










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista Científica Ciencia Médica
Print version ISSN 1817-7433On-line version ISSN 2220-2234
Rev Cient Cienc Méd vol.23 no.2 Cochabamba 2020
ARTÍCULO DE REVISIÓN
ESTUDIO ANATÓMICO DEL HIPOCAMPO COMO UNA DE LAS REGIONES DE NEUROGÉNESIS MÁS RELEVANTE
ANATOMICAL STUDY OF THE HIPPOCAMPUS AS ONE OF THE MOST RELEVANT NEUROGENESIS REGIONS
Msc. Vladimir Camacho Ugarte1![]() , Dra. Agatha Bastida Codina PhD2
, Dra. Agatha Bastida Codina PhD2![]() , Dra. Sandra Giovana Zarate PhD3
, Dra. Sandra Giovana Zarate PhD3![]()
1 Doctorante en Ciencias Biomédicas. Docente Anatomía Humana Facultad de Ciencias Tecnológicas de la Salud. Universidad Mayor Real y Pontífice de San Francisco Xavier de Chuquisaca.
2 Científico Titular. Instituto de Química Orgánica General, IQOG, CSIC. 28006-Madrid-España.
3Universidad Mayor Real y Pontífice de San Francisco Xavier de Chuquisaca. Facultad de Tecnología. Carrera de Ingeniería Química. Sucre-Bolivia.
Correspondencia a: Vladimir Camacho Ugarte
Correo electrónico: vladicamachougartr@gmail.com
Celular: +(591) 7286202 - 64301572
ORCID: https://orcid.org/0000-0001-9447-3877
https://orcid.org/0000-0002-5141-9595
https://orcid.org/0000-0003-2501-388X
Procedencia y arbitraje: no comisionado, sometido a arbitraje externo.
Recibido para publicación: 24 de octubre de 2019
Aceptado para publicación: 24 de diciembre de 2020
Citar como: Camacho Ugarte V, Bastida Codina A, Giovana Zarate S. Estudio anatómico del hipocampo como una de las regiones de neurogénesis más relevante. Rev Cient Cienc Med 2020; 23(2): 221-230
RESUMEN
Los avances tecnológicos en el campo de la medicina regenerativa han abierto un sin fin de posibilidades terapéuticas, que hace algún tiempo hubiera sido imposible de concebir. Dichos avances se han enfocado en el estudio regenerativo de células propias del Sistema Nervioso Central (SNC), gracias a la identificación plena de las funciones de las Células Madre Neurales (CMNs) y de ciertos marcadores neurogliales que intervienen en la formación de células troncales neuronales, originando no solo astrocitos, sino también neuronas, oligodendrocitos y células ependimarias.Teniendo en cuenta estos aspectos, la presente revisión bibliográfica se centra en el estudio anatómico de la zona del hipocampo como una de las regiones génicas más importantes e interesantes. Su papel fundamental es la formación de "nichos neurogénicos", lugar donde se producen los procesos de diferenciación y formación de nuevas neuronas a partir de células troncales presentes durante toda la vida.
Palabras clave: Hipocampo, células madre, neurogénesis.
ABSTRACT
Technological advances in the field of regenerative medicine have opened an endless of therapeutic possibilities, that some time ago would have been impossible even to conceive. These developments have focused their attention so important in the study of regenerative cells of the Central Nervous System (CNS), thanks to the full identification of the functions of neural stem cells (NSCs) and some of theglial markers involved in the formation of neural stem cells, causing not only astrocytes, but also neurons, oligodendrocytes, and ependymal cells. Having this in mind, the present review focuses its attention on the anatomical study of the area of the hippocampus as one of the most important and interesting gene regions, from its fundamental role in the formation of neurogenic niches, place where the processes of differentiation and formation of new neurons from neural stem cells present throughout life.
Keywords: Hippocampus, stem cells, neurogenesis.
INTRODUCCIÓN
El cuerpo humano, está compuesto por una gran cantidad de elementos orgánicos que interactúan dentro de un orden funcional conjunto. Uno de estos elementos y tal vez el más complejo es el Sistema Nervioso Central (SNC) que tiene la capacidad de percibir y responder a estímulos extrínsecos e intrínsecos, además está encargado de integrar el funcionamiento de los sistemas que componen a un organismo, así como de coordinar y controlar las interacciones con su medio1.
Las vías del Sistema Nervioso (SN) forman circuitos complejos que tienen la capacidad de modificar respuestas en su estructura y funciones (lesiones, presiones ambientales, alteraciones en el estado interno, etc.). Esta capacidad, se llama plasticidad cerebral o neuronal y es responsable de la flexibilidad adaptativa del SN. Dicha flexibilidad depende de su integridad orgánica/química y permite adaptar sistemas de prevención y protección mecánica, incluyendo su reparación (formación de nuevas células nerviosas)2.
Las regiones del cerebro capaces de formar estas nuevas células nerviosas mediante dichos procesos de reparación se conocen como "nichos neurogénicos" (hipocampo y bulbo olfatorio). En el cerebro humano existen dos regiones anatómicas: la zona subventricular con sus ventrículos laterales (SVZ) y la zona subgranular (SGZ) del giro dentado (GD) del hipocampo (Fig.1), con una alta presencia de células madre y precursores químicos específicos, tales como la GFAP (Proteína glíal fibrilar ácida) y la NCAM (Molécula de adhesión celular neural)3. En especies animales, estas dos regiones generan nuevas células nerviosas permanentemente, aunque la cantidad de producción varía según la edad y la especie4. También se han detectado la presencia de células madre y precursores neurales en otras regiones del cerebro adulto tales como: corteza cerebral, hipotálamo, locus Niger, amígdala y corteza piriforme . La mayor parte de estas investigaciones mantienen la idea de que las regiones "no neurogénicas" producen únicamente células gliales, debido a que carecen del microambiente necesario para la formación de neuronas, mientras que la formación de nuevas neuronas en estas regiones se produciría sólo tras un estímulo externo drástico11,12.
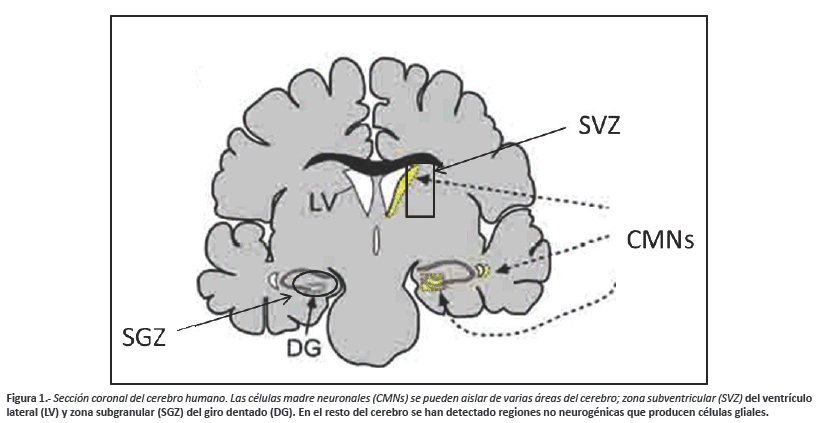
Por otro lado, la capacidad neurogénica en otras regiones que no sean las incluidas en la zona subventricular (SVZ) y subgranular (SGZ), queda hasta ahora poco establecida, generando controversias de amplio análisis. En la presente revisión se exponen los últimos avances y teorías respecto a la anatomía del circuito hipocampal en el proceso de integración neuronal en la neurogénesis del cerebro adulto.
METODOLOGÍA DE BÚSQUEDA DE ARTÍCULOS, NIVELES DE EVIDENCIA
Los métodos empleados en este artículo se basaron en la búsqueda automatizada en bases de datos de los últimos años; MEDLiNE, EMBASE, PubMed, y Current Contents. Las palabras claves que se usaron fueron: Células neuronales, neurogénesis, glía, hipocampo, sistema nervioso central entre otras. También se realizó la búsqueda por autores relevantes en el campo de investigación tratado en este artículo, entre otros muchos cabe destacar; Rodríguez-Moreno A (86 artículos encontrados), Haladay K. (33 entradas encontradas en PubMed) y Noctor S.C (47 artículos). Para información más detallada se consultó libros del campo de investigación propuesto en este artículo tales como: "The Hippocampus BooK", autores: Per Andersen, et al. 2007 Oxford University Press, "The Hippocampus: Neurotransmision & Plasticity in the Nervous System" autor: Philippe Taupin, Nova Biomedical 2007, entre otros libros consultados. El método de selección de los artículos presentados en esta revisión fue teniendo en cuenta autores relevantes del campo y publicaciones recientes del mismo.
DESARROLLO
Antecedentes Bibliográficos
A principios del siglo XX se pensaba que el cerebro era un órgano altamente sofisticado pero condenado a un deterioro inevitable por el paso del tiempo no generando nuevas neuronas después del nacimiento, por lo tanto, se afirmaba una incompatibilidad con la generación e integración de nuevas células nerviosas durante la etapa adulta13.
Es en la década de los años '60 donde se evidencia por primera vez, la generación de nuevas neuronas en el cerebro de roedores adultos (génesis de nuevas neuronas, específicamente en el GD y en el bulbo olfatorio14,15. En la década de los '80 se logra demostrar la funcionalidad de nuevas neuronas en aves adultas, procesando señales aferentes en respuesta a estímulos sonoros16,17. En la década de los '90 se demuestra que los axones de las nuevas neuronas son capaces de establecer sinapsis con otras neuronas, empezando así los pasos a la aceptación de la neurogénesis adulta 18. Sin embargo, otros estudios negaban la existencia de neurogénesis adulta en primates prevaleciendo el antiguo dogma en mamíferos, considerando que la neurogénesis adulta en aves era solo una forma de especialización evolutiva19.
Los avances científicos y tecnológicos en el campo de la inmunohistoquímica, la citología y las técnicas moleculares abrieron y plantearon un nuevo paradigma dentro de las Neurociencias, ya que por fin se lograban demostrar repetidamente sucesos neurogénicos en zonas específicas del cerebro de distintos animales, incluido el del ser humano, siendo estas zonas principalmente el hipocampo, giro dentado y el bulbo olfatorio20,21. La neurogénesis de estas áreas, comparten algunas características con la neurogénesis embrionaria y posnatal, entre ellas la proliferación de Células Progenitoras Neuronales (CPN) que migraban, maduraban y formaban conexiones sinápticas. Estas nuevas experiencias lograron identificar áreas fuera de estos nichos neurogénicos, llamándoles zonas 'no neurogénicas, aunque en ellas las CPN proliferan y contribuyen a la gliogénesis (generación de células gliales), pero sólo en condiciones patológicas. Es así que se demostró en estudios in vitro que estas CPN son capaces de producir neuronas y glía, dicho fenómeno sugiere que existen CPN con potencial neurogénico en otras áreas del SNC adulto22.
Este nuevo paradigma neuronal ha cambiado sustancialmente las ideas de salud y enfermedad en el SNC y por otro lado se han modificado las ideas dentro de los procesos de la neurogénesis, no solo como la generación de nuevas neuronas sino también como un fenómeno que produce sus propias interrelaciones y vínculos funcionales, partiendo de elementos anatómicos y precursores químicos como dianas y proteínas que han sido identificadas en zonas especializadas denominadas 'nichos neurogénicos, presentes fundamentalmente en SVZdel VL y la SGZ del GD del hipocampo. Ya se ha demostrado que una población de CPN persisten a lo largo de toda la vida y son capaces de lograr su diferenciación para integrarse a redes neuronales existentes a través de un proceso denominado 'neurogénesis en el cerebro adulto23. La neurogénesis es el proceso por el cual se generan nuevas células en el SNC y es esta capacidad de auto regeneración del cerebro adulto, que ha despertado un gran interés por las potenciales aplicaciones terapéuticas en diversas lesiones neurológicas. A partir de todos estos hallazgos, surge la evidencia para determinar qué nuevas neuronas se continúan generando en el cerebro adulto, a partir de células primarias denominadas "células madre" (CM), con propiedades de diferenciación y capaces de producir mecanismos que garantizan la homeostasis de todo el sistema nervioso24(Figura 2).
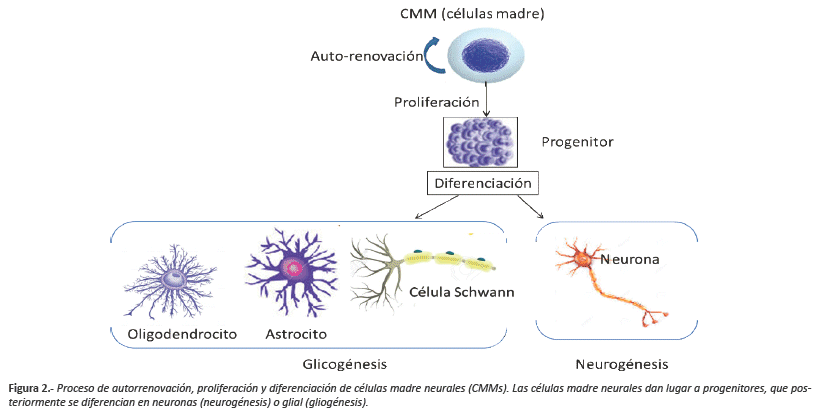
Autorenovación y diferenciación de células madre
Las células madre mesenquimales (CMMs) son células no especializadas con capacidad de autorrenovarse. Este tipo de células pueden permanecer como célula madre (división simétrica), o convertirse en cualquier otro tipo celular con nuevas funciones especializadas (división asimétrica). La localización física de dichas células es en los "nichos neurogénicos" en los cuales se encuentran astrocitos, microglía, componentes extracelulares y vasos sanguíneos esenciales para que se realicen los procesos de autorrenovación y diferenciación25. Este conjunto de elementos genera las interacciones célula-célula, a través de precursores y moléculas presentes en la matriz extracelular, incluyendo sustancias difusibles que juegan un papel importante en el fenómeno neurogénico adulto pudiendo modular la proliferación, autorenovación, potencialidad, diferenciación, supervivencia y migración celular26.
Estas células están presentes en toda la vida de un individuo y su auto-renovación en el nicho neurogénico depende de las condiciones y señales apropiadas de su microambiente, diferenciándose muchas veces en varios linajes con funciones y características especializadas como miocitos, neuronas o hepatocitos27. Por otro lado, están presentes en muchos tejidos adultos y su importancia radica en mantener la homeostasis celular y en la reparación de los tejidos28,29. Las etapas del proceso de neurogénesis hipocampal son: a) proliferación donde se generan las células progenitoras neurales generando un 1 % de células granulares por día, b) diferenciación de las células progenitoras neurales mayoritariamente en neuronas y en menor proporción en células gliales (astrocitos, oligodendrocitos, y microglía), c) migración cercana del SGZ al GD donde se produce la maduración, d) contacto dendrítico a los 14 días aproximadamente enviando su axón hacia CA3 (Cuerno de Amón) contactando allí con otras neuronas y e) integración sináptica donde tiene lugar la formación de células granulares maduras, interaccionando con diferentes neuronas diana30,31.
Descripción anatómica del hipocampo
Hablar de neurogénesis es hablar también de estructuras anatómicas, como un vínculo fisiológico referido a un microambiente con un conjunto de componentes celulares y no celulares que interactúan con la masa glíal y poblaciones neuronales, dándoles soporte físico y modulando su actividad química a través de un balance entre proliferación y auto mantenimiento, bajo el contexto de los "nichos neurogénicos" descritos anteriormente.
Uno de los órganos importantes reconocido como principal centro neurogénico es el Hipocampo, que es una estructura cerebral con forma de "caballito de mar", que forma parte del sistema límbico y se ubica en el lóbulo temporal y va desde el hipotálamo hasta la amígdala.
Esta parte del cerebro está relacionada con la memoria. En cuanto a su anatomía se encuentra el subículo, el cuerno de Amón (dividido en tres secciones: CA1, CA2 y CA3) y GD, este último considerado como principal nicho neurogénico32,33. El GD se asemeja a una "C" y se extiende desde el septum al lóbulo temporal del cerebro, presentando una organización en capas: la capa de somas de las células granulares y el hilus que delimitan la SGZ y que contiene los precursores neuronales, la capa molecular constituida por tres subcapas (interna, medial y externa) donde las neuronas granulares del GD presenta su aferencia y su eferencia en el hilus y cuerno de Amón (en particular en la zona CA3). Estas neuronas granulares poseen axones de diferente procedencia que son específicas a los estímulos que llegan a cada porción del árbol dendrítico de las células granulares34.
El mejor modelo anatómico que permite explicar esta estructura y sus distintas proyecciones estructurales es la de los roedores, debido a la disposición sagital a partir de su eje septo-temporal, perfectamente reconocible en los procesos de disección in situ 35. En los roedores la neo corteza manda proyecciones al hipocampo dorsal y las proyecciones del hipocampo ventral vienen de la corteza prefrontal, el hipotálamo, la amígdala y otros núcleos subcorticales36,37 (Fig.3).
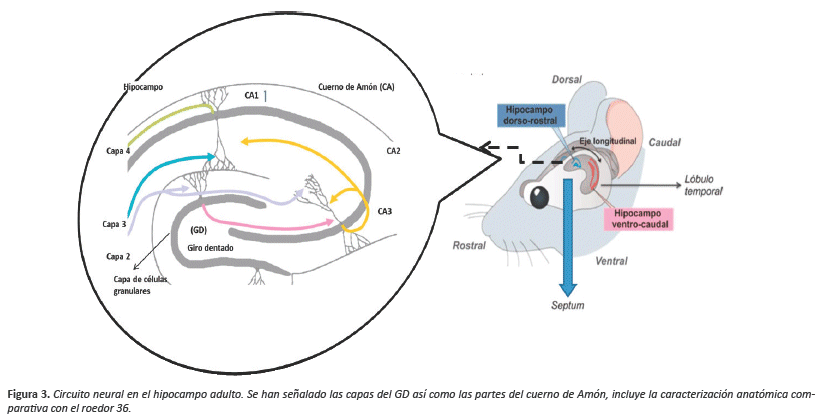
Las células madre en la capa de células granulares son una población de astrocitos similares a las que se encuentran en SVZ, las cuales son capaces de dividirse siendo precursores de neuronas, ya que posteriormente son capaces de alcanzar la capa granular pasando a ser neuronas granulares maduras38.
Principales mecanismos de señalización y regulación en la neurogénesis hipocampal.
La neurogénesis producida en la zona del hipocampo recurre a mecanismos de señalización. Se ha demostrado que la capacidad neurogénica tiene lugar en la zona subventricular de los ventrículos laterales (SVZ) y en la zona subgranular del giro dentado. Dicho fenómeno se debe a que las células madre neuronales encuentren en dicha zona un microambiente adecuado para dividirse y general neuronas. Estose demostró cuando se trasplantaron precursores de la SVZ de un animal en la misma zona de otro animal, mientras que si dicho trasplante tiene lugar en otra zona del cerebro dichas células no sobreviven (en algunos casos dichos precursores se convirtieron en glía).Todo esto nos indica que para que tenga lugar la neurogénesis necesitamos la presencia de las células pluripotenciales y además que estas estén inmersas en un nicho neurogénico adecuado (GD). Una de las moléculas esenciales que deben encontrarse en estos nichos neurogénicos son los factores de crecimiento (proteínas) los cuales permiten regular los procesos de formación, reparación, supervivencia neuronal, proliferación y migración de las nuevas células neuronales. Estos factores de crecimiento pueden ser: factores de crecimiento de fibroblastos (FGF), factores de crecimiento endotelial (EGF), factor de crecimiento endotelial vascular (VEGF), factor de crecimiento derivado del epitelio pigmentario (PEDF) y algunos neurotransmisores como GABA (ácido v-aminobutírico)39,40. Por otro lado, también se encuentra el factor neurotrófico ciliar (CNTF) derivado de la familia de las citoquinas neuropoyéticas, encargado de promover el mantenimiento de las nuevas células neuronales en el adulto y de gran actividad en lesiones neurales. También hay que tener en cuenta que la presencia de elementos químicos afecta en gran medida en los mecanismos de señalización, como son las bases purinas, las cuales contienen altas concentraciones de bases nitrogenadas y regulan negativamente la diferenciación neuronal en el caso de la SVZ, debido a que sus señales proliferativas son específicas para progenitores neurales41. Otra molécula química presente es el óxido nítrico (NO) inhibiendo la proliferación de las neuronas nitrérgicas vía mitocondrial42.
Finalmente la evidencia experimental demuestra que factores de crecimiento como las neurotrofinas, moléculas pequeñas como los neuropéptidos y biomoléculas como los neurotransmisores (NT) que generan un microambiente apropiado para las células madre permitiendo el crecimiento de nuevas neuronas del SNC, promoviendo su ruta hacia su localización final, así como modulando el crecimiento axonal su función43. No se debe olvidar que los factores sociales también pueden modular el proceso de la neurogénesis así como la actividad física, y la interacción social44.
Integración neuroquímica de las nuevas neuronas en el circuito hipocampal.
El proceso de neurogénesis tiene lugar en varias etapas: proliferación, migración, diferenciación, y maduración en los circuitos existentes. El proceso de integración de las nuevas células neuronales en el circuito hipocampal, recurre a varias etapas, empezando con el silenciamiento sináptico, estimulación GABAérgica de carácter excitatorio, ingresos glutamatérgicos y estimulación GABAérgica de carácter inhibitorio45,46.
En la estructura del cerebro adulto el GABA es el mayor neurotransmisor inhibidor, pero también ejerce diferentes funciones sobre las células pluripotentes y su progenie, quedando demostrada la presencia de receptores GABAérgicos en precursores neurogénicos47,48. Una vez que se determina el fenotipo neuronal en las nuevas neuronas inmaduras estas son activadas por GABA (efecto excitador) debido a las altas concentraciones de iones cloro intracelular, permitiendo recibir entradas sinápticas liberadas en sitios extra-sinápticos. Este "efecto excitatorio" del GABA modula el umbral de excitabilidad de las nuevas células neuronales, así como su maduración, integración y función49,50 (Figura 4).
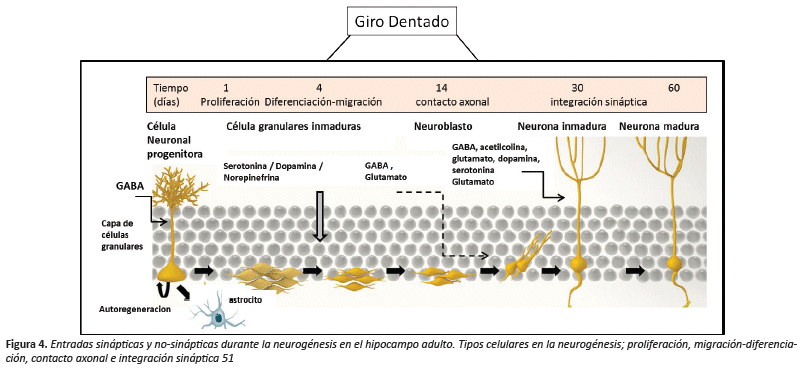
Las inclusiones de elementos glutamatérgicos en las nuevas células neuronales son evidentes una vez superada la fase de supervivencia temprana, sin embargo, muestran un umbral inferior para sus procesos de inducción y potenciación sináptica duradera (LTP, del inglés "long-term potentiation") respecto a las neuronas granulares maduras. Este fenómeno ha sido descrito en la expresión proteica del Receptor Ionotrópico Glutamato (NR2B) y también del receptor N-Metil D-Aspartato (NMDA) que contribuye a que las NCN tengan un menor umbral para la inducción de LTP. El estado fisiológico del hipocampo podría regular el reclutamiento de neuronas jóvenes del giro dentado en redes neuronales, así como su integración en los circuitos hipocampales a través de la señalización GABAérgicos52.
A nivel perisomático se registran las primeras corrientes GABAérgicas, una vez ocurridas las inclusiones glutamatérgicas, recibiendo las neuronas inmaduras entradas inhibidoras locales de las tres subregiones del GD: capa molecular, capa de células granulares e hilus. Finalmente en la fase de la maduración, la corriente GABAérgica inhibitoria se va incrementando gradualmente, debido al aumento de conexiones sinápticas y de su fuerza de conducción53.
Avances y estrategias para el estudio de la Neurogénesis adulta.
Dentro de los avances más significativos en el campo de la Neurogénesis se encuentra el relacionado con el implante de tejido neural embrionario con el fin de restaurar los niveles de dopamina en el cuerpo estriado, pudiendo llevar a resultados alentadores ya que se manifiesta un proceso de recuperación a largo plazo. Este proceso consiste en la liberación de una molécula (ácido láctico) que actúa como señal inductora de la angiogénesis. Además, dichos implantes crean un entorno adecuado para la generación de nichos neurogénicos. Este procedimiento tiene la limitación para el uso clínico por su escasa disponibilidad, por ello la adopción de células troncales como material de partida para terapia regenerativa. Otros estudios han descrito experiencias de supervivencia neuronal en neuronas dopaminérgicas que pueden volver a inervar el cuerpo estriado durante largos períodos (más de 10 años), restaurando la activación cortical frontal, sobre todo en enfermedades neurodegenerativas como el Parkinson54.
En el caso de enfermedades neurodegenerativas periféricas se avanzó con modelos experimentales de desmielinización, logrando la diferenciación de las células troncales en oligodendrocitos, inyectando neuroesferas (células troncales de la ZSV) por vía intravenosa, obteniendo resultados positivos en la remielinización de nervios periféricos como de la médula espinal55.
Si bien estudios in vitro e in vivo han demostrado que tanto las células troncales embrionarias como las CPN adultas son capaces de diferenciarse en neuronas dopaminérgicas 56, queda poco claro hasta qué punto pueden estas nuevas células neuronales establecer nuevos circuitos funcionales. Factores como la modelización del microambiente, las variables en los neurotransmisores y la diferenciación celular del tejido embrionario deja la exploración del circuito hipocampal en un ámbito interesante y amplio no solamente desde el punto de vista anatómico sino también desde la perspectiva moderna del análisis químico molecular.
DISCUSIÓN
Una de las hipótesis más aceptadas infiere que el hipocampo es uno de los principales nichos neurogénicos, debido al requerimiento de las muchas tareas que genera este órgano, entre las cuales se pueden mencionar, el aprendizaje espacial, la discriminación de patrones espaciales y el condicionamiento contextual al miedo57.
Un punto de discusión interesante ha sido determinar el origen de las nuevas neuronas corticales, sugiriendo como primera teoría a la SVZ como primer punto de origen, como sucede con las nuevas neuronas granulares y periglomerulares del bulbo olfatorio. La segunda teoría hace referencia al neuroepitelio del SN adulto, ya que este epitelio retendría algunos tipos celulares, tales como las células de la glía radial, que servirían probablemente como precursores neurales equiparables a las células neuroepiteliales embrionarias, con capacidades para responder a los patrones de señales inductoras de neurogénesis en el embrión58. Por tanto, la hipótesis sustentaría la idea de que las células troncales neuronales se desarrollan de un linaje neuroepitelial tipo glía radial astrocítico, pudiendo estas tener distintos precursores.
Los grandes avances en el campo de la histología han permitido el descubrimiento de células madre latentes en áreas corticales, bulbo olfatorio, giro dentado, hipocampo y locus Niger. Por otro lado, la incorporación de nuevas técnicas inmunohistoquímicas y moleculares que utilizan marcadores neurales más específicos, han contribuido a que la neurogénesis sea un proceso ampliamente investigativo y con una gran base experimental59.
Si bien los recientes estudios concluyen que los CPN mantienen la capacidad de autoregeneración (característica de las células troncales), sólo se ha demostrado que pueden dar origen a tipos celulares específicos (astrocitos, oligodendrocitos y neuronas)60, quedando todavía poco claro el papel de las células gliales en la neo-formación de las sinapsis neuronales debido a sus particularidades histoquímicas de interacción.
CONCLUSIONES
Actualmente la perspectiva científica sobre la contribución de la neurogénesis hipocampal adulta a los circuitos cerebrales y procesos cognitivos es un tema altamente en auge en el ámbito de la neurociencia. La idea de incrementar la proliferación de células nerviosas neuronales en las zonas neurogénicas del cerebro adulto promoviendo su migración, diferenciación, integración y supervivencia en las redes preexistentes con el fin de promover una recuperación funcional en los casos de daños tisulares de tejido nervioso por causas traumáticas, envejecimiento o procesos neurodegenerativos (Parkinson, Alzheimer, Huntington y en la esclerosis múltiple) sería un gran avance en Medicina. El proceso de neurogénesis hipocampal tiene lugar en el SVZ y en la zona subgranular del giro dentado, donde la presencia de Factores de Crecimiento es esencial para los "nichos neurogénicos"en el que las células madre neuronales puedan dividirse y generar neuronas. A pesar de producirse una ligera reducción en la cantidad de neuronas generadas durante el envejecimiento, un gran número de estas neuronas se encuentra aún presente en aquellos individuos que no padecen ninguna enfermedad neurológica al menos hasta edad muy avanzada. La detección de nuevas neuronas es muy compleja ya que su tratamiento requiere el uso de compuestos químicos que descomponen y modifican dichas muestras.
REFERENCIAS
1.- Thompson DM., Koppes AN., Hardy JG., Schidt CE. Eslectrical stimuli in the central nervous system microenvironment. Ann Rev Biomed Eng. 2014; 16, 397-430. Disponible en: doi: 10.1146/annurev-bioeng-121813-120655
2.-Mateos-Aparicio P. Rodriguez-Moreno A. The impact of studying brain plasticity. Front. Cell. Neurosci., 2019; 13,66. Disponible en: doi.org/10.3389/fncel.2019.00066
3.-Haladaj R. Anatomical variations of the dentate gyrus in normal adult brain. Surg Radiol Anat. 2020; 42(2),193-199. Disponible en: doi: 10.1007/s00276-019-02298-5
4.-Gordon P. Animal models and the question of adult neurogenesis. Taconic. 2019; 21. Disponible en: https://www.taconic.com/taconic-insights/neuroscience/neurogenesis-animal-models.html# [ Links ]
5.- Bernier P.J., Bedard A., Vinet J., Levesque M. Parent, A. Newly generated neurons in the amígdala and adjoining cortex of adult primates. Proceedings of the National Academy of Sciences USA, 2001; 99: 11464–11469. Disponible en: https://pubmed.ncbi.nlm.nih.gov/12177450-newly-generated-neurons-in-the-amygdala-and-adjoining-cortex-of-adult-primates/
6.- Martinez-Cerdeño V. Stephen C. Noctor S. C. Neuronal progenitor cell terminology. Front. Neuroanat.; 2018,6. Disponible en: https://doi.org/10.3389/fnana.2018.00104 [ Links ]
7.- Dayer A.G., Cleaver K.M., Abou Antoun T. Cameron H.A. New GABAergic interneurons in the adult neocortex and striatum are generated from different precursors. Journal of Cell Biology, 2005; 168: 415–427. Disponible en: https://pubmed.ncbi.nlm.nih.gov/15684031-new-gabaergic-interneurons-in-the-adult-neocortex-and-striatum-are-generated-from-different-precursors/
8.- Cameron H.A. Dayer A.G. New interneurons in the adult neocortex: small, sparse, but significant? Biological Psychiatry, 2008; 63: 650-655. Disponible en: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2423203/ [ Links ]
9.- Feliciano D.M. Bordey A. Newborn cortical neurons: only for neonates?. Trends in Neurosciences. 2013; 36, 51–61. Disponible en: https://pubmed.ncbi.nlm.nih.gov/23062965-newborn-cortical-neurons-only-for-neonates/
10.- Yuan T-F., Liang Y-X., So, K-F. Occurrence of new neurons in the piriform cortex. Frontiers in Neuroanatomy, 2015; 8:167.Disponible en: https://www.frontiersin.org/articles/10.3389/fnana.2014.00167/full [ Links ]
11.- Suhonen J.O., Peterson D.A., Ray J. Gage F.H. Differentiation of adult hippocampus derived progenitors into olfactory neurons in vivo. Nature, 1996; 383:624-627.Disponible en: https://pubmed.ncbi.nlm.nih.gov/8857538-differentiation-of-adult-hippocampus-derived-progenitors-into-olfactory-neurons-in-vivo/ [ Links ]
12.-Ming G.L. Song H. Adult neurogenesis in the mammalian central nervous system. Annual Review of Neuroscience, 2005; 28:223-250.Disponible en: https://pubmed.ncbi.nlm.nih.gov/16022595-adult-neurogenesis-in-the-mammalian-central-nervous-system/ [ Links ]
13.- Gross C.G. Neurogenesis in the adult brain: Death of a dogma. Nature Reviews Neuroscience, 2000; 1:67-73. Disponible en: https://pubmed.ncbi.nlm.nih.gov/11252770-neurogenesis-in-the-adult-brain-death-of-a-dogma/ [ Links ]
14.- Altman J. Autoradiographic investigation of cell proliferation in the brains of rats and cats. Anatomical Record, 1963; 145: 573-591. Disponible en: https://onlinelibrary.wiley.com/doi/abs/10.1002/ar.1091450409 [ Links ]
15.- Altman J. Autoradiographic and histological studies of postnatal neurogenesis, IV. Cell proliferation and migration in the anterior forebrain, with special reference to persisting neurogenesis in the olfactory bulb. Journal of Comparative Neurology, 1969; 137(4): 433-457. Disponible en: https://pubmed.ncbi.nlm.nih.gov/5361244-autoradiographic-and-histological-studies-of-postnatal-neurogenesis-iv-cell-proliferation-and-migration-in-the-anterior-forebrain-with-special-reference-to-persisting-neurogenesis-in-the-olfactory-bulb/ [ Links ]
16.- Goldman S.A. Nottebohm F. Neuronal production, migration, and differentiation in a vocal control nucleus of the adult female canary brain. Proc. Natl. Acad. Sci. U. S. A 1983; 80, 2390-2394. Disponible en: https://pubmed.ncbi.nlm.nih.gov/6572982-neuronal-production-migration-and-differentiation-in-a-vocal-control-nucleus-of-the-adult-female-canary-brain/ [ Links ]
17.- Paton J.A. Nottebohm F.N. Neurons generated in the adult brain are recruited into functional circuits. Science 1984; 225, 1046-1048. Disponible en: https://pubmed.ncbi.nlm.nih.gov/6474166/ doi: 10.1126/science.6474166. [ Links ]
18.- Reynolds B. A. Weiss S. Generation of neurons and astrocytes from isolated cells of the adult mammalian central nervous system. Science, 1992; 255 (5052): 1707-1710. Disponible en: https://pubmed.ncbi.nlm.nih.gov/6474166-neurons-generated-in-the-adult-brain-are-recruited-into-functional-circuits/ [ Links ]
19.- Rakic P. Limits of neurogenesis in primates. Science 1985; 227, 1054-1056. Disponible en: https://pubmed.ncbi.nlm.nih.gov/3975601-limits-of-neurogenesis-in-primates/ [ Links ]
20.- Gage F.H. Mammalian neural stem cells. Science 2000; 287: 1433-8. Disponible en: https://pubmed.ncbi.nlm.nih.gov/10688783-mammalian-neural-stem-cells/ [ Links ]
21.- Eriksson PS, Perfilieva E, Björk-Eriksson T, Alborn AM, Nordborg C, Peterson DA, et al. Neurogenesis in the adult human hippocampus. Nat Med 1998; 4: 1313-7. Disponible en: https://pubmed.ncbi.nlm.nih.gov/9809557-neurogenesis-in-the-adult-human-hippocampus/ [ Links ]
22.- Arias-Carrión O, Olivares-Bañuelos T, Drucker-Colín R. Neurogénesis en el cerebro adulto. Rev Neurol 2007; 44: 541-550. Disponible en: https://ginde.webs.ull.es/wp-content/uploads/2013/06/neurogenesis.pdf [ Links ]
23.- Abrous DN, Koehl M, Le Moal M. Adult neurogenesis: from precursors to network and physiology. Physiol Rev 2005; 85: 523-569. Disponible en: https://pubmed.ncbi.nlm.nih.gov/15788705-adult-neurogenesis-from-precursors-to-network-and-physiology/. [ Links ]
24.-Ghosh H.S. Adult Neurogenesis and the Promise of adult neural stem cells. J. Exp Neurosci. 2019; 13:1-12. Disponible en: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6600486/ [ Links ]
25.- Palmer T.D., Willhoite A.R. Gage, F.H. Vascular niche for adult hippocampal neurogenesis. Journal of Comparative Neurology, 2000; 425: 479–494. Disponible en: https://pubmed.ncbi.nlm.nih.gov/10975875-vascular-niche-for-adult-hippocampal-neurogenesis/
26.-Peng L. Bonaguidi M.A. Function and dysfunction of Aduct hippocampal Neurogenesis in Regeneration and Disease. The American Journal of Pathology, 2018; 188 (1): 23-28. Disponible en: https://www.sciencedirect.com/science/article/pii/S0002944017305916 [ Links ]
27.- Ihn Han B-S K, Hun-Kuk Park K S K. Differentiation Potential of Mesenchymal Stem Cells Is Related to Their Intrinsic Mechanical Properties. Int Neurourol J. 2017; 21: S24-31. Disponible en: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5426435/ [ Links ]
28.- Ni F, Yu WM, Wang X, Fay ME, Young KM, Qiu Y, et al.. Ptpn21 Controls Hematopoietic Stem Cell Homeostasis and Biomechanics. Cell Stem Cell. 2019; 424(4):608-620.e6. Disponible en: https://pubmed.ncbi.nlm.nih.gov/30880025-ptpn21-controls-hematopoietic-stem-cell-homeostasis-and-biomechanics/ [ Links ]
29.- Dulak J, Szade K, Nowak W, Jozkowicz A. Adult stem cells: hopes and hypes of regenerative medicine. Acta Biochim Pol. 2015; 62(3):329-337. Disponible en: https://pubmed.ncbi.nlm.nih.gov/26200199-adult-stem-cells-hopes-and-hypes-of-regenerative-medicine/ [ Links ]
30.- Zhao C., Teng E.M., Summers R.G. Jr., Ming G.L. Gage, F.H. Distinct morphological stages of dentate granule neuron maturation in the adult mouse hippocampus. Journal of Neuroscience, 2006; 26(1): 3-11. Disponible en: https://pubmed.ncbi.nlm.nih.gov/16399667-distinct-morphological-stages-of-dentate-granule-neuron-maturation-in-the-adult-mouse-hippocampus/ [ Links ]
31.- Toni N., Laplagne D.A., Zhao C., Lombardi G., Ribak C.E., Gage F.H. Schinder, A.F. Neurons born in the adult dentate gyrus from functional synapses with target cells. Nature Neuroscience, 2008; 11(8):901-907. Disponible en: https://pubmed.ncbi.nlm.nih.gov/18622400-neurons-born-in-the-adult-dentate-gyrus-form-functional-synapses-with-target-cells/ [ Links ]
32.- Altman J. Are new neurons formed in the brains of adult mammals? Science, 1962a; 135(3509): 1127-1128. Disponible en: https://pubmed.ncbi.nlm.nih.gov/13860748-are-new-neurons-formed-in-the-brains-of-adult-mammals/
33.- Altman J. Autoradiographic study of degenerative and regenerative proliferation of neuroglia cells with tritiated thymidine. Experimental Neurology, 1962b; 5: 302-318. Disponible en: https://www.sciencedirect.com/science/article/pii/0014488662900407
34.- Forster E., Zhao S. Frotscher, M. Laminating the hippocampus. Nature Reviews Neuroscience, 2006; 7:259-267. Disponible en: https://pubmed.ncbi.nlm.nih.gov/16543914-laminating-the-hippocampus/ [ Links ]
35.- Amaral D.G. Witter M.P. The three-dimensional organization of the hippocampal formation: a review of anatomical data. Neuroscience, 1989; 31:571-591.Disponible en: https://pubmed.ncbi.nlm.nih.gov/2687721-the-three-dimensional-organization-of-the-hippocampal-formation-a-review-of-anatomical-data/ [ Links ]
36.- Suárez I. Papel de la Neurogénesis hipocampal adulta en los procesos cognitivos que dependen del hipocampo. Tesis, Universidad de Olavide, Sevilla 2015, España.
37.- Strange B.A., Witter M.P., Lein E.S. Moser E.I. Functional organization of the hippocampal longitudinal axis. Nature Reviews Neuroscience, 2014; 15(10):655-569. Disponible en: https://pubmed.ncbi.nlm.nih.gov/25234264-functional-organization-of-the-hippocampal-longitudinal-axis/ [ Links ]
38.- Seri B., García-Verdugo J.M., McEwen B.S. Alvarez-Buylla A. Astrocytes give rise to new neurons in the adult mammalian hippocampus. Journal of Neuroscience, 2001;21 (18): 7153-7160. Disponible en: https://pubmed.ncbi.nlm.nih.gov/11549726-astrocytes-give-rise-to-new-neurons-in-the-adult-mammalian-hippocampus/ [ Links ]
39.-Ge SY, Pradhan DA, Ming GL, Song HJ. GABA sets the tempo for activity-dependent adult neurogenesis. Trends Neuroscience 2007; 30, 1-8. Disponible en: https://pubmed.ncbi.nlm.nih.gov/17116335-gaba-sets-the-tempo-for-activity-dependent-adult-neurogenesis/ [ Links ]
40.- Zhao C., Deng, W., Gage F.H. Mechanisms and functional implications of adult neurogenesis. Cell, 2008a; 132:645–660.Disponible en: https://pubmed.ncbi.nlm.nih.gov/18295581-mechanisms-and-functional-implications-of-adult-neurogenesis/
41.- Lin JH, Takano T, Arcuino G, Wang X, Hu F, Darzynkiewicz Z, Nunes M, Goldman SA, Nedergaard M. Purinergic signaling regulates neural progenitor cell expansion and neurogenesis. Dev Biol 2007; 302, 356-366.Disponible en: https://pubmed.ncbi.nlm.nih.gov/17188262-purinergic-signaling-regulates-neural-progenitor-cell-expansion-and-neurogenesis/ [ Links ]
42.- Moreno-López B, Romero-Grimaldi C, Noval JA, Murillo-Carretero M, Matarredona ER, Estrada C. Nitric Oxide Is a Physiological Inhibitor of Neurogenesis in the Adult Mouse Subventricular Zone and Olfactory Bulb. J Neurosci. 2004; 24:85-95. Disponible en: https://pubmed.ncbi.nlm.nih.gov/14715941-nitric-oxide-is-a-physiological-inhibitor-of-neurogenesis-in-the-adult-mouse-subventricular-zone-and-olfactory-bulb/ [ Links ]
43.- Bauer S, Kerr BJ, Patterson PH. The neuropoietic cytokine family in development, plasticity, disease and injury. Nat Rev Neurosci 2007; 8, 221-232.Disponible en: https://pubmed.ncbi.nlm.nih.gov/17311007-the-neuropoietic-cytokine-family-in-development-plasticity-disease-and-injury/ [ Links ]
44.- Kuhn H.G. Toda T. Gage F.H. Adult hippocampal Neurogenesis: A coming-of-Age-Story. Journal of Neuroscience, 2018; 38 (49): 10401-10410. Disponible en: https://www.jneurosci.org/content/38/49/10401 [ Links ]
45.- Espósito M.S., Piatti V.C., Laplagne D.A., Morgenstern N.A., Ferrari C.C., Pitossi F.J. Schinder A.F. Neuronal differentiation in the adult hippocampus recapitulates embryonic development. Journal of Neuroscience, 2005; 25(44): 10074-10086.Disponible en: https://www.jneurosci.org/content/25/44/10074 [ Links ]
46.- Vivar C, van Praag H. Functional circuits of new neurons in the dentate gyrus. Frontiers of Neural Circuits, 2013; 7-15. Disponible en: https://pubmed.ncbi.nlm.nih.gov/23443839-functional-circuits-of-new-neurons-in-the-dentate-gyrus/ [ Links ]
47.- Tozuka Y., Fukuda S., Namba T., Seki T. Hisatsune, T. GABAergic excitation promotes neuronal differentiation in adult hippocampal progenitor cells. Neuron, 2005: 47: 803–815.Disponible en: https://pubmed.ncbi.nlm.nih.gov/16157276-gabaergic-excitation-promotes-neuronal-differentiation-in-adult-hippocampal-progenitor-cells/
48.- Wang L.P., Kempermann G. Kettenmann H. A subpopulation of precursor cells in the mouse dentate gyrus receives synaptic GABAergic input. Molecular and Cellular Neuroscience, 2005; 29(2): 181-9. Disponible en: https://pubmed.ncbi.nlm.nih.gov/15911343-a-subpopulation-of-precursor-cells-in-the-mouse-dentate-gyrus-receives-synaptic-gabaergic-input/ [ Links ]
49.- Walker M.C. Semyanov, A. Regulation of excitability by extrasynaptic GABA (A) receptors. Results and Problems in Cell Differentiation, 2008; 44: 29–48. Disponible en: https://pubmed.ncbi.nlm.nih.gov/17671772-regulation-of-excitability-by-extrasynaptic-gabaa-receptors/
50.- Chancey J.H., Adlaf E.W., Sapp M.C., Pugh P.C., Wadiche J.I. Overstreet-Wadiche L.S. GABA depolarization is required for experience dependent synapse unsilencing in adult-born neurons. Journal of Neuroscience, 2013; 33(15):6614-22. Disponible en: https://pubmed.ncbi.nlm.nih.gov/23575858-gaba-depolarization-is-required-for-experience-dependent-synapse-unsilencing-in-adult-born-neurons/ [ Links ]
51.- Toda T., Gage F.H. Review: adult neurogenesis contributes to hippocampal plasticity. Cell and Tissue Research, 2018, 373:693–709.Disponible en: https://pubmed.ncbi.nlm.nih.gov/29185071-review-adult-neurogenesis-contributes-to-hippocampal-plasticity/
52.- Zhu Y. Gao H. Tong L. Li Z. Wang L. Zhang C. Yang Q. Yan B. Emotion regulation of Hyppocampus Using Real-Time fMRI Neurofeedback in Healthy Human. Front. Hum. Neurosci. 2019; 16. Disponible en: https://www.frontiersin.org/articles/10.3389/fnhum.2019.00242/full [ Links ]
53.- Li Y., Aimone J.B., Xu X., Callaway E.M. Gage F.H. Development of GABAergic inputs controls the contribution of maturing neurons to the adult hippocampal network. Proceedings of the National Academy of Sciences USA, 2012; 109: 4290-4295.Disponible en: https://www.pnas.org/content/109/11/4290 [ Links ]
54.- Drucker-Colín R. Verdugo-Díaz L. Cell transplantation for Parkinson’s disease: present status. Cell Mol Neurobiol 2004; 24:301-316.Disponible en: https://pubmed.ncbi.nlm.nih.gov/15206816-cell-transplantation-for-parkinsons-disease-present-status/
55.-Rodriguez-jimenez F.J. Clemente E. Moreno-Manzano V. Erceg S. Organized Neurogenic-Niche-Like Pinwheel Structures Discovered in Spinal Cord Tissue-Derived Neurospheres. Front. Cell Dev. Biol. 2019; 20. Disponible en: https://www.frontiersin.org/articles/10.3389/fcell.2019.00334/full [ Links ]
56.-Noisa P. Raivio T. Cui W. Neural Progenitor Cells derived from human Embryonic Stem Cells as an Origin of Dopaminergic Neurons. Stem Cells International. 2015; 1-10. Disponible en: https://www.hindawi.com/journals/sci/2015/647437/ [ Links ]
57.- Deng W., Aimone J.B. Gage, F.H. New neurons and new memories: how does adult hippocampal neurogenesis affect learning and memory? Nature Review Neuroscience, 2010; 11(5): 339-350.Disponible en: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2886712/ [ Links ]
58.- Cotman CW, Nieto-Sampedro M H. EW. Synapse replacement in the nervous system of adult vertebrates. Physiol. Rev. 1981; 61: 684-784.Disponible en: https://pubmed.ncbi.nlm.nih.gov/7019944-synapse-replacement-in-the-nervous-system-of-adult-vertebrates/
59.-Seki T. Hori T. Muyata H. Maehara M. Namba T. Analysis of proliferating neuronal progenitors and inmatyre neurons in the humen hippocampus surgically removed from control and epileptic patiens. Scientific Reports 2019; 9: 14194. Disponible en: https://www.nature.com/articles/s41598-019-54684-z [ Links ]
60.- Coronel R. Lachgar M. Bernabeu-Zornoza A. Palmer C. Domínguez-Alvaro M. Revilla A. Ocaña I. Fernández A. Martínez-Serrano A. Cano E. Liste I. Neuronal and Glial Differentiation of Human Neural Stem Cells Is Regulated by Amyloid Precursor Protein (APP) Levels. Mol Neurobiol 2019; 56(2):1248-1261. https://www.ncbi.nlm.nih.gov/pubmed/29881946 [ Links ]

