










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkActa Nova
On-line version ISSN 1683-0789
RevActaNova. vol.7 no.1 Cochabamba Mar. 2015
ARTÍCULO CIENTÍFICO
Ocurrencia de Batrachospermum macrosporum y su fase Chantransia (=Audouinella macrospora) en la cuenca del río Yacuma (Beni, Bolivia)
Ocurrence of Batrachospermum macrosporum and its Chantransia phase (=Audouinella macrospora) in the Yacuma river basin (Beni, Bolivia)
Eduardo A. Morales1, Sinziana F. Rivera1,2
1Herbario Criptogámico, Universidad Católica Boliviana "San Pablo". Calle M. Márquez esq. Plaza Jorge Trigo s/n, P.O. Box 5381, Cochabamba, Bolivia
2Maestría Universitaria en Ciencias del Medio Ambiente (MUSE), Instituto de Ciencias del Medio ambiente, Universidad de Ginebra. Route de Drize, 1227 Carouge, Ginebra, Suiza.
edu_mora123@outlook.com
Recibido: 20 de marzo 2015
Aceptado: 14 de abril 2015
Resumen: Reportamos el hallazgo de Batrachospermum macrosporum y su fase Chantransia en el río Yacuma y dos de sus tributarios, localizados dentro del Área Protegida Municipal Pampas del río Yacuma, Beni, Bolivia. Se detallan las características morfológicas a nivel macro y microscópico y se discute la ecología en función a observaciones propias, consulta con expertos y revisión de la literatura. Proponemos que la investigación de algas rojas se hace urgente ante una carencia casi total de su conocimiento a nivel nacional y de un escenario de acelerada degradación ambiental. También se sugiere que las fases gametofítica y esporofítica de B. macrosporum pueden ser utilizadas para el monitoreo de la calidad del agua en una zona del Yacuma que es severamente afectada por actividades antropogénicas.
Palabras clave: Algas rojas, áreas protegidas, Audouinella, Batrachospermum, Bolivia, Chantransia, Rhodophyta, río Yacuma.
Abstract: We report the finding of Batrachospermum macrosporum and its Chantransia phase in the Yacuma River and two of its tributaries, located within the Yacuma Plains Municipal Protected Área, Beni, Bolivia. Details are given of the morphological features at the macro and microscopic levéis, and the ecology is discussed based on own observations, consultation with experts and revisión of the literature. We propose that research on red algae is urgent due to an almost complete lack of knowledge at the national level and in the face of an accelerated environmental degradation in the country. It is also suggested that the gametophyte and sporophyte phases of B. macrosporum can be utilized to monitor water quality in a zone of the Yacuma that is severely affected by anthropogenic activity.
Keywords: Audouimlla, Batrachospermum, Bolivia, Chantransia, red algae, protected areas, Rhodophyta, Yacuma River.
1. Introducción
Las algas rojas (División Rhodophyta) que habitan aguas epicontinentales son diversas, aunque solamente representan un 3 a 5% de la diversidad total de algas rojas [16]. Las formas dulceacuícolas se clasifican en 9 órdenes distintos y el número de especies descritas varía entre 200 a 300 [7][19].
Las historias de vida de las algas rojas están dentro de las más complejas entre los organismos fotosintéticos e incluyen, de manera general, una alternancia haplodiplonte de 3 generaciones heteromorfas (gametofito-carposporofito-tetrasporofito). El gametofito haploide es dominante y produce gametos por mitosis. Después de la fecundación del óvulo contenido en una estructura llamada carpogonio, allí mismo desarrollan una serie de filamentos denominados gonimoblastos (o ramas carpogoniales) que constituyen la fase carposporofítica, parasítica sobre el gametofito. El carposporofito produce carposporas diploides por mitosis, las cuales desarrollan en tetrasporofitos independientes también diploides. Los tetrasporofitos a su vez se reproducen meióticamente por tetrasporas haploides. Cada tetraspora genera un nuevo gametofito, reiniciando el ciclo. Entre las algas rojas, existen numerosas variaciones de esta historia de vida generalizada, incluyendo la alternancia de generaciones gametofítica y esporofítica isomorfas, es decir, prácticamente indistinguibles mediante la observación de sus partes vegetativas.
El orden Batrachospermales contiene aproximadamente unas 150 especies, todas con distribución continental y que prefieren ríos de flujo bajo [16][6]. Las especies de Batrachospermum, el género más especioso dentro de las Batrachospermales, se hallan ampliamente distribuidas en América y otros continentes, con varios casos de endemismo [4]. Las carposporas de Batrachospermum producen filamentos uniseriados y ramificados correspondientes al esporofito. Estos filamentos fueron clasificados dentro del género Chantransia cuando aún no se conocía bien la historia de vida de Batrachospermum [18]; hoy la fase esporofítica de Batrachospermum todavía se conoce como la "fase Chantransia" de su ciclo vital. Los filamentos del esporofito producen monosporas por mitosis (las tetrasporas meióticas están ausentes del ciclo); el proceso meiótico ocurre en las células apicales del gametofito antes de producirse los gametos, de manera que el talo del gametofito es en parte diploide y en parte haploide [7]. Las monosporas pueden producir nuevos filamentos esporofíticos y, por tanto, la fase Chantransia puede extenderse indefinidamente dentro del ciclo vital [7] [14].
La especie Batrachospermum macrosporum Montagne está ampliamente distribuida en Norte, Centro y Sudamérica [16]. Su talo está típicamente ramificado en verticilos producidos en cada unión entre las células contiguas del filamento principal. Esta especie es relativamente fácil de reconocer por su coloración azul verdosa y oscura, que forma una especie de collares de cuentas embebidas en abundante mucilago; cada una de las cuentas corresponde a un verticilo de ramificación profusa.
Un segundo género que ha estado históricamente relacionado con Batrachospermum es Audouinella, que también pertenece a las algas rojas y también tiene representantes dulceacuícolas, pero que por sus características morfológicas y reproductivas se han colocado dentro del orden Achrochaetiales. Los filamentos de este género son también uniseriados y ramificados pero no tan profusamente ni en verticilos como los de Bartrachospermum y producen monosporangios que se hallan al término de ramificaciones cortas. La diversidad de Audouinella es más baja que la de Batrachopermum, con unas 20 especies que habitan en ríos de flujo variable desde zonas nórdicas hasta ecuatoriales [2].
Algunas especies de Audouinella tienen coloración rojiza, mientras que otras, como es el caso de Audouinella macropora (Wood) Sheath & Burkholder son de color azul verdoso. En 1997, Necchi y Zucchi [12] demostraron, mediante seguimiento de la reproducción y crecimiento en cultivos, que A. macropora era la fase esporofítica del ciclo vital de Batrachopermum, sugiriendo que todas las formas filamentosas azul verdosas incluidas actualmente en Audouinella están conectadas con especies de Batrachopermum. Pueschel et al. [14], trabajando con ARN ribosómico y las características de las punteaduras areoladas que comunican células contiguas en los filamentos de A. macrospora y B. macroporum, confirmaron ultraestructural y molecularmente que ambas son parte de una misma historia de vida. La coloración azul verdosa de tanto B. macrosporum como A. macropora se debe a la presencia de una ficoeritrina que no se halla en ninguna otra alga roja y que constituye otra característica unificadora entre ambas fases [3].
Los estudios sobre algas rojas de Bolivia son escasos y ninguno de ellos hace un recuento de la diversidad ni de la distribución de géneros o especies particulares en ecosistemas acuáticos del país [8]. Esta carencia se suma a la falta de información para otros grupos algales, dejando un gran potencial para estudios ficológicos de diversa índole y para el descubrimiento de una diversidad algal todavía no reportada [8][9][10].
Se presenta aquí el hallazgo simultáneo de los gametofitos y esporofitos (fase Chantransia) de B. macrosporum en el río Yacuma y 2 riachuelos afluentes, 3 meses después de un episodio desastroso de inundación correspondiente a la época de lluvias (noviembre, 2013 a abril, 2014) en el Departamento del Beni, Bolivia. Se presentan las características morfológicas de ambas fases, así como las características ecológicas generales de las localidades de muestreo.
2. Metodología
2.1 Zona de estudio
Cuatro muestras fueron colectadas entre el 9 y el 11 de julio, 2014, a lo largo del río Yacuma y dos afluentes dentro del Área Protegida Municipal Pampas del río Yacuma. Esta última está ubicada en el municipio de Santa Rosa del Yacuma, departamento del Beni, y ocupa una superficie de 616.453 ha, constituyéndose en una de las áreas protegidas municipales más grandes de Bolivia. La zona alberga y protege ecosistemas de humedales y sabanas inundables que cuentan con una alta diversidad biológica. El ecoturismo se constituye en una de las principales fuentes de ingresos económicos del municipio. Esta actividad se lleva a cabo principalmente a lo largo del río Yacuma donde se han instalado 10 albergues para alojar a los cerca de 16.000 turistas que visitan el lugar anualmente, con las implicaciones que esta actividad supone para el medio ambiente. Al interior del área protegida se encuentran también 9 comunidades campesinas (Mojón, San Cristóbal, San Bartolomé, Picaflores, Villa Fátima, Awaizal, El Triunfo, El Rosario y El Candado) así como 180 estancias ganaderas.
El río Yacuma es parte de la cuenca del río Madera que se extiende entre Bolivia y Brasil. El Yacuma nace en los últimos flancos de la Cordillera Oriental y fluye unos 570 km en sentido Norte hasta desembocar en el río Mamoré. En la época de lluvias (entre noviembre y abril, aproximadamente) el Yacuma desborda e inunda grandes extensiones de territorio. La inundación del 2013-2014 fue excepcionalmente mayor a la de años anteriores, causando muerte masiva en ganado vacuno y equino y obligando al gobierno central boliviano a rescatar y refugiar en campamentos provisorios a cientos de personas en la localidad de Rurrenabaque.
Las aguas del río Yacuma tienen velocidades relativamente bajas y son naturalmente turbias debido a los suelos arcillo-limosos por los que fluye y permanentemente erosiona. La ganadería, agricultura, navegación con motores fuera de borda y la influencia de estancias y albergues constituyen las fuentes de perturbación en el río que visiblemente contribuyen al ingreso de nutrientes y contaminantes al medio acuático (aunque no existen estudios formales sobre este aspecto).
2.2 Colecta de muestras y métodos de laboratorio
Las muestras colectadas corresponden a crecimientos filamentosos hallados adheridos a ramas semi-sumergidas de árboles ribereños, a una distancia de 1,5 y 10 km del albergue "Caracoles". Se colectaron también pequeñas porciones de ramas sumergidas de árboles vivos o de ramas en descomposición, halladas a lo largo de un brazo del río a 300 m del albergue "Caracoles" y de otro brazo que corre a lo largo del camino antiguo Rurrenabaque-Trinidad, a unos 5 km del albergue mencionado. Se colectaron en total 4 muestras, 2 del río Yacuma y una de cada tributario (Tabla 1).

En todos los casos, porciones de los filamentos fueron recogidos con la mano y guardados en bolsas zip-lok para su posterior traslado al laboratorio de la Universidad Católica "San Pablo", Cochabamba. Al tiempo de colecta de las muestras biológicas se midieron la temperatura, pH y conductividad eléctrica del agua del río (Tabla 1). Con la finalidad de identificar los organismos biológicos se hicieron preparaciones acuosas sobre portaobjetos utilizando Lugol para crear contraste en estructuras a nivel de células y filamentos. El análisis se realizó bajo un microscopio óptico Zeiss Universal equipado con una cámara digital Jenoptik ProgRes CF y a aumentos entre 125X y 1250X. Las láminas con fotografías se montaron utilizando Adobe Photoshop CS3 Extended v. 10.0 para Windows 7 Enterprise. La identificación se llevó a cabo utilizando literatura especializada (ver Resultados) y consulta a especialistas. Las fotografías en campo se tomaron con una cámara Sony Cyber-shot HD AVCHD Progressive con un lente Sony Lens G de 20X de aproximación. Las muestras aquí descritas fueron depositadas en la colección del Herbario Criptogámico de la Universidad Católica Boliviana "San Pablo" (HCUCB), bajo los códigos HCUCB-1075, HCUCB-1077 (B. macrosporum) y HCUCB-1073, HCUCB-1076 (fase Chantransia).
2.3 Resultados y Discusión
En las fechas que se colectaron las muestras del río Yacuma, colectas realizadas en la zona de mayor concentración de albergues ecoturísticos o zonas inmediatamente aledañas, todavía existía vegetación arbórea ribereña semi-sumergida en las aguas del río (Figs 1A-B, ID-E). De muchas de estas ramas crecían los filamentos de los gametofitos de apariencia catenulada (Figs 1B y 1C). Estos crecimientos son conspicuos ya que se ubican inmediatamente por debajo de la superficie del agua y se extienden como mechones de cabellos de color verde oscuro. Al tacto, los crecimientos son duros y resbalosos debido a la matriz mucilaginosa en la que se hallan embebidos. La matriz envuelve a cada verticilo de ramificaciones laterales, dando el aspecto de un filamento de cuentas o cadena de pequeñas bolitas translúcidas (Fig. 1C).

En el gametofito, el filamento principal (eje axial) está compuesto por una sola hilera de células que varían en tamaño desde aproximadamente 70 X 30 μm, hacia la base del eje, hasta unas 15 X 15 μm en el extremo distal (Figs 2A-C y 3D). Como consecuencia de la variación de la longitud de las células del eje axial, los verticilos de fascículos laterales también se hallan más alejados o más cercanos y profusos, yendo de la base hacia la punta del eje axial (Figs 2A-C). Los fascículos laterales empiezan con células más alargadas en su origen en las zonas nodales del eje axial y terminan en células mucho más pequeñas en su extremo distal (Figs 2B-D) y existe una extensa ramificación secundaria produciendo en total de 5 a 10 células desde el origen del fascículo hasta su terminación (Figs 3A-C). El ápice de las ramificaciones secundarias muchas veces termina en una célula que posee una prolongación notoria (Figs 2D y 3C).
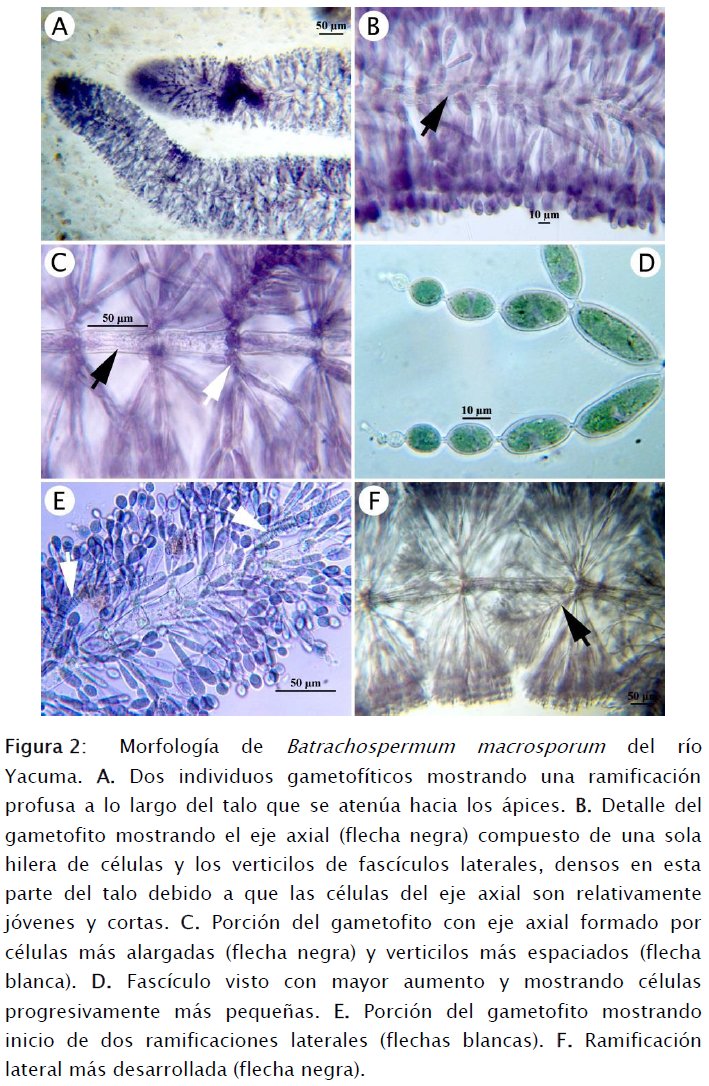
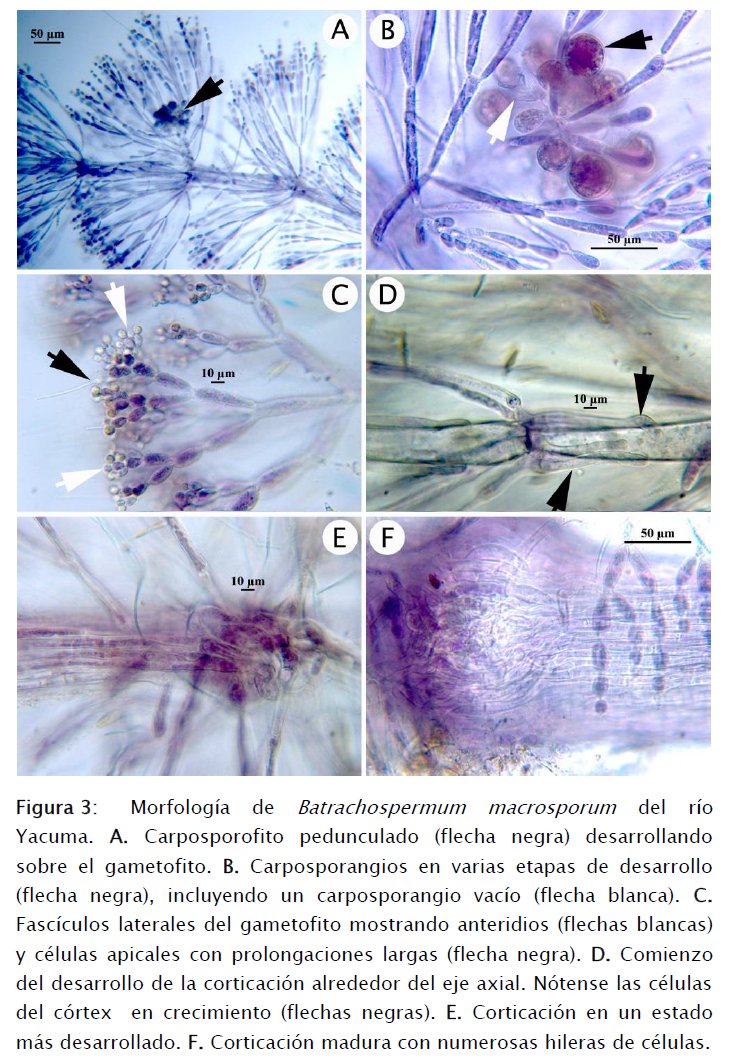
También existen ramificaciones laterales que luego crecen convirtiéndose en filamentos morfológicamente idénticos al principal. Al igual que los fascículos laterales, las ramificaciones crecen de las zonas nodales del eje axial del filamento principal como filamentos de células que son más anchas que largas (aproximadamente 4 X 11 μm, Fig. 2E). Finalmente, las células se alargan y verticilos laterales crecen de las zonas nodales de la nueva ramificación (Fig. 2F).
Las ramas carpogoniales y las tricóginas (estructuras sexuales femeninas) no fueron observadas por lo cual no aparecen ilustradas en el presente trabajo. O. Necchi (com. pers., 2015) asegura que la forma de las tricóginas no tiene relevancia decisiva en la identificación de B. macrosporum debido a que exhiben una amplia plasticidad fenotípica. Por tanto, aun cuando no las hayamos ilustrado, nuestra identificación del material hallado en la zona del Yacuma como B. macrosporum se valida con las otras características detalladas aquí.
Sí se observaron los anteridios, los cuales desarrollan en las puntas de las ramificaciones secundarias de los fascículos laterales. Estos anteridios son globosos y pequeños, midiendo aproximadamente unas 3-4 μm de diámetro (Fig. 3C).
Los carposporofitos son pedunculados y las células del pedúnculo se originan de la zona nodal del eje axial y están compuestas por 4 células relativamente pequeñas (30 X 16 μm) y sostienen a carpospoporangios obovoides de hasta 35 μm de diámetro (Figs 3A-B). Muchos de estos carposporangios estaban vacíos al tiempo de la colecta tal como se aprecia en la Fig. 3B.
Creciendo sobre el internodo o conexión entre células componentes del filamento axial, se hallan células alargadas y que recubren a la célula axial subyacente. Las Figuras 3D-F muestran la secuencia de este proceso de corticación que termina en un estado de desarrollo muy profuso y cuya finalidad es la de conferir resistencia a todo el talo del gametofito.
Las características del gametofito y carposporofito aquí detalladas concuerdan con las presentadas por Sheath et al. [17], quienes trabajaron con el material tipo de B. macrosporum.
También creciendo sobre ramas o sobre trozos de ramas sumergidas, en los tributarios del Yacuma que poseen corrientes más lentas, se hallaron crecimientos filamentosos que al ser inspeccionados bajo microscopía de luz tenían el color azul verdoso típico de A. macrospora (Figs 4A-D). Las células miden hasta 70 μm de largo y hasta 30 μm de ancho y poseen plastidios discoides. Los filamentos son ramificados y producen monosporangios elipsoides terminales que miden entre 20-25 μm de largo (Figs 4B-D). Necchi & Cabral [11] aseveran que en función a estudios filogenéticos, el hallazgo de A. macrospora en un ecosistema acuático es ya una indicación de que se trata de la fase Chantransia de B. macrosporum. Las características de la forma de crecimiento y color que aquí reportamos son también indicativas de esta conexión y están de acuerdo con lo establecido por Kato et al. [5], Necchi & Zucchi [12], Necchi et al. [13] y Pueschel et al. [14]).
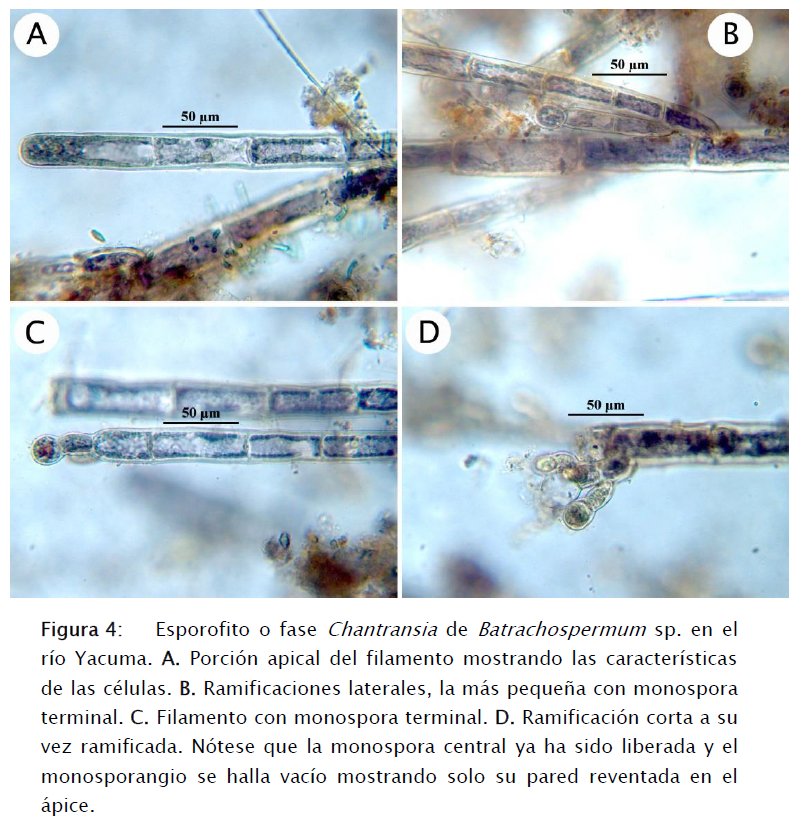
En la Tabla 1 se presentan las características fisicoquímicas de los 4 sitios de colecta en el río Yacuma y tributarios. Estos datos no difieren substancialmente de las características citadas por Necchi et al. [13] y Sheath et al. [17] para ríos de Norteamérica: temperatura 21° C; pH 6,1; conductividad 0,02 mS/cm; color del agua: amarillo. El pH del río Yacuma es ligeramente más neutral y el color en vez de amarillo es café claro debido a los sólidos en suspensión (en todos los sitios: 0,07 partes por mil). M. Vis (com. pers, 2015) asegura que B. macrosporum es una especie que crece en aguas negras y arenas blancas. Ninguno de los sitios en el Yacuma posee sustratos de arenas blancas, sino más bien de arcillas y limos que son suspendidos en la columna de agua impartiéndole la coloración café característica. Aunque en este estudio no se tomaron medidas de nutrientes ni indicadores bacterianos, es muy posible que la influencia de estancias, albergues, agricultura, ganado y tráfico de botes ocasione una fuerte descarga de contaminantes orgánicos que propicia el desarrollo de condiciones eutrófico-contaminadas (de hecho, se observan residuos sólidos flotando en tramos del río), las cuales favorecen el desarrollo de B. macrosporum. El hecho de que se haya encontrado B. macrosporum en el río Yacuma y de que esté asociada con aguas negras ofrece una realidad diferente a la que se ha descrito anteriormente en la que se pondera la efectividad del proceso de conservación en la cuenca (ver por ejemplo Shaw [15]). El uso de B. macrosporum y otras algas como indicadoras de la presión antrópica dentro del Área Protegida Municipal Pampas del río Yacuma merece ser evaluado con mayor profundidad.
Algas rojas han sido citadas con anterioridad para Bolivia (ver por ejemplo, Johnston et al. [4] y McClintic et al. [8]), sin embargo, ni Batrachopermum ni Audouinella figuran en la lista de Algas de Bolivia de Cadima et al. [1]. El presente estudio constituye el segundo reporte de Batrachopermum macrosporum y el primero de su fase Chantransia. Debido a la carencia de conocimiento de este grupo de algas en Bolivia y a la rápida degradación ambiental que están experimentando sus hábitats potenciales por la deforestación, contaminación y cambio climático, su estudio sistemático se hace imperativo.
3. Conclusión
El presente estudio es el primero a nivel detallado para algas rojas de Bolivia. Batrachospermum macrosporum, con sus fases gametofítica y esporofítica, está presente en el río Yacuma y tributarios del Departamento del Beni, Bolivia. Ambas fases del ciclo vital de esta alga roja poseen características morfológicas que las hacen fácilmente identificables y su ecología es similar a aquella presentada para otras partes del mundo. Ya que los ecosistemas acuáticos bolivianos están sufriendo una rápida degradación, el estudio de algas rojas y otros grupos algales es de máxima urgencia. En el caso específico de Batrachospermum macroporum y su fase Chantransia, su utilización como organismo indicador en el monitoreo de la calidad del agua debe ser explorado a profundidad.
Agradecimientos
Agradecemos la ayuda de los doctores M. Vis (Ohio University, EUA) y O. Necchi (Universidade Estadual Paulista Julio de Mesquita Filho, Brasil) en la identificación del material colectado para el presente trabajo y facilitación de bibliografía sobre algas rojas.
Rerefencias
[1] Cadima, F.M.M.; Fernández, T.E. & López, Z.L.F. 2005. Algas de Bolivia con énfasis en el fitoplancton. Importancia, ecología, aplicaciones y distribución de géneros. Fundación Simón I. Patiño. 377 pp. [ Links ]
[2] Garbary, D.J. 1987. The Acrochaetiaceae (Rhodophyta): an annotated bibliography. Bibliotheca Phycologica 77: 1-267. [ Links ]
[3] Glazer, A.N.; Chan, C.F. & West, J.A. 1997. A unsual phycocyanobilin-containing phycoerythrin of several bluish-colored acrochaetoid, freshwater red algal species. Journal of Phycology 33: 617-624. [ Links ]
[4] Johnston, E.T.; Lim, P-E.; Buhari, N.; Keil, E.J.; Djawad, M.I. & Vis, M.L. 2014. Diversity of freshwater red algae (Rhodophyta) in Malaysia and Indonesia from morphological and molecular data. Phycologia 53: 329-341. [ Links ]
[5] Kato, A.; Morita, N.; Hiratsuka, T. & Suda, S. 2009. Recent introduction of a freshwater red alga Chantransia macrospora (Batrachospermales, Rhodophyta) to Okinawa, Japan. Aquatic Invasions 4: 567-574. [ Links ]
[6] Kumano, S. 2002. Freshwater Red Algae of the World. Biopress Ltd, Bristol, 375 pp.
[7] Lee, R.E. 2008. Phycology. 4th edition. Cambridge University Press, USA, 547 pp.
[8] McClintic, A.S.; Casamatta, D.A. & Vis, M.L. 2003. A survey of algae from montane cloud forest and alpine streams in Bolivia: Macroalgae and associated microalgae. Nova Hedwigia 76: 363-379. [ Links ]
[9] Morales, E.A.; Fernández, E.; Fernandez, C.E.; Lizarro, D. & Alcoreza, P. 2008. Algal studies in Bolivia: a compilation and preliminary analysis of existing phycological literature. Gayana Botánica 65: 93-109. [ Links ]
[10] Morales, E.A.; Wetzel, C.E.; Rivera, S.F.; Van de Vijver, B. & Ector, L. 2014. Current taxonomic studies on the diatom flora (Bacillariophyceae) of the Bolivian Altiplano, South America, with possible consequences for palaeoecological assessments. Journal of Micropalaeontology: 33: 121-129. [ Links ]
[11] Necchi, O. Jr. & Cabral, O.M. 2011. Phylogenetic affinities of “Chantransia” stages in members of the Batrachospermales and Thoreales (Rhodophyta). Journal of Phycology 47: 680-686.
[12] Necchi, O. Jr. & Zucchi, M.R. 1997. Audouinella macrospora (Achrochaetiaceae, Rhodophyta) is the Chantransia stage of Batrachospermum (Batrachospermaceae). Phycologia 36: 220-224. [ Links ]
[13] Necchi, O.Jr.; Sheath, R.G. & Cole, K.M. 1993. Systematics of freshwater Audouinella (Acrochaetiaceae, Rhodophyta) in North America. 2. The bluish species. Algological Studies 71: 13–21.
[14] Pueschel, C.M.; Saunders, G.W. & West, J.A. 2000. Affinities of the freshwater red alga Audouinella macrospora (Florideophyceae, Rhodophyta) and related forms based on SSU rRNA gene sequence analysis and pit plug ultrastructure. Journal of Phycology 36: 433-439. [ Links ]
[15] Shaw, D. 2013. Community and conservation in Bolivia´s Yacuma Pampas. The Ecologist, January 22nd: http://www.theecologist.org. Consultada abril 19, 2015.
[16] Sheath, R.G. 1984. The biology of freshwater red algae. In Round, F.E. & Chapman, D.J. [Eds.] Progress in Phycological Research, Vol. 3. BioPress Ltd, Bristol, UK, pp. 89–157.
[17] Sheath, R.G.; Vis, M.L. & Cole, K.M. 1994. Distribution and systematics of Batrachospermum (Batrachospermales, Rhodophyta) in North America. 5. Section Aristata. Phycologia 33: 404-414. [ Links ]
[18] Trainor, F.R. 1978. Introductory Phycology. John Wiley & Sons, USA. 525 pp. [ Links ]
[19] Van den Hoek, C., Mann, D.G. & Jahns, H.M. 1995. Algae. An introduction to Phycology. Cambridge University Press. 623 pp. [ Links ]

