











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkJournal of the Selva Andina Research Society
versión On-line ISSN 2072-9294
J. Selva Andina Res. Soc. v.4 n.1 La Paz 2013
Artículo Original
Validación de marcadores moleculares para resistencia a sequía en trigo harinero (Triticum aestivum L.) bajo condiciones de invernadero
Molecular markers validation to drought resistance in wheat meal (Triticum aestivum L.) under greenhouse conditions
Gabriel Julio1*, Veramendi Silene1, España Pablo1
1Fundación PROINPA, Regional Valles Norte, Casilla 4285, Cochabamba, Bolivia.
*Dirección de contacto: Julio Gabriel
Fundación PROINPA, Casilla 4285, Cochabamba, Bolivia. Tel: (591) 4 4319595, Fax: (591) 4 4319500.
E-mail address: j.gabriel@proinpa.org
Recibido Enero, 2013.
Devuelto Junio 2013
Aceptado Julio, 2013.
Disponible en linea, Agosto, 2013
Resumen
Con objetivo de estudiar la resistencia genética a sequía y validar marcadores moleculares co-localizados con genes/QTLs para este factor, se evaluó 16 variedades y líneas avanzadas de trigo harinero (Triticum aestivum) en dos estadios de desarrollo del cultivo. Se consideraron los parámetros fisiológicos: cantidad de clorofila (clo), grado de marchitez o severidad (sev) y capacidad de recuperación (recup), los parámetros morfológicos: materia seca de follaje (MSF) y materia seca de raíz (MSR), los mecanismos de resistencia integrados: eficiencia de uso de agua (WUE), otros parámetros: número de granos (Ngrano) y peso de granos (Pgrano), parámetros bioquímicos: Catalaza (CAT), Ascorbato Peroxidasa (APX) y Guaiacol Peroxidasa (POX) y tres marcadores microsatélite (Xwmc603, Xwmc596, Xwmc9). Los resultados mostraron diferencias significativas para MSR y Ngrano. Se observó que Anzaldo, ERR2V.L-20, EARII2V.L-5, EARIZV.L-11, ERR2V.L-11 y EE2V.L-19 fueron los más resistentes al estrés hídrico por sequía. Hubo una correlación negativa alta y significativa para clo y sev. En cambio, se observó una correlación moderada y significativa entre las variables clo y MSR. También se observó una correlación alta negativa y significativa entre la recup y la MSR. Finalmente se encontró una alta correlación negativa entre la MSR y el Ngrano. Todas las demás variables mostraron correlaciones bajas y no significativas. En los análisis bioquímicos la variedad Anzaldo mostró una actividad enzimática incrementada respecto de los controles en todos los casos (CAT-APX y POX), siendo la más resistentes al estrés hídrico por sequía. Finalmente se encontró que los marcadores SSR (Xwmc596 y Xwmc9) están co-localizados con el gen/QTL de resistencia a sequía y pueden ser utilizados para la selección asistida por marcadores moleculares.
Palabras clave:
Materia seca, clorofila, severidad, correlación, bioquímico.
Abstract
With the aim to study the genetic resistance to drought and validate molecular markers co-localized with genes/QTLs for this factor, 16 varieties were evaluated as well as advanced lines of wheat meal (Triticum aestivum) in two stages of crop development. Physiological parameters were considered: amount of chlorophyll (clo), wilting or severity degree (SEV) and recovery (reco), morphological parameters: foliage dry matter (FDM) and root dry matter (RDM), the integrated resistance mechanisms: water use efficiency (WUE), other parameters: number of grains (Ngrain) and grain weight (Wgrain), biochemical parameters: Catalaza (CAT), Ascorbate Peroxidase (APX) and Guaiacol Peroxidase (POX) and three microsatellite markers (Xwmc603, Xwmc596, Xwmc9). Results showed significant differences for MSR and Ngrain. It was observed that Anzaldo, ERR2V.L-20, EARII2V.L-5, EARIZV.L-11, ERR2V.L-11 and EE2V.L-19 were the most resistant to drough water stress. There was a highly significant negative correlation between the MSR and Ngrain. All other variables showed low and non-significant correlations. In biochemical analyzes, the Anzaldo variety showed an increased enzymatic activity compared to controls in all cases (CAT-APX and POX), being the most resistant to water stress by drought. Finally, it was found that SSR markers (Xwmc596 and Xwmc9) are co-located with the gene / QTL of drought resistance and can be used for marker-assisted selection.
Key words:
Dry matter, chlorophyll, severity, correlation, biochemistry.
Introducción
El trigo (Triticum aestivum L.), una especie anual adaptada a condiciones diferentes de clima y suelo y es una fuente importante de proteínas e hidratos de carbono para la población boliviana. Un estudio realizado por la ONU-FAO (2010), menciona que la superficie cultivada de trigo en Bolivia es de aproximadamente 182.134 ha (que corresponde a 17% de la superficie cultivada nacional) con un rendimiento promedio en grano de 1.31 t/ha, Las principales zonas productoras de trigo están ubicadas en los valles de Tarabuco y Yamparaez en Chuquisaca, Anzaldo y Carrasco en Cochabamba, Yesera Norte y cercado en Tarija, Pampas de Lequezana en Potosí y Okinawa en la Región Integrada en Santa Cruz, En Okinawa – Santa Cruz, se siembran unos 107000 ha con rendimientos en grano de 1.90 t/ha (ANAPO 2012).
El (MAGDR 2000), indica que en Bolivia más del 80 % de la superficie cultivada es con cereales a secano y la producción nacional de trigo satisface al 20% de la demanda nacional (INE 2011). En estos sitios donde la agricultura es a secano, el estrés hídrico por sequía es uno los principales factores ambientales que limita la producción del trigo. Esto implica que el cultivo se desarrolla durante la época de lluvias o la siembra se realiza al final del periodo lluvioso y el cultivo solamente dispone para su desarrollo, de la humedad almacenada en el suelo antes de la siembra. Los cultivos a secanos producen menos que los cultivos bajo riego y está sujeta frecuentemente a periodos de déficit de agua para el desarrollo del cultivo (Herbas & Montaño 2011).
En los últimos años, como efecto probablemente del cambio climático hay deficiencia en las precipitaciones pluviales y una mala distribución durante el periodo lluvioso, por lo que hay un frecuente déficit hídrico en la mayor parte de los cultivos a secano, este hecho repercute negativamente en el rendimiento de los cultivos poniendo en riesgo la seguridad alimentaria de la población boliviana.
Se sabe que el estrés hídrico por sequía limita la producción de los cultivos. Las pérdidas por estrés hídrico por sequía pueden superar a todas las pérdidas combinadas, causadas por otros factores (Kramer 1980). La intensidad de la respuesta a la sequía depende de la severidad, la duración del mismo y el estadio de desarrollo de la planta en el cual ocurre el estrés (Mukandama et al 2003, Morgan 2004, Frahm et al 2004, Machado 2004).
Una sequía moderada, en la fase de desarrollo vegetativo, puede afectar levemente el crecimiento del cultivo y acelerar la maduración del mismo; en cambio, una sequía durante la floración puede afectar drásticamente la formación del polen y su fertilización, reduciendo significativamente el número de granos (Herbas & Montaño 2011). O’Toole & Moya (1981) observaron que durante la formación de los gametos masculinos y femeninos, la planta es sensible al déficit de agua y el más afectado es el gametofito masculino, que puede llegar a ser estéril.
Cuando el estrés hídrico por sequía, ocurre en etapas tempranas del desarrollo reproductivo, ocasiona una reducción del número de granos debido principalmente a la absorción del ovario o la esterilidad del polen, esto a causa de que se inhibe la fotosíntesis y disminuye la translocación de fotosintatos a los órganos en desarrollo (Boyer & Westgate 2004). Una deficiencia de agua también determina una menor eficiencia de conversión de radiación interceptada en materia seca, lo que contribuye a reducir el número de granos cuando el estrés se produce durante el período de crecimiento de la espiga (Gallagher et al 1976, Herbas & Montaño 2011).
Por otra parte, se sabe que cuando ocurre el estrés abiótico en plantas, se produce la ROS (Reactive Oxygen Species) continuamente en el cloroplasto, mitocondria y peroxisomas. En condiciones normales, la producción y remoción de la ROS está estrictamente controlada por antioxidantes. Sin embargo, el equilibrio entre la producción y la depuración de éstas puede ser perturbado por diversos factores fisicoquímicos, como son el déficit hídrico, la salinidad, las temperaturas extremas, la excesiva o insuficiente radiación luminosa, la anaerobiosis por encharcamiento o inundación, los factores mecánicos como el viento o la compactación del suelo, viento fuerte y las lesiones (Mano 2002).
La alteración del equilibrio entre los peroxidantes y los antioxidantes, incrementa a las primeras y la tasa de producción de ROS que son formas parcialmente reducidas del oxígeno y que puede conducir a la destrucción de los sistemas celulares. Las cantidades excesivas de la ROS pueden dañar proteínas, lípidos y ácidos nucleicos. Por lo tanto, el incremento en la capacidad de eliminación de estas ROS por parte de la planta es considerada como síntoma de tolerancia, mientras que la ausencia de respuesta o una disminución respecto de los valores del control se considera síntoma de sensibilidad (Hernández et al 2000, Toumi et al 2010).
El mantenimiento de las ROS a un nivel compatible con el funcionamiento celular normal se lleva a cabo por diversas actividades enzimáticas con capacidad antioxidante y se clasifican en enzimáticos y no enzimáticos (Foyer et al 2002). Dentro de los antioxidantes enzimáticos se conoce la Catalasa (CAT) que elimina H2O2 (peróxido de hidrógeno) en los peroxisomas, ascorbato peroxidasa (APX) que elimina H2O2 en diversos compartimentos, guaiacol peroxidasa (POX) (Moller & Jensen 2007).La catalasa es una enzima que contiene hierro en su estructura y cataliza la dismutación del H2O2 en agua y oxigeno molecular (Arora et al 2002).
Esta enzima se encuentra en todos los organismos eucariotas aeróbicos y constituye un mecanismo esencial para la eliminación del H2O2 generado en los peroxisomas por oxidasas que participan en la ![]() -oxidación de Acidos grasos, la fotorespiración y el catabolismo de las purinas. La catalasa fue una de las primeras enzimas en ser aislada y obtenida con un alto grado de pureza. Todas las formas enzimáticas de la catalasa son tetraméricas, con pesos moleculares aproximadamente de 220 KDa (Arora et al 2002).
-oxidación de Acidos grasos, la fotorespiración y el catabolismo de las purinas. La catalasa fue una de las primeras enzimas en ser aislada y obtenida con un alto grado de pureza. Todas las formas enzimáticas de la catalasa son tetraméricas, con pesos moleculares aproximadamente de 220 KDa (Arora et al 2002).
El Ascorbato peroxidasa (APX) es una enzima que contiene un grupo hemo y requiere de la presencia de ascorbato en cantidades considerables para su estabilidad y funcionalidad. Reduce el H₂O₂ a agua utilizando el ascorbato como cofactor. La actividad ascorbato peroxidasa ha sido reportada principalmente en cloroplasto y citosol, es un antioxidante cuantitativamente predominante en las células vegetales, se encuentra en todos los compartimentos subcelulares, incluido el apoplasto, en concentraciones que oscilan entre 2-25 mM (Arora et al 2002).
Para realizar la cuantificación de la actividad de Guaiacol Peroxidasa se utiliza como sustratos el guaiacol y el peróxido de hidrogeno en el que se mide la oxidación a una longitud de onda de 470 nm, zona del espectro donde absorbe el guayacol oxidado de coloración carmelita. Se considera la variación de densidad óptica en el tiempo (![]() DO)/(
DO)/(![]() t), durante un tiempo determinado.
t), durante un tiempo determinado.
Por todo lo anteriormente mencionado, el mejoramiento genético de variedades resistentes a sequía es un paso importante para incrementar el rendimiento bajo estrés hídrico (Zhang et al 1999). La transferencia de genes de interés puede ser acelerada mediante la identificación de marcadores moleculares asociados con estos genes de resistencia a sequía. Varios estudios mencionan que el gen/QTL de resistencia sequía está ubicado en el cromosoma 7A (Morgan & Tan 1996, Galiba 2002, Cattivelli et al 2002), y se co-localizo un RFLP sobre el brazo corto a 13 cM de distancia del centrómero. Hasta el día de hoy ya se cuenta con marcadores moleculares microsatélite o SSR que están co-localizados con el gen/QTL de resistencia a sequía en trigo (Zane et al 2002, GrainGenes 2013).
Por los indicado anteriormente y dada la importancia del trigo en la seguridad alimentaria de la población boliviana, la presente investigación tuvo como objetivo estudiar la resistencia a la sequía y validar marcadores moleculares co-localizados con genes/QTLs de resistencia a sequía en 16 variedades y líneas avanzadas de trigo harinero (Triticum aestivum) en dos estadios de desarrollo del cultivo.
Materiales y métodos
Evaluación de la resistencia a estrés hídrico por sequía. La investigación se realizó en la campaña 2012 en uno de los invernaderos de la Fundación PROINPA (zona de El Paso), con T° promedio de 18°C y HR promedio de 50%, ubicada a 15 km de la ciudad de Cochabamba provincia de Quillacollo del departamento de Cochabamba (Bolivia), comprendido entre los paralelos 17° 18’ de latitud Sur y 66° 14’ de longitud Oeste, a una altitud de 2540 msnm.
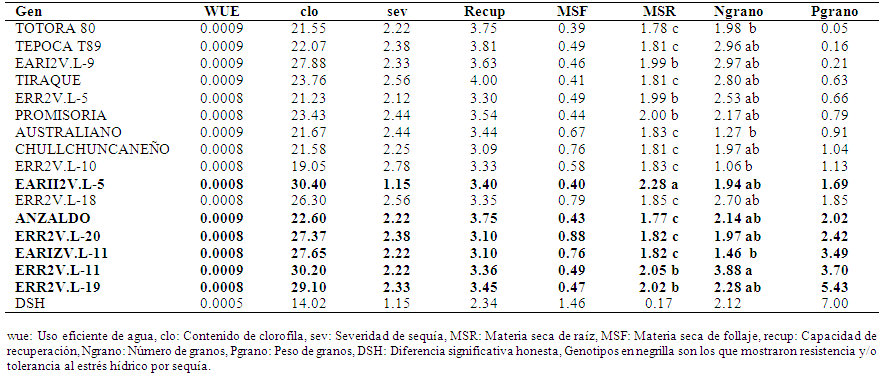
Se utilizó semilla de 16 variedades y líneas avanzadas de trigo del programa de mejoramiento genético de trigo de PROINPA, que fueron sembrados en macetas de 2 kg, (Tabla 1).Para evaluar la resistencia para estrés hídrico por sequía, el experimento se implementó en un diseño de bloques completos al azar con tres repeticiones, se distribuyó en arreglo de parcelas divididas, las parcelas principales fueron: 1 -macetas con sequía, 2 - macetas sin sequía, las sub-parcelas con las 16 variedades y líneas de trigo (Martinez-Garza 1998). Se hicieron dos tratamientos de sequía: 1-en el estadio de macollaje, 2-en el estadio de llenado de grano.
Tabla 1 Variedades y líneas avanzadas de trigo harinero (Triticum estivum) utilizadas para evaluar la resistencia a estrés hídrico por sequía.
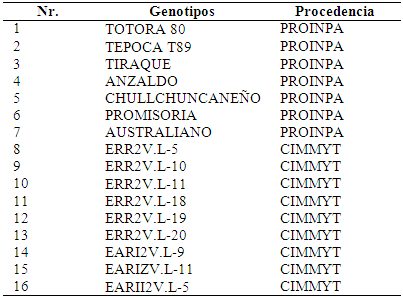
Para el primer estrés hídrico por sequía se suprimió el riego durante 15 días a los 35 días después de la siembra en el estadio de macollamiento. El segundo estrés hídrico por sequía se realizó a los 125 días después de la siembra en el estadio de madurez fisiológica del grano de trigo. Una vez finalizado el periodo de estrés por sequía, se evaluaron parámetros fisiológicos, parámetros morfológicos, mecanismos integrados de resistencia a la sequía y componentes de rendimiento.
Parámetros fisiológicos. Grado de marchites o severidad (S), la capacidad de recuperación (R). La severidad del daño por sequía se evaluó utilizando la escala Blum (1993) adaptada por Angulo et al (2009), se evaluó una vez finalizado el periodo de sequía. También se evaluó el contenido de clorofila con SPAD 502, en las plantas estresadas y no estresadas, según lo recomendado por Gonzales (2009).
Una vez finalizada la evaluación de daño por sequía, se aplicó un riego de recuperación, al cabo de cinco días se evaluó la R, de acuerdo a la escala de severidad de Blum (1993) modificada. La escala comprende valores del 1 al 5, 1 = 0% de recuperación, 5 = 100% de recuperación completa.
Parámetros morfológicos. Se evaluaron fueron materia seca de follaje (MSF) y materia seca de raíz (MSR). Estos parámetros morfológicos se evaluaron al momento de la cosecha. Al final del periodo de estrés y antes del riego de recuperación se evaluó la variable R y/o P (o turgencia), de acuerdo a la escala de Mamani (2000).
Mecanismos de resistencia integrados. Comprendieron la eficiencia de uso de agua (WUE) recomendado por Blum (2005), al inicio y final del estrés hídrico se pesaron las plantas, macetas y sustrato en una balanza de capacidad de 30-0.5 Kg. Marca Ferton Profesional, a fin de calcular la biomasa ganada, después obtener la WUE con relación a la cantidad de agua utilizada durante el estrés, su cálculo se determinó por la formula recomendada por Gabriel et al (2011).
Componentes de rendimiento. La evaluación para determinar el efecto de la sequía aplicada en macollaje y madurez fisiológica sobre el rendimiento, los componentes evaluados fueron número de granos/espiga (Ngrano), peso de grano por espiga (Pgrano). Para evaluar el Ngranos, se contó los granos de cada espiga y se pesó en una balanza de precisión Marca Acculab modelo AL-64 balanza de precisión.
Se realizó un análisis de correlación de Pearson entre S, R, las demás que variables se encontraban correlacionadas y en que magnitud con S y/o R (Martínez-Garza 1988).
Los datos de cada una de las variables de respuesta que se aproximaron a los supuestos de distribución normal y homogeneidad de varianzas, se analizaron de acuerdo a los modelos estadísticos planteados (Martínez-Garza 1988).
Sobre la base del modelo mencionado se realizaron análisis de varianza para probar hipótesis acerca de los efectos fijos y comparaciones de medias mediante contrastes de un grado de libertad para determinar las variedades y líneas avanzadas de trigo más resistentes y/o que se recuperaron. El análisis de varianza también sirvió para estimar los componentes de varianza para los efectos aleatorios. Los análisis indicados se realizaron utilizando el Proc GLM de SAS (SAS 2004).
Análisis bioquímico. La determinación de la actividad enzimática, se utilizaron dos variedades y dos líneas avanzadas de trigo harinero (Anzaldo, Promisoria, ERR2V.L-5 y ERR2V.L-19). La colecta de hojas se realizó después del primer estrés hídrico por sequía en el estadio de macollamiento, se identificó y conservó a -80° C en un freezer Continental 260, hasta su análisis posterior.
Preparación de los Extractos Enzimáticos. El material colectado fue congelado y triturado en nitrógeno líquido (Thermo Scientific modelo 8123), el buffer de extracción enzimático se preparó según la metodología de Frary et al (2010). Se utilizó 0.1 % de polivinilpolipirrolidona (PVPP), 100mM de tampón fosfato de potasio a pH 7.0, 1 mM de EDTA, 1 mM de ácido ascórbico. Los extractos se centrifugaron en una microcentrífuga Legend Micro 21 marca Thermo Scientific, a 10000 rpm por 20 minutos a 4° C, los sobrenadantes se retiraron a otro tubo para las determinaciones: Actividad Catalasa, Ascorbato peroxidasa y Guaiacol peroxidasa las que se realizaron de inmediato.
Actividad Catalasa. La actividad de la enzima CAT, se preparó una mezcla de reacción conteniendo 100 mM tampón fosfato de potasio (pH 6.5), 1.0 mM EDTA, 60.0 mM H₂O₂ (Aebi 1983). La actividad se determinó siguiendo la descomposición de H₂O₂ peróxido de hidrógeno a 240 nm en un espectrofotómetro Marca Biorad.
Actividad Ascorbato Peroxidasa. Se determinó su actividad utilizando el método descrito por Nakano & Asada (1987). La mezcla de ensayo contenía 90 mM de tampón fosfato de potasio (pH 7.0), 0.1 mM EDTA, 0.65 mM ascorbato, 1.0 mM H₂O₂. La actividad se determinó siguiendo la descomposición de H₂O₂ dependiente de ascorbato a 290 nm. Las mediciones se realizaron en un espectrofotómetro marca Biorad.
Actividad Guaiacol Peroxidasa. La determinación de la actividad de esta enzima se preparó una mezcla de reacción conteniendo 100 mM tampón fosfato de potasio (pH 6.5), 1.0 mM EDTA, guaiacol, 50% H₂O₂, (Aebi 1983). La actividad se determinó siguiendo la descomposición de H₂O₂ peróxido de hidrógeno, utilizando guaiacol como agente donador de hidrógeno a 470 nm, en un espectrofotómetro Smart Spec Plus marca Biorad.
Cálculo de la actividad Enzimática. El cálculo de la actividad de las enzimas antioxidantes se utilizó la siguiente fórmula (Aebi 1983): CAT, APX, POX activity (Unit) = (![]() abs/min x reaction volumen)/0.001, realizó la colecta de foliolos jóvenes y tiernos en bolsas plásticas debidamente identificadas con el nombre de cada variedad, luego se almacenó a -20º C, las muestras fueron molidas en morteros de porcelana preenfriados (Teckrik Alemania) y con nitrógeno líquido para obtener un polvo fino y uniforme, se depositó aproximadamente 100 mg en tubos Eppendorf identificados, almacenadas a -20° C, para la extracción de ADN.
abs/min x reaction volumen)/0.001, realizó la colecta de foliolos jóvenes y tiernos en bolsas plásticas debidamente identificadas con el nombre de cada variedad, luego se almacenó a -20º C, las muestras fueron molidas en morteros de porcelana preenfriados (Teckrik Alemania) y con nitrógeno líquido para obtener un polvo fino y uniforme, se depositó aproximadamente 100 mg en tubos Eppendorf identificados, almacenadas a -20° C, para la extracción de ADN.
Selección asistida por marcadores moleculares. El material utilizado, Tabla 1, después de 2 meses se
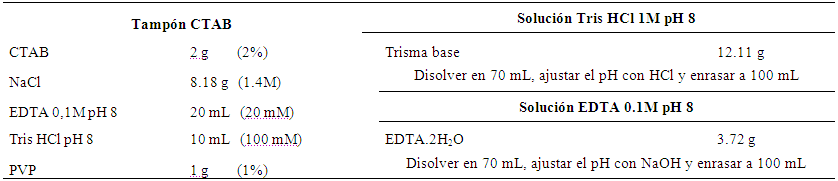
Extracción de ADN. Se realizó mediante el Protocolo CTAB 2X (hexadecil bromuro de trimetil amonio) de Doyle & Doyle (1990) con ligeras modificaciones (Tabla 2).
Tabla 2 Soluciones de trabajo que se utilizan en la Extracción de ADN genómico.
Cuantificación de ADN genómico. El ADN extraído se cuantificó por comparación de la intensidad de las bandas de las muestras, con las bandas del marcador de peso molecular de concentración conocida, para esto se disolvió 1 ![]() L de ADN total en 8
L de ADN total en 8 ![]() L de agua destilada estéril y 1
L de agua destilada estéril y 1 ![]() L de tampón de cargado 6X Amresco Bioexpress con syber green, estas muestras fueron cargadas en geles de agarosa al 1% con buffer de corrida TBE 1X con un marcador de peso molecular de 10000 pb se visualizaron y fotografiaron en un transluminador de luz UV marca Biorad, se comparó las bandas de ADN de las muestras con las del marcador de peso molecular de 10000 pb para determinar la concentración de ADN en ng.
L de tampón de cargado 6X Amresco Bioexpress con syber green, estas muestras fueron cargadas en geles de agarosa al 1% con buffer de corrida TBE 1X con un marcador de peso molecular de 10000 pb se visualizaron y fotografiaron en un transluminador de luz UV marca Biorad, se comparó las bandas de ADN de las muestras con las del marcador de peso molecular de 10000 pb para determinar la concentración de ADN en ng.
Análisis de marcadores. Se han utilizado diferentes marcadores que han sido compendiados en una lista de marcadores potenciales que están ligados y co-localizados con gen/QTLs para sequía (GrainGenes 2013), tabla 3.
Tabla 3 Locus, secuencias, tamaño en pares de bases (pb) y Temperatura de los tres iniciadores microsatélites utilizados en el análisis molecular.
Condiciones de la PCR. 15 ![]() L de mezcla conteniendo 15 ng de ADN molde, 1X de tampón PCR 10X, 0.2 mM dNTP, 1 pmol/
L de mezcla conteniendo 15 ng de ADN molde, 1X de tampón PCR 10X, 0.2 mM dNTP, 1 pmol/![]() L de cada iniciador y 0.025 U/
L de cada iniciador y 0.025 U/![]() L de la enzima Taq polimeraza fue utilizado. El programa de amplificación fue realizado en un termociclador en gradiente (T100 Thermal Cycler marca BioRad), utilizando un rango de 8 temperaturas de 50° C a 60° C definidas automáticamente, que consistió en un tiempo de desnaturalización inicial de 5 min del ADN a 94º C, 35 ciclos a 94º C por 1 min, temperatura de anillamiento 60, 59 y 55º C por 45 s, y 72º C por 1 min, 1 ciclo a 72° C por 5 min y una extensión final de 1 ciclo a 15° C por 1 min, según el marcador. Subsecuentemente, los productos de amplificación fueron visualizados y cuantificados mediante electroforesis en geles de agarosa al 1.8%, aplicándose 5V/cm.
L de la enzima Taq polimeraza fue utilizado. El programa de amplificación fue realizado en un termociclador en gradiente (T100 Thermal Cycler marca BioRad), utilizando un rango de 8 temperaturas de 50° C a 60° C definidas automáticamente, que consistió en un tiempo de desnaturalización inicial de 5 min del ADN a 94º C, 35 ciclos a 94º C por 1 min, temperatura de anillamiento 60, 59 y 55º C por 45 s, y 72º C por 1 min, 1 ciclo a 72° C por 5 min y una extensión final de 1 ciclo a 15° C por 1 min, según el marcador. Subsecuentemente, los productos de amplificación fueron visualizados y cuantificados mediante electroforesis en geles de agarosa al 1.8%, aplicándose 5V/cm.
Resultados
El análisis de varianza de las variables de respuesta al estrés hídrico por sequía en trigo (Tabla 4) mostró solamente diferencias notables al 1% de probabilidad (Pr<0.01) para Número de granos y Peso de grano, con un CV de 16.24 y 38.59 respectivamente. Las demás variables no fueron notables. Esto está indicando que los genotipos (tratamientos) tuvieron una respuesta diferenciada a la sequía que fue expresada básicamente en el Número (Ngrano) y Peso de granos (Pgrano).
Tabla 4 Análisis de varianza de variables de respuesta al estrés hídrico por sequía en trigo, El Paso, Cochabamba, 2012.
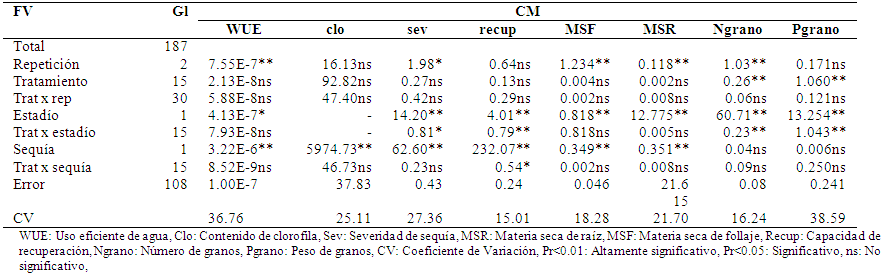
Respecto al estadio de sometimiento de sequía, es decir en el macollaje y en la madurez fisiológica, todas las variables evaluadas mostraron diferencias visibles al 1% y 5% de probabilidad, con un CV 15.01 a 38.59. Esto indica que hubo un efecto importante y diferente entre la sequía sometida en macollaje y la sequía sometida en madurez fisiológica, el efecto indicado se confirmó en la interacción entre tratamiento por estadio, donde las variables Severidad (sev), Recuperación (recup), Número de granos (Ngrano) y Peso de granos (Pgrano), mostraron diferencia visibles al 5% y 1% respectivamente.
También se observó que el efecto de la sequía fue notoria al 1% de probabilidad para las variables cantidad de clorofila (clo), sev, recup, materia seca de follaje (MSF) y materia seca de raíz (MSR), lo que indicó que estas variables son las que mostraron una respuesta al estrés hídrico por sequía. No hubo diferencias notables para Ngrano y Pgrano, esto está denotando que aparentemente las plantas estresadas y no estresadas no mostraron diferencias en la producción, esto probablemente debido al sustrato utilizado, dado que no se observaron diferencias notorias en la madurez de las plantas. Sin embargo, se observó diferencias notorias (Pr<0.05) de respuesta en la capacidad de recuperación (recup) de las plantas estresadas de los genotipos resistentes.
Una comparación de medias mediante la prueba de tukey de los genotipos de trigo para estadío (macollaje y madurez fisiológica) (Tabla 5) confirmó que hubo diferencias significativas para las variables de sev, recup, MSF, MSR, Ngrano y Pgrano, no así para eficiencia de uso de agua (wue). Los datos de la Tabla 5 indican además, que el estadio de madurez fisiológica es el que mayor efecto por estrés hídrico por sequía fue para las variables wue, sev y Pgrano, en cambio en el estadio de macollamiento las variables más afectadas fueron la recup, MSF y Ngrano.
Tabla 5 Comparación de medias mediante la prueba de Tukey en variedades y líneas avanzadas de trigo para estadio y sequía con cada variable evaluada, El Paso, Cochabamba, 2012.
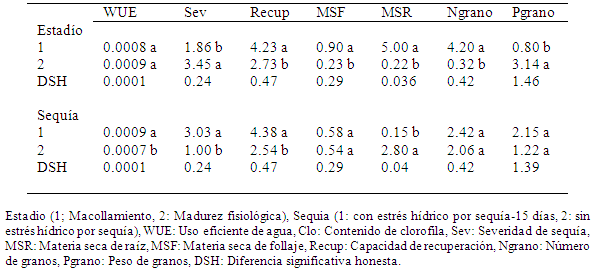
Para el caso de sequía (con y sin estrés hídrico por sequía) se observó (Tabla 5) diferencias palpables para wue, sev, recup y MSR; y no así para MSF, Ngrano y Pgrano. Esto, como se dijo antes, probablemente es a causa de que no se observó diferencias marcadas entre plantas estresada y no estresadas en el momento de la cosecha, debido a que el sustrato utilizado no ayudó a que las plantas no estresadas (testigos) tuvieran mejor vigor y desarrollo. Está claro por los resultados de la Tabla 5 que en general todas las variables evaluadas tuvieron mayor efecto en el tratamiento con sequía, con excepción de la MSR, que un producto de la acumulación de fotosintatos en la raíz.
La comparación de medias mediante la prueba de tukey (Tabla 6) mostró que los genotipos evaluados no mostraron diferencias visibles para wue, sev, clo, recup, MSF y Pgrano, pero si se observó diferencias notables entre genotipos para las variables MSR y Ngrano, se observó que los genotipos Anzaldo, ERR2V.L-20, EARII2V.L-5, EARIZV.L-11, ERR2V.L-11 y EE2V.L-19 fueron los más resistentes y/o tolerantes al estrés hídrico por sequía.
Tabla 6 Comparación de medias mediante la prueba de Tukey en variedades y líneas avanzadas de trigo, El Paso, Cochabamba, 2012.
La correlación de Pearson realizada entre todas las variables evaluadas (Tabla 7) mostró una correlación negativa alta y significativa para clo y sev, esto indicó que a mayor severidad o marchitez (sev) hay menor contenido de clorofila (clo). En cambio, se observó una correlación moderada y significativa entre las variables clo y MSR, indicando esto que el contenido de materia seca de la raíz (MSR) depende del contenido de clorofila, que se refleja en la acumulación de fotosintatos.
Tabla 7 Correlaciones de Pearson para las variables de respuesta evaluadas en trigo, 2012.
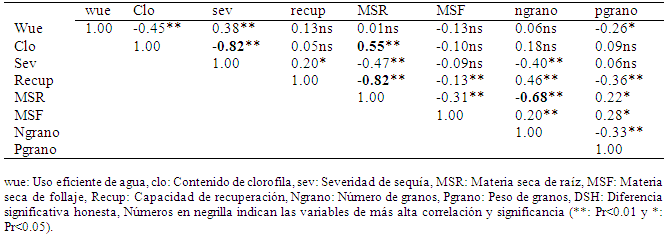
También se observó una correlación alta negativa y significativa entre la recup y la MSR, esto indicó que los genotipos con mayor capacidad de recuperación tienen menor acumulación de MSR, lo cual es comprensible, porque la planta gasta mucho de la energía en recuperarse del estrés en detrimento de la acumulación de fotosintatos en la raíz. Finalmente se encontró una alta correlación negativa entre la MSR y el Ngrano, que está indicando que los genotipos que con alta MSR producen menor número de granos, este efecto no fue fácil de explicar, pero probablemente las plantas estresadas acumulan fotosintatos para sobrevivir y no necesariamente para producir granos.
Todas las demás variables mostraron correlaciones bajas y no significativas.
Evaluaciones Bioquímicas a estrés hídrico por sequía. Está descrito que en situaciones de estrés abiótico la planta incrementa su velocidad de producción de oxidantes o da una disminución en la actividad de la defensa antioxidantes, terminando en el aumento de las concentraciones de las ROS (Mano 2002). Sin embargo, la actividad antioxidante no solo se interpreta como el proceso de atrapar ROS, como un mecanismo que evita la formación de estas especies reactivas de oxígeno, junto a procesos de reparación y eliminación de los productos de oxidación, por lo que resulta importante evaluar de todos en conjunto.
La evaluación de la actividad enzimática (enzimas antioxidantes) indica la capacidad de detoxificar el H₂O₂, en las plantas, entonces si la actividad enzimática es elevada respecto al control, indicaría que la planta tiene capacidad de eliminar especies reactivas de oxígeno - ROS y considerarla tolerante (Mano 2002).
En la Tabla 8, se observa tres de las cuatro variedades de trigo sometidas a estrés hídrico por sequía, que mostraron una actividad enzimática CAT elevada respecto de su control. Anzaldo, Promisoria y ERR2V.L-5 tuvieron valores de 217.98, 152.04 y 274.59 U/g, mientras que el valor más bajo para la actividad catalasa se encontró en ERR2V.L-19 con una actividad enzimática de 278.46 U/g.
Tabla 8 Actividad de Enzimas antioxidantes: Catalasa (CAT), Ascorbato Peroxidasa (AsPx) y Guaiacol Peroxidasa (POX).
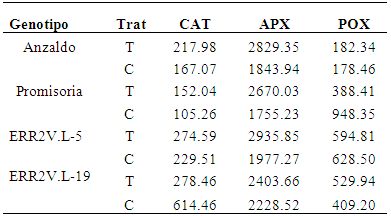
Cuando se analizó la actividad específica del APX en los extractos de hojas (Tabla 8), se observó un incremento respecto del control para todas las variedades evaluadas con valores que estuvieron entre 2403.66 a 2935.85 U/g, que correspondieron a las líneas ERR2V.L-19 y a ERR2V.L-5 respectivamente.
Resultados diferentes fueron observados para la actividad específica POX para las variedades Promisoria y ERR2V.L-5 con valores bajos, 388.41 y 594.81 U/g respectivamente.
APX y POX desempeñan función catalítica importante al reducir los niveles de H₂O₂, peróxido de hidrógeno en tejidos vegetales (Asaka 1992), evitando que este compuesto se difunda en las membranas biológicas para reaccionar con iones libres como el Fe2+ y generar el potente radical hidroxilo, a través de la reacción de Fenton (Vogiatzi et al 2009).
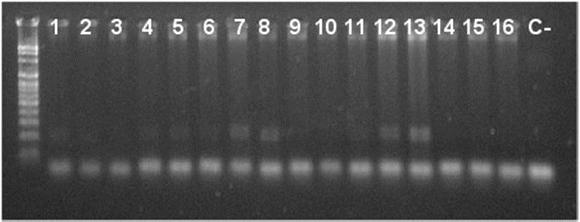
Figura 1 Gel de agarosa al 1.8 % que muestra la migración del fragmento microsatélite Xwmc 596. Leyendas de al 16. 1. Totora 80, 2.Tepoca T89, 3.Tiraque, 4.Anzaldo, 5.Chullchullcaneño, 6.Promisoria, 7.Australiano, 8.ERR2V.L-5, 9.ERR2V.L-10, 10. ERR2V.L-11, 11. ERR2V.L-18, 12. ERR2V.L-19, 13. ERR2V.L-20, 14. EARI2V.L-9, 15.EARI2V.L-11, 16. EARI2V.L-5, C-.Control Negativo.
Se observó (Tabla 8) que la variedad Anzaldo mostró una actividad enzimática incrementada respecto de los controles en todos los casos (CAT-APX y POX), siendo por lo tanto la más resistentes al estrés hídrico por sequía.
Se realizó una correlación entre las variables bioquímicas y las variables fisiológicas, morfológicas y componentes de rendimiento. Aun cuando algunos de las correlaciones mostraron ser altas, ninguna fue significativa. Indicando esto que no hubo asociaciones importantes entre todas las variables analizadas. Validación de marcadores moleculares. De los tres iniciadores (primers) utilizados, uno no amplificó (Xwmc603), para los iniciadores, Xwmc596 y Xwmc9 se determinó una temperatura de anillamiento de 55º C y 59º C respectivamente.
Figura 2 Gel de agarosa al 1.8 % que muestra la migración del fragmento microsatélite Xwmc 9. Leyendas de al 16. 1. Totora 80, 2.Tepoca T89, 3.Tiraque, 4.Anzaldo, 5.Chullchullcaneño, 6.Promisoria, 7.Australiano, 8.ERR2V.L-5, 9.ERR2V.L-10, 10. ERR2V.L-11, 11. ERR2V.L-18, 12. ERR2V.L-19, 13. ERR2V.L-20, 14. EARI2V.L-9, 15.EARI2V.L-11, 16. EARI2V.L-5, C-.Control Negativo.
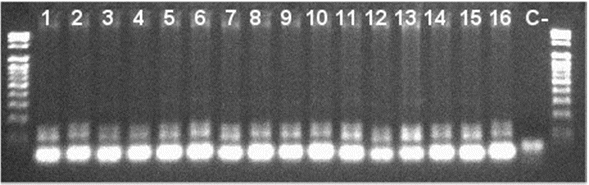
El marcador Xwmc603 no mostró amplificación en la región microsatélite, debido probablemente a que las variedades analizadas no tienen la región conservada para este iniciador (Li et al 2002).
Los productos PCR para los marcadores Xwmc596 y Xwmc9 mostraron polimorfismo en las variedades y líneas avanzadas de trigo harinero (Triticum aestivum) (Figuras 1 y 2), resultado que concuerda con los reportado por Zhang (1999). Las variedades Tiraque, Anzaldo y ERR2V.L-19 (Figura 2) mostraron un polimorfismo diferente al resto al aplicar con el marcador SSR Xwmc9, que según Galiba (2002), el gen de resistencia a sequía no solo se encuentra en el cromosoma 7A, sino también en el cromosoma 1A. Lo cual explica la diferencia observada.
No todas las variedades evaluadas con el marcador Xwmc 596 muestran un perfil similar para resistencia a sequía. Los marcadores SSR (Xwmc596 y Xwmc9) están co-localizados con el gen/QTL de resistencia a sequía y pueden ser utilizados para la selección asistida por marcadores moleculares.
Discusión
Está claro que un estrés hídrico por sequía causa pérdidas en cualquier estadio del desarrollo de la planta, sin embargo, en nuestro trabajo hemos observado que el efecto más drástico está en el estado fisiológico del llenado de grano, que definitivamente no permite la producción de grano. Estos resultados concuerdan con lo encontrado por otros investigadores (O’Toole & Moya 1981, Morgan 2004, Frahm et al 2004, Machado 2004, Mukandama 2005, Herbas & Montaño 2011). También es notorio el hecho que, en este tipo de resistencia intervienen probablemente muchos genes (poligénico) que trabajan en diferentes estadios de la planta (Asfaw 2011), lo que puede observare en la presente investigación, la respuesta de las variables evaluadas no fueron las mismas en los dos estadios.
Pudimos observar diferencias marcadas para wue, sev, recup y MSR entre plantas estresadas y no estresadas, pero no para MSF, Ngrano y Pgrano. Esto, probablemente se deba a que no se observó diferencias marcadas entre plantas estresada y no estresadas en el momento de la cosecha, debido a que el sustrato utilizado no ayudó a que las plantas no estresadas (testigos) tuvieran mejor vigor y desarrollo.
Por otra parte las evaluaciones bioquímicas nos han permitido determinar que el CAT, APX y POX mostraron una actividad enzimática elevada permitiendo ser útiles para la selección de líneas resistentes al estrés hídrico por sequía, esto indica que la función catalítica de reducción del peróxido de hidrógeno en los tejidos vegetales, tal como lo indica Asaka (1992). Sin embargo, debemos mencionar, que si bien algunas correlaciones fueron altas, no fueron significativas entre las variables bioquímicas, fisiológicas, morfológicas y los componentes de rendimiento. Esto no significa que no sean útiles, sino que está confirmando el hecho de que hay una complejidad importante en la resistencia al estrés hídrico por sequía, de hecho están involucrados muchos genes de efectos aditivos (herencia poligénica de baja heredabilidad), tal como lo menciona Asfaw (2011).
Finalmente pudimos observar que los productos PCR, que los marcadores Xwmc596 y Xwmc9 mostraron polimorfismo en las variedades y líneas avanzadas de trigo harinero (Triticum aestivum), y se co-localizan con el gen/QTL para resistencia a sequía en el cromosoma 7A, resultado que concuerda con los reportado por Zhang (1999), esto es de suma importancia para la utilización de estas técnicas modernas en la selección asistida por marcadores moleculares (SAMM).
Conflictos de intereses
Esta investigación recibió financiamiento total del Proyecto “Fortaleciendo capacidades de innovación participativa para luchar contra la pobreza rural” (IP–Holanda) y no presenta conflictos de interés.
Agradecimientos
Se agradece al apoyo económico del proyecto: “Fortaleciendo capacidades de innovación participativa para luchar contra la pobreza rural” (IP – Holanda). Gracias al M. Sc. Mario Crespo de la Fundación PROINPA, por la revisión y sugerencias al presente documento.
Literatura citada
Aebi H. Catalase in vitro. Meth Enz. 1983; (105):121-126. [ Links ]
Angulo A, Siles M, Ríos R, Gabriel J. Caracterización de 118 accesiones de arveja (Pisum sativum L,) del Banco de Germoplasma del Centro de Investigaciones Fitoecogenéticas de Pirumani para resistencia a sequía. Revista de Agricultura, Bolivia. 2009; 42(60):25-31. [ Links ]
Arora A, Sairam RK, Srivatava GC. Oxidative stress and antioxidative system in plants. Current Science. 2002; 82:1227-1238. [ Links ]
Asada K. Ascorbate peroxidase - a hydrogen peroxide scaveng-ingenzyme in plants. Physiol Plant. 1992; 85:235-241. [ Links ]
Asfaw A. Breeding for drought tolerance by integrative design: The case of common bean (Phaseolus vulgaris L.) in Ethiopia. Thesis Wageningen University, Wageningen, NL. 2011; 185 pp. [ Links ]
Asociación de Productores de Oleaginosas y Trigo-ANAPO. ANAPO Noticias No, 82, El desafío de producir más, Santa Cruz, Bolivia, Disponible en: http://www,elsol,com,bo. 2012; (revisado el 29 de mayo de 2012). [ Links ]
Blum A. Drought resistance, water-use efficiency, and yield potential—are they compatible, dissonant, or mutually exclusive? Austr J Agri Res. 2005; 56:1159-1168.
Blum A. Selection for sustained production in water deficit environments, Crop Sci. 1993; I: 343-347. [ Links ]
Boyer J, Westgate M. Grain yields with limited water. J Exp Bot. 2004; 55:2385-2394. [ Links ]
Cattivelli L, Baldi P, Crosatti C, Di Fonzo, Faccioli NP, Grossi M, et al. Chromosome regions and stress-related sequences involved in resistance to abiotic stress in Triticeae. Plant Mol Biol. 2002; 48(5-6):649-665. [ Links ]
Doyle JJ, Doyle JL A rapid total DNA preparation procedure for fresh plant tissue. Focus. 1990; 12:13-15. [ Links ]
Foyer CH. Oxidant and antioxidant dignalling in plants: a reevaluation of the concept of oxidative stree in a physiological contex. Plant Cell Envir. 2002; 28:1056-1071. [ Links ]
Frahm M. Breading beans for resistance to terminal drought in the lowland tropics. Euphytica. 2004; 136(2):223-232. [ Links ]
Frary A, Göli D, Keles D. Salt tolerance in Solanum pennellii: antioxidant response and related QTL. Mol Biol.2010; 10(1):58. [ Links ]
Gabriel J, Porco P, Angulo A, Magne J, La Torre J, Mamani P. Resistencia genética a estrés hídrico en variedades de papa (Solanum tuberosum L,) bajo invernadero. Revista Latinoamericana de la Papa. 2011; 16(2):173-208. [ Links ]
Galiba G. Mapping of genes regulating abioticstress tolerance in cereals. Acta Agron Hungarica.2002;50(3):235-247. [ Links ]
Gallagher J, Biscoe P, Hunter B. Effects of drought on grain growth. Nature.1976; 264:541-542. [ Links ]
Gonzáles A. Aplicación del medidor portátil de clorofila En programas de mejora de trigo y cebada. Agroecología. 2009; 4:111-116. [ Links ]
GrainGenes: A Database for Triticiae and Avena [homepage on the Internet]. Whitehouse: Agricultural Research Service of the US Department of Agriculture. 2013 [actualizacion 2012 Mayo 31; cited 2012 Jun 30]. Disponible en http://wheat.pw.usda.gov/GG2/index.shtml. [ Links ]
Herbas J, Montaño I. Periodos de estrés hídrico en trigo harinero (Triticum aestivum L,) variedad Totora 80.Revista de Agricultura, Bolivia. 2011; 50:1-8. [ Links ]
Hernández JA, Jimenez A. Tolerance of pea (Pisum sativum L) to long term salt stress is associated with induction of antioxidant deferences. Plant Cell Envir. 2000; 23:853-862. [ Links ]
Instituto Nacional de Estadística (INE). Estadísticas Agropecuarias. La Paz, Bolivia. 2008. [ Links ]
Kramer P. Drought, stress, and the origin of adaptations, Page 7-20 in N, Turner and P, Kramer (eds,), Adaptation of plants to water and high temperature stress. John Wiley & Sons, New York, USA. 1980. [ Links ]
Li Y Chu, Korol AB, Fahima T, Nevo E. Microsatellites Within Genes: Structure, Function, and Evolution. Mol Bio Evol. 2002; 21(6):991-1007. [ Links ]
Machado A. Estrés hídrico, mecanismos de resistencia y tolerancia a la sequía. Universidad de Granma, Cuba. 2004. [ Links ]
Mamani P. Effet de la secheresse sur six varietés de ponme de terre dans les andes boliviennes. Tesis M.Sc., Universite Catholique de Louvain Faculte des Sciences Agronomiques, Bélgica. 2000; 43 pp. [ Links ]
Mano J. Early events in enrionmental stresses in plants-induction mechanisms of oxidative stress. Page 217-246 in Oxidative Stress in Plants, UK. 2002. [ Links ]
Martínez-Garza A. Diseños experimentales: Métodos y elementos de teoría. Editorial Trillas, México D.F. México. 1988; 756 pp. [ Links ]
Ministerio de Agricultura, Ganadería & Desarrollo Rural - MAGDR. Inventario Nacional de Sistemas de Riego, Dirección General de Suelos y Riego (DGRS) – Programa Nacional de Riego (PRONAR). La Paz, Bolivia. 2000.
Moller IM, Jensen. Oxidative modifications to cellular components in plants. Ann Rev Plant Biol. 2007; 58:459-481. [ Links ]
Morgan JM. Osmoregulation as a selection criterion for drought tolerance in wheat, Austr J Agri Res. 2004; 34(6):607-614. [ Links ]
Morgan JM, Tan MK. Chromosomal location of a wheat osmoregulation gene using RFLPanalysis. Austr J Plant Physiol.1996;23(6):803-806. [ Links ]
Mukandama JP, González MC, Suárez L, Alvarez Y. Incremento de alto potencial productivo en tomate (Lycopersicon esculentum Mill) por rayos gamma 60Co en condiciones de sequía agrícola. Alimentario. Revista de Tecnología e Higiene de los Alimentos. 2003; 340: 95-98. [ Links ]
Nakano Y, Asada K. Purification of ascorbate peroxidase in spinach chloroplasts, its inactivation in Ascorbate-depleted medium and reactivation by monodehydroascorbate radical. Plant cell physiol. 1987; 28(1):131-140. [ Links ]
O’Toole J, Moya T. Water deficits and yield in upland rice. Field Crops Res. 1981; 4:247-259. [ Links ]
Organización de las Naciones Unidas para la Alimentación y la Agricultura (ONU-FAO). Perspectivas de cosechas y situación alimentaria. Disponible en: http://www,fao,org (revisado el 29 de mayo de 2012). 2011. [ Links ]
SAS, Institute Inc. User's guide, Statistics, Version 9,2. SAS Institute Inc. Cary, North Carolina, USA. 2004. [ Links ]
Somers DJ. A high-density wheat microsatellite consensus map for bread wheat (Triticum aestivum L.). Theo Appl Gen. 2004; 109:1105-1114. [ Links ]
Toumi I, Moschou PN, Paschalidis KA. Abcisic acid signals reorientation of polyamine metabolism to orchestrate stress responses via the polyamine exodus pathway in grapevine. J Plant Physiol. 2010; 167:519-525. [ Links ]
Vogiatzi G, Dimitris D, Christodoulus S. The role of oxidative stress in atherosclerosis hellenic. J Cardiol. 2009; 50:402 [ Links ]
Zane L, Bargellon L, Paternello T. Strategies for microsatellite insolation. Mol Ecology. 2002; 11: 1-16. [ Links ]
Zhang J, Nguien HT, Blum A. Genetic analysis of osmotic adjustement in crop plants. J Exp Bot.1999; 50(332):291-302. [ Links ]

