alelo de resistencia en todas las variedades con excepción de la variedad Waycha. El alelo para el marcador RB que co-localiza con el gen RB para resistencia a tizón, no fue hallado en ninguna de las variedades evaluadas. El alelo para el marcador CP60 se co-localizó con el gen Rx1 para resistencia a PVX a una distancia de 350 pb y se observó en casi todas las variedades, con excepción de la variedad Chota Ñawi. Finalmente, el alelo para el marcador NL25 se co-localizó con el gen Sen1 para resistencia a verruga a una distancia de 1000 pb y fue observado en todas las variedades.
Palabras clave:
Tizón tardío, verruga de la papa, virus PVX, severidad, absorvancia, inoculo.
Abstract
In two consecutive agricultural campaigns (2010-2011 and 2011-2012), 18 improved potato varieties were planted in the PROINPA Foundation greenhouses as well as two witness varieties (Waych’a and Desirée) with the aim to study: i) late blight resistance (Phytophthora infestans), wart (Synchytrium endobioticum) an virus PVX and ii) validate molecular markers GP94, RB, CP60 and NL25 for co-localizing with major resistance genes of diseases mentioned. The results showed significant differences among varieties for AUDPCrel (late blight), severity (Wart) and severity and absorbance (PVX). Different levels of resistance were observed in varieties, such varieties with high resistance and moderate resistance. Molecular analysis were highly significant to Pr<0.01 of probability for three of the market used. It was observed that the allele for marker GP94 was co-located with the resistance gene Rpi-phu1 for late blight at a distance of 350 bp, showing the resistance allele in all varieties except the Waych'a variety. The allele for RB marker that is co-localizes with the RB gene for resistance to blight was not found in any of the varieties tested. The allele for marker CP60 was co-located with Rx1 gene to PVX resistance at a distance of 350 bp and it was observed in almost all varieties except Chota Ñawi variety. Finally, the allele for marker NL25 is co-located with Sen1 gene resistance to wart at a distance of 1000 bp and it was observed in all varieties.
Key words:
Late blight, potato wart, PVX virus, severity, absorbance, inoculum.
Introducción
Bolivia forma parte del centro de origen, domesticación y diversificación de la papa (Solanum tuberosum L.). La biodiversidad de la papa en el país abarca a más de 1500 variedades nativas, 39 especies silvestres y ocho especies cultivadas (Zeballos et al. 2009, Gabriel 2010). El cultivo de papa tiene importancia económica, social y política para el país (Zeballos et al. 2009). Se cultiva en los altiplanos norte, medio sud, y los valles mesotérmicos del país.
Entre las enfermedades y plagas de la papa se destacan 25 virus, 38 hongos, 6 bacterias, 2 micoplasmas, 1 viroide, 68 nemátodos y 128 insectos, estimándose las pérdidas de rendimiento por enfermedades por año cerca al 22% (Estrada 2000).
De estos patógenos, tizón tardío, oomycete Phytophthora infestans (Mont.) De Bary, es la enfermedad de importancia de la papa en el mundo y en Bolivia. En nuestro país, se estima que más de 20000 ha de papa son atacadas por tizón tardío, afectando a 40000 familias de agricultores, ocasionando una pérdida directa de 30 millones de dólares anuales (Navia et al. 2009).
Se identificó 11 genes de resistencia vertical que provienen de especies silvestres, Solanum demissum, que inducen respuesta hipersensible a la infección con razas específicas de P. infestans, también se identificó QTLs de resistencia en los doce cromosomas de la papa que confieren un tipo de resistencia horizontal (Mosquera et al. 2008). La especie silvestre diploide Solanum bulbocastanum, revelo una resistencia contra todas las razas conocidas de tizón tardío gracias al gen RB localizado en el cromosoma VIII (Colton et al. 2006). En la especie Solanum phureja, se identificó el gen Rpi-phu1, que confiere una amplia resistencia al tizón tardío y se identificó en el cromosoma IX, próximo al marcador GP94 (Sliwka et al. 2010).
Por otra parte la verruga causada por el hongo Synchytrium endobioticum (Schilb.) Perc., ocasiona pérdidas totales del cultivo de papa. También perjudica a futuros cultivos de papa, ya que sus esporas de descanso sobreviven en el suelo más de 30 años (Torres 2002). Existen variedades de papa con resistencia vertical contra este patógeno. El gen dominante Sen1, localizado en el cromosoma XI de la papa, confiere resistencia al patotipo1 (D1) de S. endobioticum (Mosquera et al. 2008). Este gen se co-localiza con el marcador molecular NL25, a una longitud de 700 pb (Bormann 2004).
Entre las enfermedades virales que atacan al cultivo de papa, la ocasionada por PVX o virus del mosaico puede reducir el rendimiento en más del 20%, y en combinación con otros virus puede provocar severas pérdidas (Alvarez et al. 2009). El mejoramiento para resistencia al virus PVX es de tres tipos: resistencia a la infección, hipersensibilidad local y resistencia extrema. La resistencia a la infección sólo se mantiene en la progenie cuando ambos padres tienen alta resistencia. El mejoramiento se basa en las reacciones de hipersensibilidad (Nb, Nx) y resistencia extrema (Rx: Rx1 en el cromosoma XII y Rx2 en el cromosoma V). La resistencia extrema o inmunidad está determinada por el gen dominante Rx, este gen confiere resistencia a todas las razas (strains) del virus PVX (Estrada 2000). Las especies cultivadas de papa con resistencia al virus PVX son: Solanum tuberosum, S.x ajanhuiri, S.x juzepczukii y S.x curtilobum. La especie cultivada S. phureja presenta resistencia de campo o tolerancia al ataque del virus (Ochoa 2001). Las especies silvestres S. acaule y S. sucrense fueron reportadas como resistentes por Nayudu (2008). En el cromosoma XII de la papa se halla el gen Rx1, que da resistencia al virus PVX, que se co-localiza con el marcador molecular CP60 (Bendahmane et al. 1997).
La forma de control de estos patógenos es a través de productos químicos, aislamiento de cultivos, medidas cuarentenarias u otros, siendo métodos costosos. Por lo que el desarrollo de variedades resistentes y menos costosa, se puede lograr un control óptimo, duradero, y así se podría contribuir a una mayor producción en calidad y en cantidad (Rojas et al. 2007).
Para ello, el programa de mejoramiento genético en la Fundación PROINPA, generó nuevas variedades de papa mediante el uso de variedades y especies silvestres para la introgresión de genes valiosos a través de mejoramiento genético convencional. Estas variedades obtenidas cuentan con diversas resistencias a enfermedades y virus (Gabriel et al. 2011).
Por otra parte, la selección asistida por marcadores moleculares (SAM), podría facilitar la selección precoz de los genes de interés para su posterior introducción a las variedades, ya que la mayoría de los rasgos importantes en los cultivos como producción, calidad y formas de resistencia a enfermedades, son controlados por genes que pueden asociarse a estos marcadores (Mosquera et al. 2008, Gebhardt et al. 2006).
Por lo anteriormente mencionado, el presente trabajo de investigación tuvo como objetivos: (i) Evaluar la resistencia a tizón tardío (P. infestans), verruga (S. endobioticum) y virus PVX en plantas adultas de variedades mejoradas de papa y (ii) Validar los marcadores moleculares GP94, RB, CP60 y NL25 para co-localizarlos con los genes mayores de resistencia de tizón, verruga y PVX respecti-vamente.
Materiales y métodos
Ubicación. La investigación se realizó entre los años 2010 al 2012 en instalaciones del Laboratorio de Biología Molecular y Bioinformática y en un invernadero de la Fundación para la Promoción e Investigación de Productos Andinos (PROINPA), ubicada en la zona de El Paso, a 15 km de la ciudad de Cochabamba en la provincia de Quillacollo a 17° 18 de latitud Sud y 66° 14 de longitud Oeste, a una altitud de 2540 msnm.
Material biológico. Fueron sembrados en dos campañas consecutivas (2010-2011 y 2011-2012) en invernadero, 18 variedades mejoradas de papa y dos variedades testigo - susceptibles (Waych´a - S. andigena y Desirée - S. tuberosum) (Tabla 1), proporcionados por el programa de mejoramiento genético de la Fundación PROINPA (Gabriel et al. 2011).
Análisis de la resistencia en plantas. Antes de la siembra se realizó un tratamiento de los tubérculos con giberelina a 50 ppm (0.05 g de ácido giberélico en polvo en 1 L de agua) durante 10 min para inducir a la brotación. Posteriormente se sembraron los tubérculos de las 20 variedades de papa en un sustrato estéril compuesto por chala de arroz, arena lama y tierra vegetal en una proporción 1:1:1; se sembraron cuatro tubérculos de cada variedad. Se usaron variedades testigo como: Runa Toralapa, resistente para el virus PVX y la verruga, e India como resistente para el tizón tardío; y se usó la variedad Waych'a como susceptible para todos los patógenos evaluados.
Resistencia a tizón tardío (P. infestans). El análisis de resistencia al tizón tardío (P. infestans) se realizó en foliolos sueltos dentro de placas Petri con agar-agua tal como lo recomiendan Plata (1998) y Vleeshowers et al. (1999), y comprendieron: La preparación del medio agar agua, la colecta de foliolos sanos, la obtención del inoculo y la inoculación del tizón tardío.
Preparación del agar agua. Se diluyeron 15 g de agar agua en 1 L de agua destilada. Se autoclavó la solución de agar agua durante 15 min a 1.5 atm de presión, se dejó enfriar la solución y se procedió al plaqueado con 15 mL de agar agua por placa Petri. Una vez que la solución gelificó, se almacenaron las placas Petri a 4º C.
Colecta de foliolos sanos. Se regaron las plantas a primeras horas de la mañana y se identificó la planta de cada variedad que tenía mejor follaje. Se colectaron 6 foliolos del tercio superior de la planta en bolsas plásticas previamente etiquetadas. Se sumergieron los foliolos en una solución de hipoclorito de sodio al 1.3% durante 1 min, se enjuagaron los foliolos en agua destilada y se los puso sobre papel toalla para secarlos. Se colocaron los foliolos con el envés hacia arriba en las placas Petri con agar agua invertidas, se colocó los foliolos en las placas de acuerdo al diseño experimental.
Obtención del inoculo. Se colectaron hojas de papa con síntomas de tizón tardío (raza compleja del tipo de apareamiento A2) en la zona de Colomi, Cochabamba. Se lavó el micelio de la superficie de las hojas con un chorro potente de agua destilada y se filtró el agua. Se midió la concentración del inoculo con un hemocitómetro, y se llevó la concentración a 40 μL de suspensión esporangial (40000 μL/mL).
Inoculación del tizón tardío en foliolos. Los foliolos sueltos fueron distribuidos en placas Petri en un diseño completamente aleatorio, con cuatro unidades experimentales por cada variedad. Para la inoculación se depositó una gota de 40µL de suspensión esporangial en el envés de los foliolos, se evaluaron 4 foliolos por variedad de papa y se depositaron 2 foliolos por placa Petri. Se iluminaron las placas con focos fluorescentes de 40 Watts y 50-60 Hertz durante el día. Para la evaluación de severidad del tizón tardío, se tomaron fotos de los foliolos y la lesión causada por el oomycete al 4to, 5to y 6to día, después de la inoculación (Plata 1998).
Resistencia a la verruga (S. endobioticum). Se utilizó el patotipo 1, recolectada de tubérculos en la zona de Colomi, Cochabamba. Para evaluar la resistencia en invernadero, se sembraron en macetas cuatro tubérculos por cada una de las 20 variedades de papa, dando un total de 60 macetas. Se distribuyeron las macetas en diseño completamente aleatorio. Al momento de la cosecha se midió con una regla graduada el tamaño de las verrugas encontradas, y se comparó con la escala de daño: 1=sanos sin verruga, 2=trazas con verrugas de 3 mm de diámetro, 3=regular con verrugas mayores a 3 mm y menores a 10 mm de diámetro y 4=severo con verrugas de 10 mm de diámetro o más (Camargo 1999).
Resistencia al virus PVX. Para evaluar la resistencia de la variedad al virus PVX, se inocularon las plantas con el serotipo PVXA, enviado en hojas infectadas desde el Centro Internacional de la Papa (CIP) del Perú.
Previo a la inoculación se frotó las hojas con carborundo, para romper los tricomas y facilitar el ingreso del patógeno. Luego se froto la savia con el virus en cada hoja tratada. Fueron inoculadas cuatro plantas por variedad, las mismas que se distribuyeron en un diseño completamente aleatorio. La evaluación de severidad se realizó midiendo la absorbancia mediante DAS-ELISA (Sosa-Moss et al. 1997).
Análisis molecular: Genes/QTLs de resistencia. Se analizó la presencia de dos genes/QTLs de resistencia al tizón tardío (P. infestans), un gen de resistencia a la verruga (S. endobioticum) y un gen de resistencia al virus PVX, localizados en diferentes cromosomas de la papa (Tabla 2).

Extracción de ADN
Se colectaron y almacenaron a -20° C foliolos de las 20 variedades de papa. Se molió el tejido foliar con nitrógeno líquido a -195° C y se realizó la extracción de ADN genómico siguiendo el protocolo de CTAB propuesto por Doyle & Doyle (1990), Ghislain et al. (1998).
Cuantificación de ADN genómico. La cuantificación de ácidos nucleicos se realizó mediante electroforesis en geles de agarosa al 1%. Se usó el fluoroforo SYBR Green y se sometió las muestras a 90 V por 45 min. Posteriormente se visualizó con el transluminador UV marca Biorad y se comparó la intensidad de las bandas con el marcador de 10000 pb de la firma Eurogentec.
Amplificación mediante PCR. Se realizó un PCR Hot Start en gradiente para la determinación de la Temperatura de anillamiento (Tm) para los iniciadores de cada marcador, basándose en el tamaño de pares de bases de cada primer (Tabla 3).

Se depositó en cada pocillo 4 ng de ADN molde, 1X de tampón PCR 10X, 0.2 mM de DNTPs, 1 pmol/µL de cada primer y 0.025 U/µL de la enzima Taq polimerasa. Se utilizó el aparato PCR (PTC-100 Thermal cycler) y se inició el programa de ciclaje de 1 ciclo a 94° C por 1 min, 35 ciclos de 94° C por 1 min, Tm por 45s y 72° C por 5 min. Los productos amplificados fueron cuantificados mediante electroforesis en geles de agarosa al 1.8%.
El marcador CP60 tipo CAPS fue posteriormente sometido a digestión de los productos amplificados con la enzima de restricción Ddel. Para ello se utilizó 5ng/ µL de ADN molde, Buffer ER 10X, BSA 10 µg/µL y 10 u/µL de la enzima (Makula & Meagher 1980). Los productos digeridos fueron cuantificados en geles de agarosa al 2%.
Análisis estadísticos. Para el tizón tardío, se calculó el área bajo la curva de progreso de P. infestans (AUDPC) con los datos de daño en foliolos. Para que las variedades sean estadísticamente comparables, se calculó el Área Bajo la Curva de Progreso de P. infestans relativa (AUDPCrel) (Bonierbale et al. 2008). Se realizó un Análisis de varianza para los datos obtenidos usando el programa estadístico SAS, y posteriormente se realizó la prueba de Tukey para observar grupos de similitud (SAS 2004).
Para el análisis de la verruga, Se realizó un análisis de varianza con el Software SAS y se aplicó la prueba de Tukey para ver grupos de similitud (SAS 2004).
Para el ensayo con el virus PVX, se realizó un análisis de varianza con los datos de absorbancia y se agruparon las variedades con la prueba de Tukey (SAS 2004).
Para determinar si cada marcador molecular está o no co-localizado y cercano con su respectivo gen de resistencia se realizó un análisis comparativo del nivel de resistencia de las variedades con el alelo o banda expresada al aplicar cada marcador molecular. Para éste análisis se utilizó la prueba de T-test del SAS (2004).
Resultados
El análisis de varianza para las tres enfermedades (Tabla 4) mostró diferencias notables para la variable variedades, esto está indicando que al menos una variedad mostró diferencias notorias para AUDPCrel en tizón, severidad en verruga y absovancia y severidad para PVX. Los análisis realizados mostraron que los rangos del coeficiente de variación (CV) estuvieron entre 21 a 32%, que denota una homogeneidad de los datos y que no requirieron transformación. Los coeficientes de determinación (R2) estuvieron entre 0.66 a 0.87; esto indicó que el modelo estadístico usado explicó el 66% a 87% de la variación y el 34% y 13% de la variación fue debida a efectos ambientales. Por lo que, el modelo fue apropiado para explicar la variación existente en los datos.

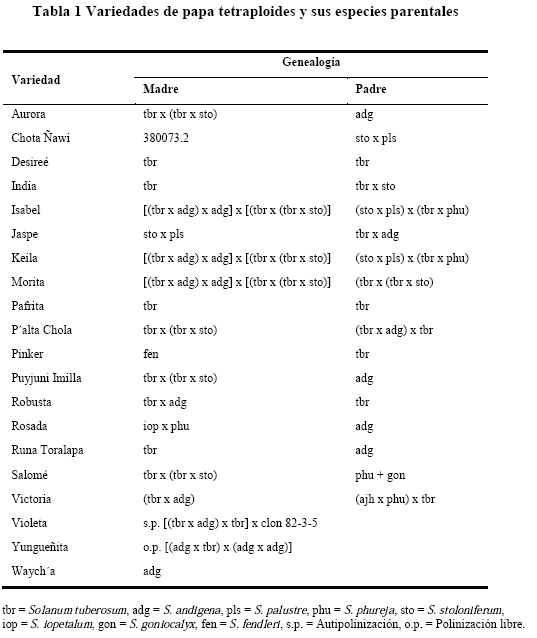
Análisis de la resistencia a tizón
El análisis mediante la prueba de Tukey para el AUDPCrel, se determinó que hubo cinco grupos diferentes de resistencia (a-e) (Figura 1). La mayor resistencia fue observada en las variedades Jaspe y Robusta (AUDPCrel de 2.01 y 2.41), seguida por la variedades India, Isabel y Salomé (AUDPCrel=4.52-6.19). Las variedades Aurora, Runa Toralapa, Violeta y Palta Chola mostraron una resistencia moderada (AUDPCrel=6.58-8.97). En cambio, se observó una resistencia parcial en las variedades Pinker, Victoria, Pafrita, Desireé, Rosada, Puyjuni imilla y Morita (AUDPCrel=9.77-13.51). La variedad Waycha fue susceptible al tizón tardío (AUDPCrel=14.65).
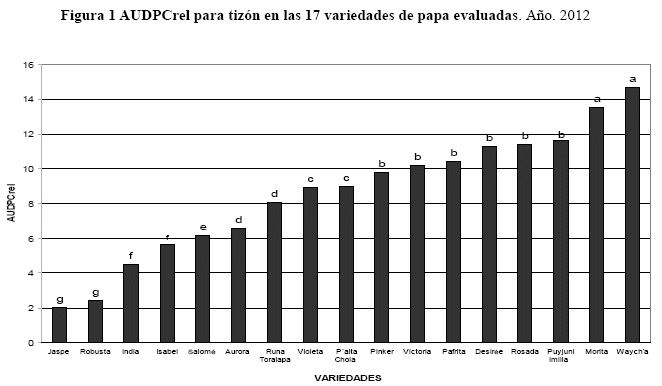
Análisis de la resistencia a verruga
De acuerdo a la escala de daño por verruga (Camargo 1999), se clasificaron las variedades en cuatro grupos de resistencia (1-4). Las variedades con resistencia extrema fueron Aurora, Chota Ñawi, Desireé, India, Isabel, Keila, Morita, Pafrita, P´alta Chola, Puyjuni Imilla, Rosada, Victoria, Violeta, Yungueñita y Runa Toralapa, estas variedades tuvieron una escala de 1. La variedad que presentó resistencia parcial fue Pinker. Las variedades con susceptibilidad moderada fueron Jaspe, Robusta y Salomé. La variedad con susceptibilidad extrema fue el control Waych´a (Figura 2).
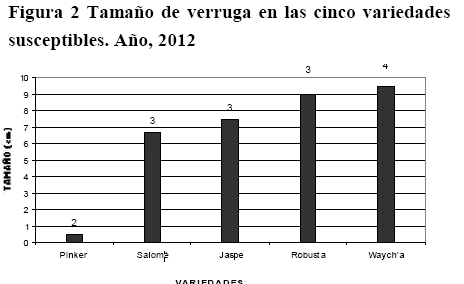
Resistencia al virus PVX
La prueba de Tukey determinó la existencia de cuatro grupos de resistencia (a-d). Las variedades con mayor resistencia fueron Rosada y Runa Toralapa. Luego fueron las variedades Pinker, Pafrita, Robusta, Victoria, Jaspe, Puyjuni Imilla, P´alta Chola, Violeta, Isabel y Keila; con resistencia moderada. Las variedades Chota Ñawi, Yungueñita, Morita, Waych´a, Aurora, Desireé e India mostraron resistencia parcial. La variedad Salomé mostró ser susceptible (Figura 3).
Análisis de alelos
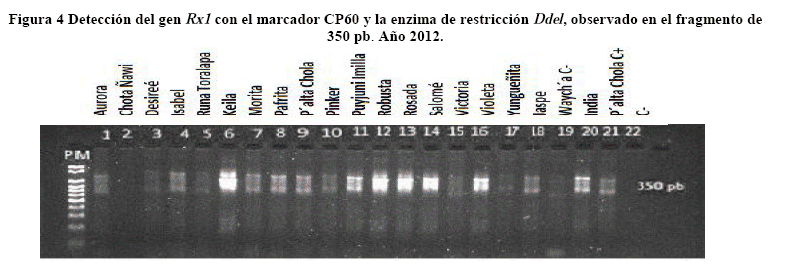
Después de la amplificación con los respectivos primers, se observó la presencia/ausencia de las bandas en los geles de electroforesis de cada uno de los marcadores en las variedades estudiadas (Tabla 5).
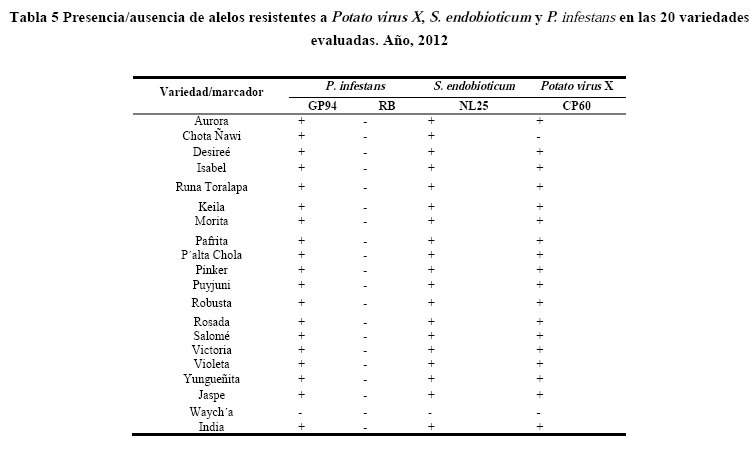
El alelo para el marcador GP94 fue co-localizado con el gen/QTL Rpi-phu1 para resistencia a P. infestans ubicado en el cromosoma IX a 350 pb (Sliwka et al. 2010). El alelo fue detectado en todas las variedades, con excepción del control susceptible Waycha (Figura 6).
El análisis estadístico mediante la prueba de t-Test al Pr<0.05 de probabilidad mostró significancia visible de la diferencia (Dif) entre las medias de V1 y V0, indicando esto que el marcador aparentemente está cercano al gen/QTL Rpi-phu1.
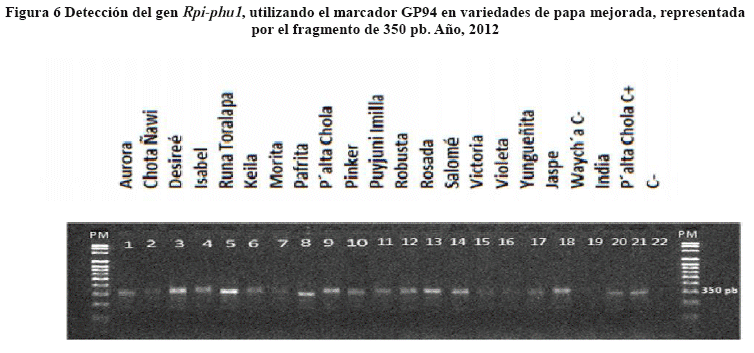
El gen Sen1 fue co-localizado mediante PCR con el marcador NL25, como lo describe Bormann (2004). Los primers amplificaron un fragmento de 1000 pb en todas las variedades excepto el testigo Waych'a (Figura 5).
El análisis estadístico de la diferencia (Dif) entre V1 y V0 para el marcador NL25 mediante la prueba de t-Test al Pr<0.01 de probabilidad, mostró diferencias altamente notables, indicando esto que el marcador aparentemente esta cercano al gen Sen1 (Tabla 6).

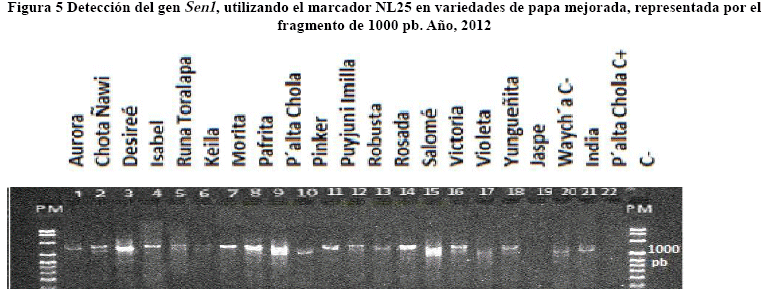
El gen Rx1 ubicado en el cromosoma XII fue co-localizado con el marcador CP60 tipo CAPS, después de su restricción con DdeI, de acuerdo a lo sugerido por Bendahmane et al. (1997). Un fragmento de 350 pb fue observado en todas las variedades evaluadas, con excepción de la variedad Chota Ñawi y el testigo Waych´a (Figura 4).
El análisis estadístico mediante la prueba del t-Test al Pr<0.05 de probabilidad de la diferencia (Dif) entre V1 y V0 del marcador CP60 mostró diferencias marcadas, indicando esto que el marcador aparentemente está cercano al gen/QTL Rx1 (Tabla 6).
Discusión
La resistencia al tizón tardío es poligénica y está gobernada por muchos genes, sin embargo se ha observado resistencia parcial conferida por genes menores de algunas especies cultivadas como S. phureja y S. goniocalyx (Gabriel et al. 2011).Esta resistencia probablemente fue conferida de las especies de S. andigena, S. phureja, S. iopetalum, S. fendleri, S, palustre y S. stoloniferum, utilizados en la generación de las nuevas variedades, y por ello el 100% de las variedades mostró resistencia al tizón tardío.
Sin embargo, se sebe mencionar que se observó que la variedad Desireé mostró el alelo de resistencia al tizón tardío. Esta variedad es susceptible al tizón tardío en follaje, pero fue reportada con la más alta resistencia en tubérculo (Baarveld et al. 2003).
Esto está indicando que la resistencia de hoja y tubérculo son eventos independientes gobernados por genes diferentes (Collins et al. 1999, Oberhagemann et al. 1999, Gabriel et al. 2009). Aparentemente, en el presente estudio la banda del alelo observado está asociada con los genes de resistencia en tubérculos.
El marcador RB utilizado para detectar el gen de resistencia RB al tizón tardío, no se observó en ninguna de las variedades evaluadas, debido a que éstas no presentan los genes de Solanum bulbocastanum que es la especie en la que se ha identificado este gen, por lo que podríamos decir que el marcador es específico para este gen (Colton et al. 2006).
La resistencia a la verruga es monogénica y está gobernada por genes R mayores (Fernandez-Northcote 1991). Todas las variedades evaluadas para esta enfermedad, es decir el 100%, mostraron el alelo de resistencia, probablemente por el gen Sen1 que fue hallado en la especie S. andigena, lo que permitió que todas las variedades mostraran resistencia a esta enfermedad. Este carácter está gobernado por un gen dominante y fue detectado en el cromosoma XI en una posición similar al gen Ryadg (Hehl et al. 1999) y el gen Sen1 está ligado a genes homólogos N.
La resistencia al virus PVX es monogénica, y se conocen dos genes dominantes de hipersensibilidad Nxtbr y Nbtbr . El gen Nbtbr ha sido observado en S. andigena, S. tuberosum, S. acaule, S. vernei, S. sparsipilum, S. chacoense, S. microdontum, S. etuberosum y S. pinnatisectum (Fernandez-Northcote 1991). En este estudio, 18 de las 19 variedades evaluadas, es decir el 95%, sin contar el control negativo Waych´a, mostraron resistencia a este virus, probablemente proveniente de las especie S. andigena y S. palustre (antes llamado S. vernei), utilizadas en la generación de estas. (Veramendi et al. 2011).
Los resultados mostraron que los marcadores moleculares utilizados para asociar los genes de resistencia a múltiples factores fue un método eficiente para asociar con los rasgos de resistencia en las variedades evaluadas. La utilización de marcadores ligados a los loci mayores de resistencia: NL25 y CP60 lograron asociarse con los genes de resistencia Sen1 y Rx1 respectivamente.
Se debe enfatizar que sólo se puede diferenciar genes/QTL seleccionables. Esto requiere que tanto el marcador sea polimórfico, como que la configuración alélica del gen/QTL sea apropiada.
Los marcadores directos permiten la identificación directa de los alelos de un gen de interés y se pueden aplicar independientes del entorno genético, son normalmente derivados de regiones codificantes (coding regions) como ESTs, TDFs, utilizando diferentes técnicas moleculares. Los genes candidato son interesantes en este sentido, pero tienen la desventaja de que no son tan polimórficos como los SSRs (Gabriel et al. 2009).
Por otra parte, existen varias aplicaciones prácticas para genes candidatos detectados como análisis de su diversidad alélica, transformación genética o referencias cruzadas a través de la integración en mapas de ligamiento. Los estudios de diversidad alélica en diferentes germoplasmas puede permitir asociar variantes alélicas específicas con un genotipo particular detectar los alelos más efectivos y de esta manera proporcionar marcadores útiles para la selección asistida por marcadores (SAM).
En el presente estudio se validaron marcadores moleculares co-localizados con genes de resistencia al tizón tardío (P. infestans), la verruga (S. endobioticum) y al virus PVX (Potato Virus X), en los cromosomas IX, XI y XII respectivamente. Estos marcadores fueron validados en nuevas variedades de papa obtenidas de la combinación de diversas especies cultivadas y silvestres de papa.
Las especies cultivadas utilizadas en los cruzamientos involucraron a Solanum andigena (4x), S. tuberosum (4x), S. goniocalyx (2x), S. phureja (2x) y S. ajanhuiri (2x), y a las especies silvestres S. stoloniferum (4x), S. iopetalum (6x), S. palustre (2x), S. fendleri (2x). Siendo éstas consideradas como los potenciales parentales (Gabriel et al. 2011).
Conflictos de intereses
Esta investigación no presenta conflictos de interés.
Agradecimientos
Se agradece al apoyo económico de los proyectos: Fortaleciendo capacidades de innovación participativa para luchar contra la pobreza rural (IP – Holanda) y Fontagro-CLIPAPA.
Literatura citada
Alvarez V, Plata G, Gandarillas A. Enfermedades causadas por virus. 77-82 pp. in Antonio Gandarillas & Noel Ortuño Eds.: Compendio de enfermedades, insectos, nemátodos y factores abióticos que afectan el cultivo de papa en Bolivia. Fundación PROINPA Ed. Cochabamba, Bolivia. 2009.
Baarveld HR, Peeter HMG, Shipper E, Shipper JK, Delleman J. Netherlands calalogue of potato varieties. NIVAP, The Netherlans. 2003; 263 pp.
Bendahmane A, Kanyuka K, Baulcombe DC. High-resolution genetical and physical mapping of the Rx gene for extreme resistance to potato virus X in tetraploid potato. Theor Appl Genet. 1997; 95: 153-162. [ Links ]
Bonierbale M, De Haan S, Forbes A. Procedures for Standard evaluation trials of advanced potato clones. In: An International Cooperators Guide. International Potato Center. Lima, Perú. 2008; 124 pp. [ Links ]
Bormann C. Genetic and molecular analysis of cualitative and cuantitative late blight resistance in tetraploid potato. Tesis Doctoral. University of Hohenheim. 2004; 114 pp. [ Links ]
Camargo T. Control químico de la verruga (Synchytrium endobioticum), y rizoctoniasis (Rhizoctonia solani) de la papa, mediante la utilización de nuevos fungicidas. Tesis de Licenciatura. Universidad Técnica de Oruro-UTO. Oruro, Bolivia. 1999.
Collins A, Milbourne D, Ramsay L, Chatot-Balandras C, Oberhagemann P, De Jong W, et al. QTL for field resistance to late blight in potato are strongly correlated with maturity and vigour. Mol Breed. 1999; 5(5): 387-398. [ Links ]
Colton L, Groza H, Wielgus S, Jiang J. Marker-Assisted Selection for the Broad-Spectrum Potato Late Blight Resistance Conferred by Gene RB Derived from a Wild Potato Species. Crop Sci. 2006; 46:589-594. [ Links ]
Doyle JJ, Doyle JL. A rapid total DNA preparation procedure for fresh plant tissue. Focus. 1990; 12:13-15. [ Links ]
Estrada N. La biodiversidad en el mejoramiento genético de la papa. La Paz, Bolivia. PROINPA/CID/CIP. 2000; 372 pp.
Fernandez-Northcote EN. Mejoramiento por resistencia a los principales virus de la papa. Rev Latinoam Papa. 1991; 4:1-21. [ Links ]
Gabriel J, Pereira R, Gandarillas A. Catálogo de nuevas variedades de papa en Bolivia. Fundación PROINPA. Cochabamba, Bolivia. 2011; 55 pp. [ Links ]
Gabriel J, Ruiz de Galarreta JI, Hernández M, Plata G, Barandalla L, López R & Ritter E. Aplicación de marcadores moleculares para cribado de QTLs en diferentes fuentes de resistencia a tizón tardío (Phytophthora infestans) en papa. Rev Agricul. 2009; 45 (61): 36-43. [ Links ]
Gabriel J. Documento marco: Estrategias y perspectivas del mejoramiento genético de papa (Solanum tuberosum L.) En Bolivia. Fundación PROINPA, Cochabamba, Bolivia. 2010; 60 pp. [ Links ]
Gebhardt C, Bellin D, Henselewski H, Lehmann W, Schwarzfischer J, Valkonen JPT. Marker-assisted combination of major genes for pathogen resistance in potato. Theor Appl Genet. 2006; 112: 1458–1464. [ Links ]
Ghislain M, Zhang D, Herrera MR. Eds. Protocolos de Laboratorio de Biología Molecular. Tipificación genética. Manual de Capacitación. Centro Internacional de la Papa (CIP). 2da Edición. Lima, Perú. 1998; 39 pp.
Hehl R, Faurie E, Hesselbach J, Salamini F, Whitham S, Baker B, et al. TMV resistance gene N homologues are linked to Synchytrium endobioticum resistance in potato. Theor Appl Genet. 1999; 98:379–386.
Makula RA, Meagher RB. A new restriction endonuclease from the anaerobic bacterium, Desulfovibrio desulfuricans, Norway. Nucleic Acids Res. 1998; 8(14): 3125–3131.
Mosquera T, Fernández C, Martínez L, Acuña A, Cuéllar D. Genética de la resistencia de la papa (Solanum tuberosum) a patógenos. Estado de arte. Agron Colomb. 2008; 26(1): 7-15.
Navia O, Plata G, Equise H, Gabriel J, Gandarillas A. Tizón tardío (Phytophthora infestans). Páginas 64-71. In Antonio Gandarillas y Noel Ortuño Eds.: Compendio de enfermedades, insectos, nemátodos y factores abióticos que afectan el cultivo de papa en Bolivia. Fundación PROINPA Ed., Cochabamba, Bolivia. 2009.
Nayudu MV. Plant viruses. Tata Mc Graw Hill. New Delhi, India. 2008; 1249 pp.
Oberhagemann P, Chalot-Balandras C, Bonnel E, Schäfer-Pregl R, Wegener D, Palomino C, et al. A genetic analysis of quantitative resistance to late blight in potato: Towards marker assited selection. Mol Breed. 1999; 5: 399-415. [ Links ]
Ochoa CM. Las papas de Sudamérica: Bolivia. CIP/COSUDE/CID/IFEA. 1ra Edición. La Paz, Bolivia. 2001; 513 pp. [ Links ]
Plata G. Fenotipos de virulencia en Morochata y tipo sexual de apareamiento en Bolivia de Phytophthora infestans que afectan el cultivo de la papa (Solanum tuberosum). Tesis de grado para obtener el título de Ingeniero Agrónomo. Universidad Mayor de San Simón - UMSS. Cochabamba-Bolivia. 1998; 193 pp.
Rojas J, Malosetti M, Van Berloo M. Uso de marcadores moleculares en el mejoramiento genético de las plantas. Fundación PREDUZA-WU. Quito, Ecuador. 2007; 199 pp. [ Links ]
SAS. The SAS system. Version 9.2. SAS OnlineDoc. HTML. Format, SAS Institute, Cary, NC, USA. 2004.
Sliwka J, Jakuczun H, Kamiñski P, Zimnoch E. Marker-assisted selection of diploid and tetraploid potatoes carrying Rpi-phu1, a major gene for resistance to Phytophthora infestans. J Appl Genet. 2010. 51 (2): 133–140. [ Links ]
Sosa-Moss C, Perdomo F, Brathwaite C, Salazar J. Manual de técnicas para el diagnóstico de las enfermedades de las plantas. Instituto Interamericano de Cooperación para la Agricultura IICA. México. 1997; 223 pp. [ Links ]
Torres H. Manual de las enfermedades más importantes de la Papa en el Perú. Centro Internacional de la Papa (CIP). Lima, Perú. 2002; 60 pp.
Veramendi S, Baldelomar M, Terán A, Gabriel J. Marcadores moleculares asociados a genes/QTLs de resistencia para factores bióticos en nuevas variedades de papa (Solanum tuberosum L.) de Bolivia. Rev Latinoam Papa. 2011; 16 (2): 209 - 232. [ Links ]
Vleeshouwers VGAA, van Dooijeweert W, Keizer LCP, Sijpkes L, Govers F, Colon LT. A laboratory assay for Phytophthora infestans resistance in various Solanum species reflects the field situation. Eur J Plant Pathol. 1999; 105:241-250. [ Links ]
Zeballos H, Balderrama F, Condori B, Blajos J. Economía de la papa en Bolivia (1998-2007). Fundación PROINPA. Cochabamba, Bolivia. 2009; 129 pp. [ Links ]
______________