








Serviços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
 Citado por SciELO
Citado por SciELO Links relacionados
 Similares em
SciELO
Similares em
SciELO  uBio
uBio Compartilhar

 Permalink
PermalinkJournal of the Selva Andina Research Society
versão On-line ISSN 2072-9294
J. Selva Andina Res. Soc. v.4 n.1 La Paz 2013
Artículo Original
Uso de marcadores moleculares microsatelite para el análisis de la diversidad genética de papa nativa de Bolivia
Using simple sequence repeats molecular markers to analyze the genetic diversity of potato landraces from Bolivia
Cadima Ximena1*, Veramendi Silene1, Gabriel Julio1
1Fundación PROINPA, Regional Valles Norte, Casilla 4285, Cochabamba, Bolivia.
*Dirección de contacto: Ximena Cadima Fundación PROINPA, Casilla 4285, Cochabamba, Bolivia. Tel: (591) 4 4319595, Fax: (591) 4 4319500.
E-mail address: x.cadima@proinpa.org
Recibido Marzo, 2013.
Devuelto Junio 2013
Aceptado Julio, 2013.
Disponible en línea, Agosto, 2013.
Resumen
Bolivia posee una enorme diversidad de papa cultivada y sus parientes silvestres por ser parte del centro de origen de este cultivo; sin embargo, esa diversidad genética en la colección boliviana de papa no es bien conocida, lo que evita que sea utilizada de manera más eficiente. Con el objetivo de conocer mejor la diversidad de la colección, se realizó un estudio utilizando un set de 22 marcadores microsatélites para la caracterización molecular de seis especies de papa cultivada: Solanum tuberosum Subsp. andigena (991 accesiones), S. x ajanhuiri (56 accesiones), S. x curtilobum (78 accesiones), S. x juzepczukii (116 accesiones), S. stenotomum (228 accesiones) y S. goniocalyx (5 accesiones). Los resultados muestran que la diversidad genética varía en función de la especie, presentando en orden de mayor a menor diversidad a Solanum tuberosum Subsp. andigena, S. stenotomum, S. x curtilobum, S. x juzepczukii, S. x ajanhuiri y S. goniocalyx. En base a los datos moleculares, se encontró además alrededor de 60 accesiones potencialmente duplicadas, y se identificaron los microsatélites STM 5114, STG0010, STM 0037 y STM1104 como los más polimórficos, con valores de PIC de 0.74, 0.73, 0.72 y 0.69 respectivamente.
Palabras clave:
Accesiones, diversidad genética, microsatélites, especies, duplicado.
Abstract
Bolivia has a huge diversity of cultivated potato and its wild relatives for being part of the center of origin of this crop; however, that genetic diversity in the Bolivian potato collection is not well known, preventing its more efficient use. With the aim to better understand the collection diversity, a study was conducted using a set of 22 simple sequence repeats (SSR) for molecular characterization of six species of cultivated potato: Solanum tuberosum Subsp. andigena (991accessions), S. x ajanhuiri (56 accessions), S. x curtilobum (78 accessions), S. x juzepczukii (116 accessions), S. stenotomum (228 accessions) and S. goniocalyx (7 accessions). Results show that genetic diversity varíes with the species, presenting from highest to lowest diversity to Solanum tuberosum ssp. andigena, S. stenotomum, S. x curtilobum, S. x juzepczukii, S. x ajanhuiri and S. goniocalyx. Based on molecular data, about 60 potentially duplícate accessions were also found and SSRs STM 5114, STG0010, STM 0037 and STM1104 were identified as the most polymorphic, with PIC valúes from 0.74, 0.73, 0.72 and 0.69, respectively.
Kev words:
Accessions, genetic diversity, simple sequence sepeats, species, duplicates.
Introducción
Bolivia es uno de los países más ricos en biodiversidad y forma parte del genocentro andino de origen y domesticación de cultivos importantes como la papa. Según Hawkes (1990), el área de diversidad de la papa cultivada y el punto de origen de la domesticación (alrededor de 8000 años a.C.) se encuentran en el área central de los Andes de Bolivia y Perú. Hoy en día, en la región Andina aún existe una amplia diversidad de cultivares nativos de papa concentrados en microcentros de diversidad mantenidos por familias campesinas conservacionistas. En Bolivia algunos de estos microcentros están en la zona de Colomi, ubicada en provincia Chapare en Cochabamba (Terrazas & Valdivia 1998, García & Cadima 2003), en Llallagua en el Norte de Potosí (Terrazas et al 2008), y en el altiplano Norte de La Paz (Iriarte et al 2009). Gran parte de la diversidad de papa cultivada fue colectada en siete de los nueve departamentos de Bolivia, actualmente es mantenida en el Banco Nacional de Germoplasma de Tubérculos Andinos en campo e in vitro. El Banco de Germoplasma de Tubérculos y Raíces Andinas estuvo custodiado por la Fundación PROINPA por mandato del Gobierno Boliviano hasta julio de 2010. A partir de entonces está bajo la custodia del Instituto Nacional de Innovación Agropecuaria y Forestal (INIAF).
La colección de papa que actualmente cuenta con más de 1600 accesiones de siete especies cultivadas (S. tuberosum Subsp. andigena. andigena, S. phureja, S. goniocalyx, S. stenotomum, S. x ajanhuiri, S. x curtilobum y S. x juzepczukii), fue caracterizada morfológicamente y 19 evaluada por sus cualidades agronómicas (Cadima et al 2009).
Para complementar los estudios de diversidad, se vio la necesidad de utilizar otras herramientas como los marcadores moleculares (Rojas et al 2007).
Los marcadores moleculares permiten realizar análisis de variabilidad directamente a nivel del ADN, el cual no es influenciado por el medio ambiente, además pueden ser utilizados en cualquier fase del desarrollo de la planta. Esto evita las ambigüedades inherentes a los marcadores morfológicos y/o agronómicos (de Vicente & Fulton 2004).
Se eligieron los marcadores tipo microsatélites (Simple Sequence Repeats o SSRs en inglés) por sus cualidades en estudios previos en papa tales como investigaciones filogenéticas (Ghislain et al 2009a, Merino-Méndez 2006), estudios taxonómicos (Spooner et al 2007, Lara-Cabrera & Spooner 2005), diversidad genética (de Haan et al 2010, Ghislain et al 2009b, Ghislain et al 2004), identificación de clones y cultivares de papa (Pérez 2004), y marcadores asociados a resistencia de enfermedades (Ghislain et al 2001). Los microsatélites son segmentos cortos de ADN de 1-6 pb, que se repiten en tamden y de forma aleatoria en el genoma.
Estos marcadores son herramientas para estudios genéticos porque: presentan un elevado grado de polimorfismo, su herencia es mendeliana simple, son codominantes, fáciles de medir y analizar, con una confiabilidad del 100%, repetitivos y automatizables (Milbourne et al 1998).
Considerando lo anteriormente mencionado, la presente investigación tuvo los siguientes objetivos: 1) complementar el estudio de la diversidad genética en seis especies de la colección boliviana de papa cultivada: Solanum tuberosum Subsp. andigena, S. x ajanhuiri, S. x curtilobum, S. x juzepczukii, S. stenotomum, y S. goniocalyx, y 2) determinar la estructura genética de la población correspondiente a cada especie evaluada.
Materiales y métodos
Material biológico. 1474 accesiones de papa pertenecientes a la colección boliviana fueron sujetas al estudio de diversidad con marcadores moleculares, según el detalle siguiente: 991 accesiones de Solanum tuberosum Subsp. andigena, 56 accesiones de S. x ajanhuiri, 78 accesiones de S. x curtilobum, 116 accesiones de S. x juzepczukii, 228 accesiones de S. stenotomum y 5 accesiones de S. goniocalyx.
Extracción de ADN. Los foliólos jóvenes colectados fueron molidos en nitrógeno líquido (-170° C) hasta obtener un polvo fino del que se transfirió 100 mg para el proceso de extracción de ADN, para lo cual se siguió el protocolo CTAB 2X (hexadecil bromuro de trimetil amonio) de Doyle & Doyle (1990) con algunas modificaciones. Para la cuantificación de ADN genómico se utilizó el método de electroforesis en gel de agarosa al 1 % y se visualizaron las bandas con bromuro de etidio.
Amplificación de Microsatélites por PCR. En el estudio fueron utilizados 22 marcadores microsatélites, cuyas características se describen en la Tabla 1.
Tabla 1 Marcadores microsatelites utilizados en la evaluación.
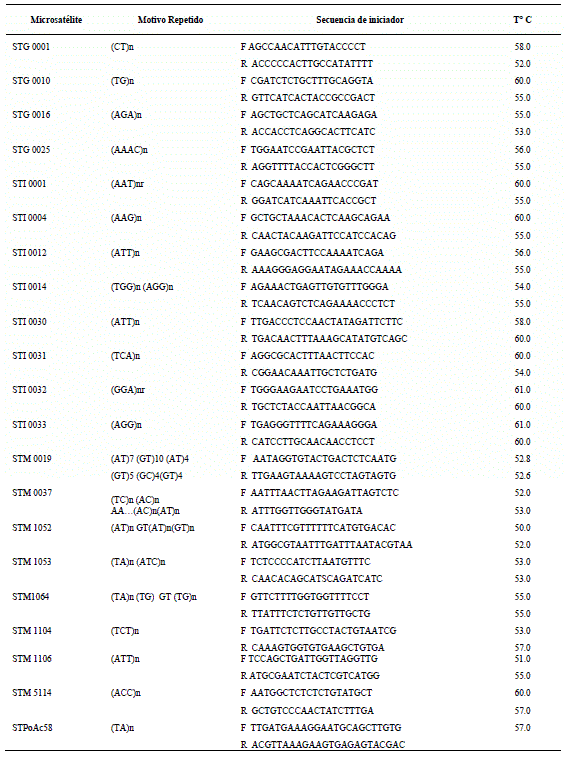
El genotipado con los microsatélites se realizó bajo los servicios del laboratorio de Genotipado del ICRISAT-Patancheru de la India.
Se utilizó el iniciador MI3 con secuencia 5'CACGACGTTGTAAAACGAC3' como cola de extensión en la terminación 5' de cada iniciador "forward" (Oetting et al 1995). Se realizó la PCR en 5 ul de volumen de reacción con 5 ng de ADN, 2.5 mM MgCl2, 0.2 mM de dNTPs, IX PCR buffer, 0.006 pM de M13-tailed forward primer, 0.09 pM de MI3-forward primer marcado con 6-Fam, Vic, Ned o Pet (Applied Biosystems), 0.09 pM del iniciador "reverse" y 0.1 U de Taq DNA polymerase (SibEnzyme Ltd., Russia), en un termociclador GeneAmp® PCR System 9700 (Applied Biosystems, USA) con la siguientes condiciones cíclicas: desnaturación inicial a 94° C por 3 min, seguida por 10 ciclos de desnaturación a 94° C por 1 min, anillamiento a 61 °C por 1 min (temperatura reducida por 1° C para cada ciclo) y extensión a 72° C por 1 min. Esto se siguió por 40 ciclos a 94° C por 1 min, anillamiento a 54° C por 1 min y extensión a 72° C por 1 min con una extensión final de 10 min a 72° C. En base al tamaño esperado de los productos de amplificación, algunos productos PCR se desarrollaron juntos ("multiplex") con un patrón estándar interno (GeneScan 500 LIZ® de Applied Biosystems) y se hizo electroforesis capilar utilizando un equipo ABI 3730x/ Genetic Analyzer (Applied Biosystems, USA). Los datos generados fueron analizados utilizando el software Genemapper (Applied Biosystems, USA) y el tamaño de los fragmentos (alelos) se determinaron en pares de bases (bp) en función a la migración del patrón estándar interno.
Análisis de la diversidad genética y estructura poblacional. Los parámetros considerados para determinar la diversidad genética en base a la amplificación de los microsatélites fueron el número y tamaño en pares de bases de los alelos encontrados en cada especie, así como la frecuencia alélica. La frecuencia alélica se define como una medida de la presencia continua o asidua de un alelo en una población, o de la proporción que representa de todos los alelos de un gen (De Vicente et al 2004).
También fue calculado el índice de Contenido Polimórfico (PIC por sus siglas en inglés) que evalúa cuan informativo en términos de polimorfismo es un marcador en la población de acuerdo a las frecuencias de los alelos (Aranguren-Mendez et al 2005). Sus valores oscilan entre 0 y 1. Para su cálculo se multiplica la probabilidad de cada posible cruzamiento (estimado a partir de las frecuencias alélicas) por la probabilidad de ser informativos, es decir, que se pueda identificar al progenitor del que procede el alelo. Se utilizó la fórmula de Botstein et al (1980).
Las relaciones existentes entre las accesiones en estudio y el análisis de la estructura poblacional se estimaron mediante el análisis de conglomerados. Los datos de los alelos en pares de bases fueron transformados en datos binarios y dispuestos en una matriz denotando la presencia de un alelo como (1) y la ausencia como (0), esta información fue utilizada para calcular matrices de valores de similitud utilizando el coeficiente de Jaccard y se construyeron dendrogramas (Jaccard, 1908, Zambrano et al 2003).
El cálculo del coeficiente de Jaccard se hizo con la fórmula recomendada por Jaccard (1908).
El análisis cluster se realizó por el método de ligamiento promedio UPGMA (Unweighted Pair Group Method with Arithmatic Mean) recomendado por Sokal & Sneath (1973) para generar los dendrogramas.
El cálculo de los valores de similitud, análisis de cluster y visualización de los dendrogramas se efectuó con el programa NTsys v. 2.10p (Rohlf 2000).
Resultados
Tabla 2 Número de accesiones y rango en pares de bases por especie.
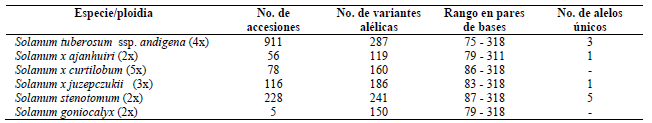
2x: Diploide, 3x: Triploide, 4x: Tetraploide, 5x: Pentaploide.
Tabla 3 Valores del PIC para los 22 microsatelites utilizados en las seis especies de papa.
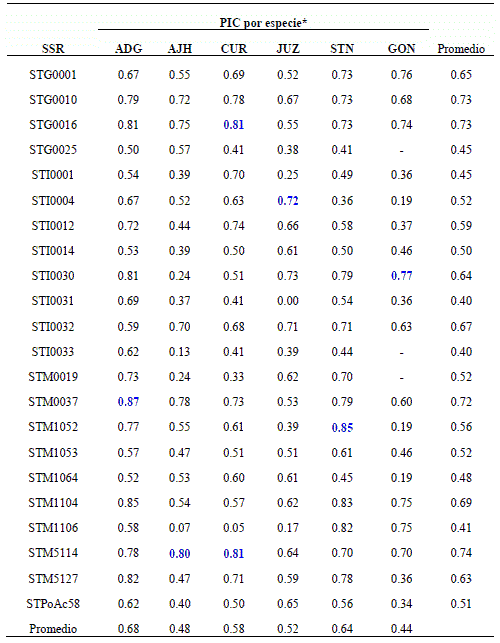
*SSR: Simple Sequence Repeats (Marcador microsatélite), ADG: Solanum tuberosum ssp. andigena; AJH: Solanum x ajanhuiri; CUR: Solanum x curtilobum; JUZ: Solanum x juzepczukii; STN: Solanum stenotomum; GON: Solanum goniocalyx. Los valores de PIC en azul son los mayores para cada marcador y especie.
Figura 1 Dendrograma generado por el programa NTSYS PC 2.10 mediante el análisis de UPGMA y coeficiente de similitud de Jaccard a partir de datos de 22 marcadores microsatélites en 1476 accesiones de seis especies de papa: adg: Solanum tuberosum ssp. andigena (991 accesiones), ajh: S. x ajanhuiri (56 accesiones), cur: S. x curtilobum (78 accesiones), juz: S. x juzepczukii (116 accesiones) y stn: S. stenotomum (228 accesiones).
Discusión
Una de las principales ventajas de los marcadores moleculares tipo microsatélites es la pequeña cantidad de ADN que se necesita para la amplificación, siendo ésta de 5 - 50 ng por reacción (Torrez & Moreno 2001), el protocolo utilizado permitió obtener ADN, (40 a 100 ngl\ú en un volumen de 50 -60 \x\ por muestra) para los análisis moleculares con microsatélites utilizados en el estudio.
Los resultados indican la presencia de 327 alelos diferentes para los 22 microsatélites amplificados en las seis especies de papa evaluadas. En la Tabla 2 se distinguen los resultados por especie en cuanto a número de variantes alélicas, rango de pares de bases y número de alelos únicos encontrados en cada caso.
Los resultados sobre el número y el tamaño de alelos (pb) de cada especie de papa estuvieron dentro de los rangos descritos por el CIP (Ghislain et al 2000).
La mayor diversidad presenta S. tuberosum Subsp. andigena, por la cantidad de alelos detectados, que coincide con reportes, donde los cultivares de esta especie son abundantes y diversos en la región andina (De Haan et al 2010, Ghislain et al 2009b). La mayor cantidad de alelos también tiene relación con el mayor número de accesiones analizadas (n=991) en relación a otras especies, puesto que aumenta la probabilidad de encontrar nuevos alelos que pudieran estar en una frecuencia más baja dentro de una población más grande. Lo anterior indicaría que el número de alelos detectados depende en gran parte de la cantidad y diversidad de germoplasma muestreado (Pérez 2004).
S. stenotomum que a pesar de ser una especie diploide (2x) presenta alta diversidad genética, mucho mayor a otras especies diploides, triploide y pentaploide. La diversidad de esta especie es menor pero comparable a la diversidad de S. tuberosum Subsp. andigena por el número de individuos analizados.
El mayor número de alelos únicos presentes en S. stenotomum (5), es otro indicador de su diversidad alélica y potencial reservorio de nuevos alelos para el mejoramiento genético.
Se corroboró que los microsatélites STPoAC - 58, STM 0037, STM 0031, STM 3012 y STM 2030 pueden ser utilizados para la determinación de la ploidia como lo menciona Ghislain et al (2000) y pueden sugerir posibles errores en la clasificación de germoplasma. Cada microsatélite representa un locus simple en todos los estudios de mapeo poblacional y constantemente revelan altos niveles de ploidia cuando se observa más alelos de los esperados (Ghislain et al 2000).
El índice de Contenido Polimórfico (PIC) para un mismo microsatélite puede variar mucho entre especies (Tabla 3). Los valores por microsatélite estuvieron en un rango de 0.13a0.87yen cada especie el mayor PIC se dio en un microsatélite diferente. Tomando los valores promedios, S. tuberosum Subsp. andigena registró el mayor valor de PIC seguido por S. stenotomum, S. x curtilobum, S. x juzepczukii, S. x ajanhuiri y finalmente S. goniocalyx. Comparando el valor promedio de polimorfismo de cada microsatélite, se observó que los microsatélites STM 5114, STG0010, STG0016, STM 0037 y STM 1104 fueron los más polimórficos. Los valores del PIC obtenidos para los microsatélites en el CIP fueron más altos (STM1104 = 0.798; STM 0037 =0.784) probablemente porque utilizaron una selección de cultivares de la colección mundial de papa con mayor diversidad (Ghislain et al 2000). Por otro lado los marcadores menos polimórficos fueron los microsatélites STI 0033 y STG0025 (Tabla 3). Estos resultados sugieren que para trabajos similares en el futuro se podría utilizar una selección de los marcadores con mayor valor de contenido polimórfico detectados en este estudio para discriminar la diversidad en papas cultivadas bolivianas.
El microsatélite STM 1104 fue uno de los más polimórficos, similares resultados fueron reportados por Ghislain et al (2000) durante el desarrollo de descriptores moleculares para la identificación genotípica de cultivares de papa. Este microsatélite conjuntamente con el STM 0019 han distinguido inequívocamente 20 cultivares de papa de 22 evaluados. Sin embargo, en nuestro trabajo este marcador fue uno de los menos polimórficos.
Análisis de la estructura poblacional. La información generada con los 22 microsatélites para las especies analizadas, fue realizada a través del cálculo de los valores de similitud entre las accesiones utilizando el coeficiente de Jaccard. Con esta información se realizó agolpamientos mediante el método UPGMA, que fueron visualizados en un dendrograma.
Se observa en el dendrograma (Figura 1) generado a partir de las seis especies una estructura genética clara y definida donde se puede observar que la población se divide básicamente en dos grandes grupos A y B a un coeficiente de similaridad de 0.26. El grupo A conformado por las especies S. tuberosum Subsp. andigena , S. stenotomum y S. x curtilobum y S. x ajanhuiri denotando la relación genética entre las especies como ya fue sugerido por Hawkes (1990). El grupo B conformado por las especies, S. tuberosum Subsp. andigena y S. xjuzepczukii.
Ghislain et al (2000) en un estudio similar con 18 microsatélites en 22 cultivares de papa de diferentes especies observó también la relación genética y taxonómica de los materiales estudiados, encontrando que el clúster de especies diploides y triploides se separaron de los clones tetraploides de S. tuberosum Subsp. tuberosum y S. tuberosum Subsp. andigena.
En el grupo A, la estructura poblacional de S. tuberosum Subsp. andigena (991 accesiones) denota la formación de dos grandes grupos I y II con un coeficiente de similaridad de 0.28, y el grupo I además subdividido en dos subgrupos claramente diferenciados con un coeficiente de similaridad de 0.30.
En el mismo grupo A, las 228 accesiones de la especie S. stenotomum no muestran grupos claramente diferenciados como en las otras especies, debido a la gran diversidad que se expresa en más de 25 grupos con un coeficiente de similitud de 0.40, las accesiones de S. x ajanhuiri están estructuradas en tres agrupaciones tomando en cuenta un coeficiente de similitud de 0.47, y se distinguen cinco subgrupos de 10 accesiones potencialmente duplicadas (100 % de similitud). Resalta que la afinidad de las accesiones dentro las tres agrupaciones es coherente con rasgos morfológicos, es el caso del primer grupo de esta especie que está conformado por solo los cultivares del tipo "yari", que según Ochoa (1990), conforman una taxa aparte ("var.") dentro de S. x ajahuiri.
Finalmente, las 116 accesiones de S. x juzepczukii dentro del grupo B, muestran a un coeficiente de similitud de 0.6, tres grupos diferentes, y se identifican ocho ramas con 31 accesiones potencialmente duplicadas.
En resumen el análisis de conglomerados permitió visualizar las diferencias y relaciones genéticas entre especies y dentro especies en cuanto a diversidad. Se corrobora la amplia diversidad de S. tuberosum Subsp. andigena y S. stenotomum porque se distingue mayor disimilitud entre accesiones en comparación con las otras especies. Solanum x ajanhuiri, S. x juzepczukii y S. x curtilobum muestran grupos genéticamente más similares, y también con un número importante de accesiones potencialmente duplicadas. Resultados similares en cuanto a diversidad intraespecífica fueron mencionados por De Haan et al (2010).
El alto polimorfismo presentado en S. stenotomum puede deberse a que es la primera especie cultivada originada probablemente de la silvestre S. leptophyes (Hawkes 1990, Hawkes & Hjerting 1989). Respecto a S. tuberosum Subsp. andigena, el nivel de polimorfismo presentado por esta especie, probablemente se debe a que evolutivamente proviene de un cruce de S. stenotomum y la especie silvestre S. x sparsipilum donde ambas especies (diversas) aportaron gametos no reducidos (2x) formando una especie tetraploide (4x) (Hawkes & Hjerting 1989). Además cultivares de esta especie son los más difundidos en la región Andina y por lo tanto sujetos a una mayor diversificación. S. x juzepczukii y S. x curtilobum tienen menor diversidad debido a su ploidía, lo cual no les permite generar semilla fértil (Estrada 2000), reduciéndose de esta manera su di versificación. S. x ajanhuiri es de distribución más localizada, lo cual explica también su variabilidad reducida.
En conjunto la variabilidad genética en las especies de papa cultivada presente en Bolivia, es crucial para la asegurar la alimentación de los agricultores y las familias campesinas, ante una diversificación de ambientes y riesgos climáticos adversos de la región Andina. Estos materiales además, son fuente invalorable en la búsqueda constante de características importantes en el mejoramiento genético de la papa para las demandas actuales y futuras.
Conflictos de intereses
Esta investigación no presenta conflictos de interés.
Agradecimientos
Se agradece a las diferentes fuentes financieras como el ex Sistema Nacional de Recursos Genéticos para la Agricultura y la Alimentación (SINARGEAA) de Bolivia, el actual Instituto Nacional de Innovación Agrícola y Forestal (INIAF) y FONTAGRO. Agradecemos las valiosas contribuciones del Dr. Jorge Rojas Bertrán en la realización de la presente investigación.
Literatura citada
Arcia A, Rodríguez I. Influencia de diferentes concentraciones de conidios de Trichoderma spp en el control de Sclerotium rolfsii Sacc. En: Compendio de resúmenes del XIII Congreso Venezolano de Fitopatología. 07-11 Noviembre, San Cristóbal, Táchira. Venezuela. 1993;81. [ Links ]
Aranguren-Méndez JA, Román-Bravo R, Isea W, Villasmil Y, Jordana J. Los microsatélites (STR's), marcadores moleculares de ADN por excelencia para programas de conservación: una revisión. Archivos Latinoamericanos de Producción Animal. 2005; l(13):30-42.
Botstein D, White RL, Skolnick M, Davis RW. Construction of a genetic linkage map in man using restriction fragment length polymorphisms. Am J Hum Gen. 1980; 32:314-331. [ Links ]
Cadima X, Terrazas F, Gandarillas A. Los sistemas de conservación de recursos genéticos de Tubérculos y Raíces Andinas: La experiencia de PROINPA. Revista de Agricultura, Bolivia 2009; 43(60):31-36. [ Links ]
De Haan S, Nunez J, Bonierbale M, Ghislain M. Multilevel agrobiodiversity and conservation of Andean potatoes in Central Perú: Species, morphological, genetic, and spatial diversity. Moun Res Develop. 2010; 30:222-231. [ Links ]
De Vicente MC, Fulton T. Tecnologías de marcadores moleculares para estudios de diversidad genética de plantas: Módulos de aprendizaje. En MC de Vicente, T Fulton, eds, Módulos de Aprendizaje sobre Marcadores Moleculares, 1 Ed. Voll. Instituto Internacional de Recursos Filogenéticos (IPGRI), Roma- Italia. 2004.
Doyle JJ, Doyle JL. Isolation of plant DNA from freshtissue. BRLFocus. 1990; 12:13-15. [ Links ]
Estrada N. La biodiversidad en el mejoramiento genético de la papa. PROINPA-CID-CIP, La Paz - Bolivia. 2000. [ Links ]
García W, Cadima X (eds.). Manejo sostenible de la agro biodiversidad de tubérculos andinos: Síntesis de investigaciones y experiencias en Bolivia. Conservación y uso de la biodiversidad de raíces y tubérculos andinas: Una década de investigación para el desarrollo (1993-2003). 1. Fundación para la Promoción y la Investigación de Productos Andinos (PROINPA), Alcaldía de Colomi, Centro Internacional de la Papa (CIP) Agencia Suiza para el Desarrollo y la Cooperación (COSUDE). Cochabamba, Bolivia. 2003; 208 pp. [ Links ]
Ghislain M, Nunez J, Herrera MdR, Pignataro J, Guzman F, Bonierbale M, Spooner DM. Robust and highly informative microsatellite-based genetic identity kit for potato. Mol Breed. 2009b; 23:377-388. [ Links ]
Ghislain M, Nunez J, Herrera MdR, Spooner DM. The single Andigenum origin of Neo-tuberosum potato materials is not supported by microsatellite and plastid marker analyses. Theo Appl Gen. 2009a; 118:963-969. [ Links ]
Ghislain M, Spooner DM, Rodríguez F, Villamon F, Nunez J, Vasquez C, Waugh R, Bonierbale M. Selection of highly informative and user-friendly microsatellites (SSRs) for genotyping of cultivated potato. Theo Appl Gen. 2004; 108:881-890. [ Links ]
Ghislain M, Trognitz B, Herrera MdR, Solis J, Casallo G, Vásquez C, Hurtado O, Castillo R, Portal L, Orrillo M. Genetic loci associated with field resistance to late blight in offspring of Solanum phureja and S. tuberosum grown under short-day conditions. Theo Appl Gen. 2001; 103:433-442. [ Links ]
Ghislain M, Rodríguez F, Villamon F, Núñez J, Waugh R, Bonierbale M. Establishment of Microsatellite Assays for Potato Genetic Identification. CIP Program Report 1999-2000. 2000; 167-174. [ Links ]
Hawkes JG, Hjerting JP. The potatoes of Bolivia: Their breeding and evolutionary relationships Oxford University Press, Oxford, UK. 1989. [ Links ]
Hawkes JG. The potato : evolution, biodiversity and genetic resources. Belhaven. London. 1990. [ Links ]
Iriarte V; Condori B, Parapo D, Acuña D. Catálogo etnobotánico de papas nativas del Altiplano Norte de La Paz-Bolivia. Cochabamba, Bolivia. 2009; 142 pp. [ Links ]
Jaccard P. Nouvelles recherches sur la distribution florale. Bull. Soc. Vaud Sci Nat. 1908; 44:223-270. [ Links ]
Lara-Cabrera S, Spooner DM. Taxonomy of Mexican diploid wild potatoes: (Solanum sect. Petota) morphological and microsatellite data. Monogr Syst Bot Missouri Bot Garden. 2005; 104:199-205. [ Links ]
Merino-Méndez CG. Divergencia de loci microsatelites entre papas silvestres y cultivadas (Familia Solanaceae Género Solanum Sección Petota), Facultad de Ciencias Biológicas, Universidad Nacional Mayor de San Marcos, Lima, Perú. 2006; 78 pp. 29 [ Links ]
Milbourne DRC. Meyer AJ, Collins LD, Ramsay C. Gebhardt, Waugh R. Isolation, characterisation and mapping of simple sequence repeat loci in potato. Mol Gen Genet. 1998; 259:233-245.
Ochoa C. The Potatoes of South America: Bolivia Cambridge Univ. Press, Cambridge, U.K. 1990. [ Links ]
Oetting WS, Lee HK, Flanders DJ, Wiesner GL, Sellers TA, King RA. Linkage analysis with multiplexed short tándem repeat polymorphisms using infrared fluorescence and M13 tañed primers. Genomics. 1995; 30:450-458. [ Links ]
Pérez JR. Evaluación de la diversidad genética de papas nativas (Solanum tuberosum L. ssp. tuberosum Hawkes) silvestres y cultivadas del sur de Chile, mediante el uso de marcadores microsatelites. Tesis de licenciatura. Universidad Austral de Chile, Valdivia - Chile. 2004. [ Links ]
Rohlf FJ. NTYSYS-pc, Numerical Taxonomy and Multivariate Analysis System, Versión 2.10e., Applied Biostatistics, Inc., New York. 2000. [ Links ]
Rojas J, Sánchez Y, Alba E, Cadima X, Franco J, Gandarillas A. Utilización de la tecnología de los marcadores moleculares en la conservación del germoplasma y el mejoramiento genético de la papa: Experiencias en Bolivia. Revista de Agricultura, Bolivia. 2007; 59(40):29-36. [ Links ]
Sokal RR., Sneath PHA. Numerical Taxonomy. Ed. Freeman. San Francisco. 1973. [ Links ]
Spooner DM, Nunez J, Trujillo G, Herrera MdR, Guzman F, Ghislain M. Extensive simple sequence repeat genotyping of potato landraces supports a major reevaluation of their gene pool structure and classification. Proceedings of the National Academy of Sciences of the United States of America. 2007; 104:19398-19403. [ Links ]
Terrazas F, Cadima X, García R, Zeballos J. Catálogo etnobotánico de papas nativas. Tradición y cultura de los Ayllus del Norte Potosí y Oruro. Ricerca & Cooperazione, Unión Europea, Centro de Apoyo a Desarrollo, GTZ, Fundación PROINPA, MDRyMA. Cochabamba - Bolivia. 2008; 189 pp. [ Links ]
Terrazas F, Valdivia G. Spatial dynamics of in situ conservation: handling the genetic diversity of Andean tubers in mosaic systems. Gen Res News. 1998; 114:9-15. [ Links ]
Zambrano AY, Demey JR, Fuenmayor F, Segovia V, Gutiérrez Z. Diversidad genética de una colección de yuca a través de marcadores moleculares RAPDS. Agron Trop, Venezuela. 2003; 2(53): 155-174. [ Links ]