










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkActa Nova
On-line version ISSN 1683-0789
RevActaNova. vol.6 no.3 Cochabamba Mar. 2014
ARTÍCULO CIENTÍFICO
Patrón de distribución y riqueza de especies de las mariposas de la tribu Satyrini (Nymphalidae: Satyrinae) en los bosques nublados de Cochabamba (Bolivia)
Species richness and distribution of butterfly tribe Satyrini in the cloud rainforest of Cochabamba (Bolivian)
Ariel Ángel Céspedes Llave[1], Marcelo Martin Aliaga Arrieta2, Daniela Aguirre Torres1, Pedro Castro1 & José Deyby Montenegro Siles1
1Museo de Historia Natural Alcide dOrbigny, Casilla 4324, Potosí N° 1458, Bolivia.
2 Casa de las mariposas Pilpintu wasi, pilpintuwasi@gmail.com. scotigera@gmail.com
Recibido: 1 de octubre 2013; Aceptado: 5 de enero 2014
Resumen: Se muestra los patrones de distribución de mariposas de las subtribus Euptychiina y Pronophilina (Nymphalidae: Satyrinae: Satyrini), en los bosques nublados tropicales de Cochabamba (Yunga). El análisis se realizó en base al gradiente elevacional desde los 700 a 3 700 m de altura, se utilizó 1 147 registros de mariposas de la tribu Satyrini. En función a la riqueza de especies y cada registro se trabajó con 55 especies de Euptychiina y 109 especies de Pronophilina, se calculó medidas de tendencia central, posteriormente se realizaron el análisis de regresión simple entre la riqueza de especies con la altura, precipitación, temperatura máxima y mínima. Los resultados muestran que la mayor riqueza de especies de Satyrini (1 800 m) presentan un patrón de distribución en forma de joroba (hump-shaped), e independientemente cada sub tribu presenta este similar patrón pero a diferente altura: Euptychiina (1 500 m) y Pronophilina (2 250 m). Y los análisis de regresión prueban que la elevación y precipitación tienen un efecto significativo sobre la distribución en forma de joroba en Satyrini. Este tipo de análisis son la clave para identificar áreas importantes de conservación en montañas, y nos proporciona criterios básicos e información sobre cuales con las causas de concentración de especies a determinadas alturas.
Palabras clave: distribución altitudinal, riqueza de especies, Satyrini
Abstract: We evaluated the distribution of subtribes Pronophilina y Euptychiina butterflies (Nymphalidae: Satyrinae: Satyrini) in tropical cloud forests (Yungas) of Cochabamba. The analysis is performed based on the elevational gradient from 700 to 3 700 m high, was used 1 147 records of the tribe Satyrini butterflies. Depending on the species richness and each record we worked with 55 species of Euptychiina and 109 species of Pronophilina, we calculated measures of central tendency, subsequently performed simple regression analysis between species richness with altitude, rainfall, temperature maximum and minimum. The results show that the highest species richness of Satyrini (1 800 m) exhibit a distribution pattern hump-shaped, and whether each subtribe presents this same pattern but at different heights: Euptychiina (1 500 m) and Pronophilina (2 250 m). And regression analyzes show that elevation and precipitation have a significant effect on the hump-shaped distribution in Satyrini. This type of analysis is the key to identifying important conservation areas in mountains, and provides basic criteria and information on the causes which species concentration at certain heights.
Key words: elevational distribution, species richness, Satyrini
1. Introducción
Los Andes Tropicales es considerado un hostpots de alta prioridad para la conservación de plantas y animales (Myers et al. 2000), por presentar importantes centros de diversidad biológica y endemismo (Soria & Kessler 2007; Young 2007). En este existen varios tipos de bosques, que se extienden desde los 800 a 3 500 metros, incluyendo los bosques nublados de montaña (yungas, ceja de monte), que cubren más de 500 000 km² entre Perú y Bolivia. Además se encuentran entre los bosques más ricos y diversos de la Tierra (Conservation International 2013).
En este sentido, Bolivia presenta ecosistemas exclusivos como los yungas dentro de los Andes Tropicales, que según Benavides (2011) con referencia a la fauna de mariposas está situada entre los cuatro países más ricos del mundo. Dado que, presenta diferentes zonas de vida, debido a su geomorfología de dominio montañoso que decrece en altura hacia el este, finalizando en una llanura, atrayendo el interés de realizar estudios del efecto del gradiente altitudinal sobre los patrones de distribución de la riqueza de mariposas. Durante la última década, en los yungas de Bolivia diferentes estudios han demostrado la relación de la alta riqueza de especies y endemismo con patrones distribución a nivel espacial y gradiente altitudinal, realizados en plantas (fanerógamas y criptógamas) (Soria & Kessler 2007) y animales (aves, anfibios y mamíferos) (Herzog et al. 2005; Young 2007).
Pero con respecto a los insectos como las mariposas es escasa la información que se tiene en Bolivia. Aunque hay que destacar la información que existe sobre los patrones de distribución en mariposas diurnas de la tribu Satyrini (Nymphalidae: Satyrinae), realizados en ecosistemas muy similares de Venezuela, Ecuador y Perú (Wells et al. 1985; Pyrcz& Viloria 1999, 2005; Pyrcz & Wojtusiak 2002; Pyrcz 2004; Pyrcz et al. 2009).
Las mariposas Satyrinae pertenecen al clado Satyroide (Garzon & Miranda 2005; Zhang et al. 2008), o como lo denomina Walhberg (2003, 2005) linaje Satyrine dentro la familia Nymphalidae. El cual está comprendido por tres subfamilias, Calinaginae, Charaxinae y Satyrinae, las dos últimas están presentes en el Neotropico. Con respecto a la subfamilia Satyrinae, está compuesto por nueve tribus, pero de interés según su distribución Neotropical son cuatro tribus: Morphini, Brassolini, Haeterini y Satyrini (Walhberg 2005).
En base a la sinopsis anterior, se hace un énfasis en la tribu Satyrini (compuesta por tres subtribus: Coenonymphina, Euptychiina y Pronophilina), ya que según Wells et al. (1985), en el Libro Rojo de los Invertebrados de la UICN (Invertebrate Red Data Book - UICN), incluyen a mariposas de la subtribu Pronophilina dentro del rango de Rara, aludiendo a que dentro este grupo taxonómico existíanespecies que son poco comunes y/o a los datos insuficientes que se tenía para entonces. Además, por la alta concentración de especies existentes por encima de los 2 300 m en los Andes tropicales, siendo el grupo más diverso de las mariposas en este tipo de ecosistemas.
Actualmente el hábitat donde vuelan estas mariposas se está degradando, debido a diferentes presiones (asentamientos ilegales e invasiones, agricultura extensiva, deforestación, etc.), que según se estima que existe una pérdida anual del 2,5 % de la cobertura vegetal natural en los ecosistemas de los yungas (Müller et al. 2003), lo cual está afectando en el decremento de esta diversidad de mariposas Satyrini, por tanto es necesario realizar acciones de conservación (Wells et al. 1985).
Con este trabajo se pretende contribuir con mayor información, al conocimiento en riqueza de especies y a nivel de distribución altitudinal, explicando como ocurre el patrón de distribución de las mariposas Satyrini en los yungas de Cochabamba (Bolivia). Además, realzar la importancia de conservación de estos ecosistemas importantes, bastante frágiles y ricos en especies como son los bosques nublados de montaña en el departamento de Cochabamba.
2. Metodología
2.1 Área de Estudio
El área de estudio comprende aproximadamente de 5 243 km2, los cuales corresponden a cinco transectos de muestreo realizados en los yungas del departamento de Cochabamba. Desde el sudeste del Parque Nacional Carrasco (PNC) (S 17,7, W 64,64) hasta el noroeste del Chapare (S 16,80, W 66,28). Tres transectos se encuentran dentro la jurisdicción del área protegida PNC (Pojo, Totora y Villa Tunari), una sobre la carretera troncal Cochabamba-Santa Cruz, (Sindicato Paracti) y el ultimo transecto en la jurisdicción de la Reserva Municipal de Flora y Fauna Norte de Tiquipaya (RMFFNT) (Fig 1). Aunque este último transecto se encuentra dentro los límites municipales del Municipio de Chapare, por cuestiones de proximidad y accesibilidad ahora al parecer pertenece al Municipio de Tiquipaya.
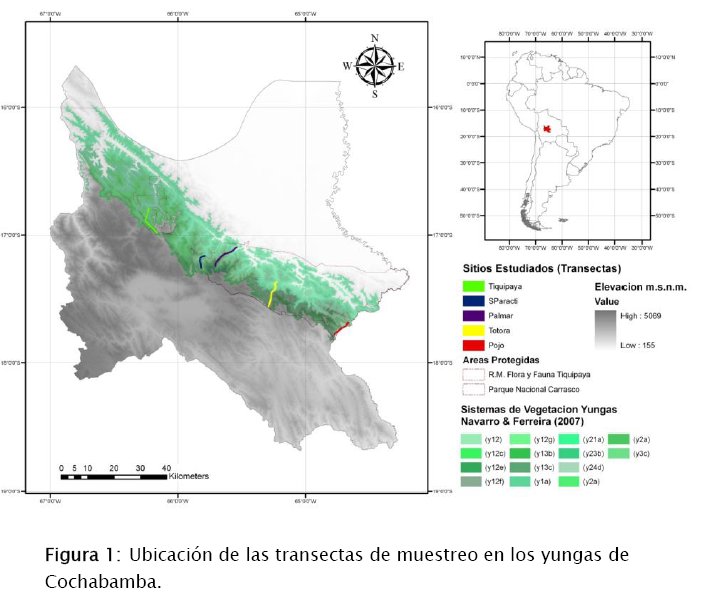
Los yungas de Cochabamba, según Navarro & Maldonado (2002) pertenece a los yungas subandinos pluviales, abarcando dos distritos biogeográficos: los yungas del Chapare (Sector Biogeográfico Yungas del Ichilo) y los yungas de Altamachi y Corani (Sector Biogeográfico Yungas del Beni). Ambos pertenecen a la formación vegetal, Macroserie de Nectandra laurel – Dyctiocaryum lamarckianum, descrito por Navarro et al. (2004). En estas dos series, se localizan 14 sistemas ecológicos de los Yungas (Navarro & Ferreira 2007), comprendido entre los 400 a 4 600 mde altura (ver figura 1). El bioclima es de tipo pluvial al sudeste y al noroeste tiende a ser mas pluvial estacional, además, los ombrotipos van desde hiper-húmedo a húmedo, es decir con una precipitación que va entre los 500 a 2 000 mm anuales (Navarro & Maldonado 2002).
2.2 Análisis de los datos
En el presente estudio se utilizó la base de mariposas de la tribu Satyrini, generado por el proyecto "Estado de conservación del linaje Satyrine (Nymphalidae) de la yunga de Cochabamba" realizado por Cespedes & Aliaga (2008) y datos complementarios de Aguirre (2004) y Aliaga & Cespedes (2007). Se consideró la información geográfica (en grados decimales)y altura, de una de las especies con datos espaciales georreferenciados identificados de las subtribus Euptychiina y Pronophilina. No se consideró a las especies subtribu Coenonymphina ya que solo contiene dos especies. De manera que se trabajó con 1147 registros de mariposas de la tribu Satyrini, específicamente 468 registros correspondientes a 55 especies de Euptychiina y de Pronophilina 679 registros que corresponden a 109 especies (Anexo 1). El registro taxonómico de la identificación de cada especie se estandarizo mediante la lista de especies (Gerardo's list.) de Lamas (2004) y Gareca et al. (2006).
Se trabajó con el número de observaciones que contenía cada una de las 164 especies, de los cuales se calculó medidas de tendencia central (la media, el desvió estándar y error estándar) a los datos de altitud. Posteriormente con las medidas de tendencia central se realizó un histograma de distribución de frecuencias para cada subtribu en función de la altura. Con ello se discrimino según la cantidad de observaciones, la cantidad es especies raras (poco frecuentes) y comunes (más frecuentes) y la cantidad de estas especies en porcentaje. Además, con los datos de distribución de frecuencia se realizó una comparación de las medias en base a boxplot generados para las dos tribus. La comparación se realizó con nivel de significancia al 95%, probando que la altura tiene un efecto en los patrones de distribución de las dos subtribus Euptychiina y Pronophilina.
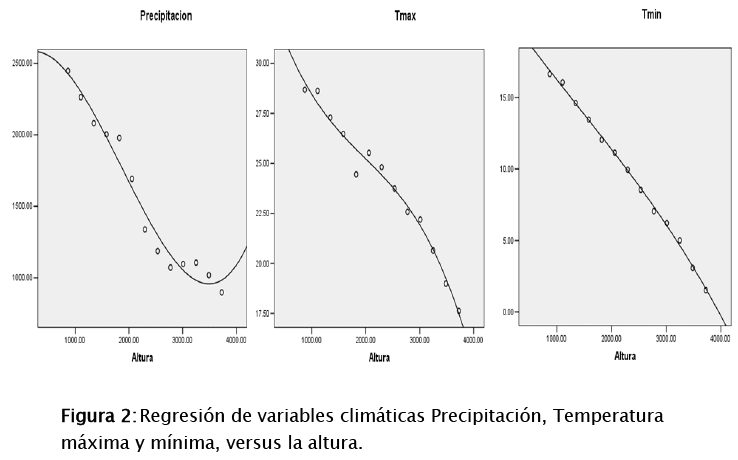
Se utilizó información climática de World Clim (Hijmans et al.2005) y los datos de altura obtenidos de un modelo de elevación digital a 30 m de resolución (Digital Elevation Model DEM) del Sistema Espacial de Japón (Japan Space Systems 2012). Con esta información se realizó correlaciones entre la altura y los datos de clima (precipitación y temperatura máxima y mínima) (Fig. 2), para luego evaluar posteriormente la relación de estos factores ambientales sobre la distribución de las mariposas.
3. Resultados
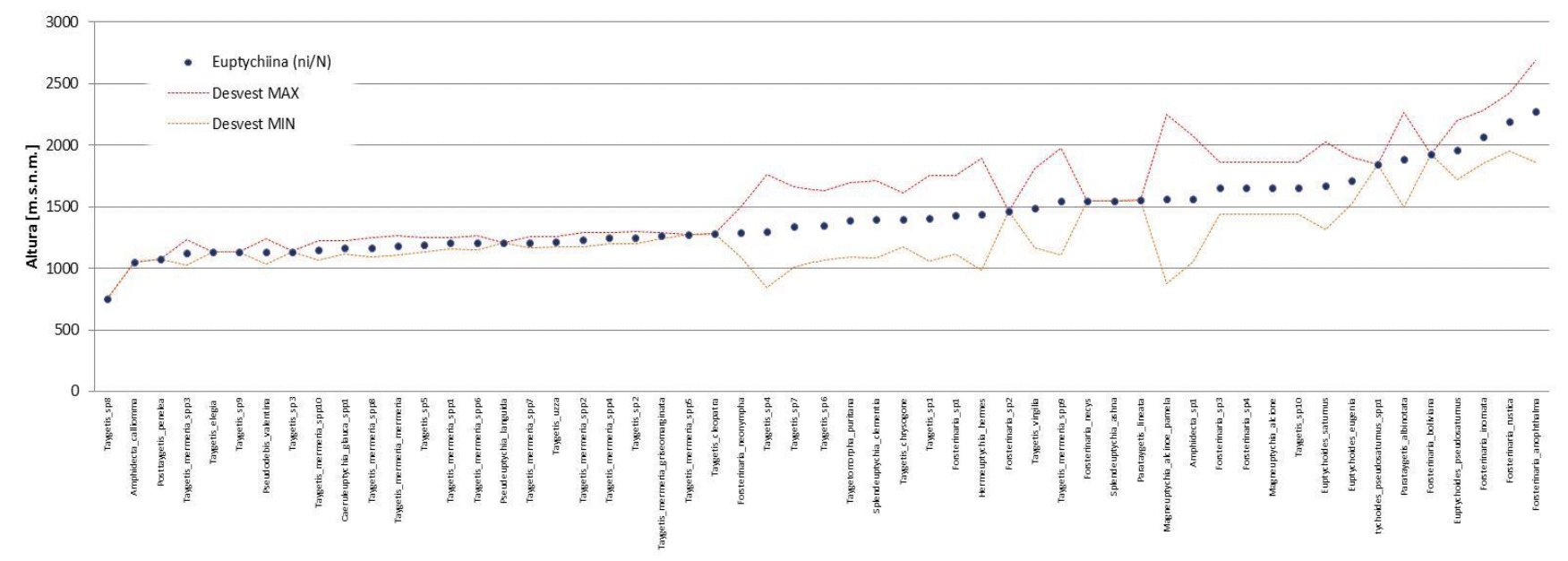
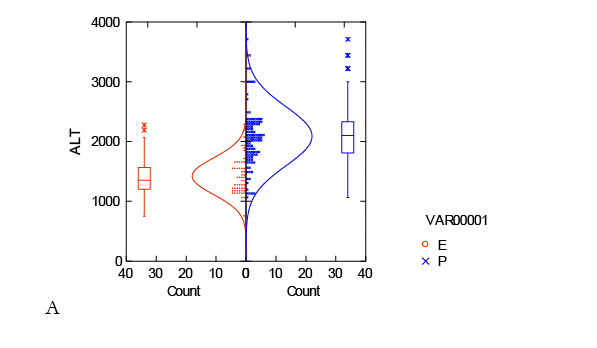
Los resultados obtenidos se tienen por separado para las dos subtribus evaluadas Euptychiina y Pronophilina. De manera que, con el cálculo de tendencia central para las 55 especies de Euptychiina, muestran una tendencia que abarca aproximadamente entre los 700 hasta los 2 500 m s.n.m. (Fig. 3).
En cambio para las 109 especies de Pronophilina muestran un rango de distribución diferente, entre los 1 000 hasta 3 800 m s.n.m (Fig. 4). Para Euptychiina la mayoría de los registros de distribución se encuentran entre los 1 000 a 2 000 m s.n.m. y Pronophilina entre 1 500 a 2 500 m s.n.m.
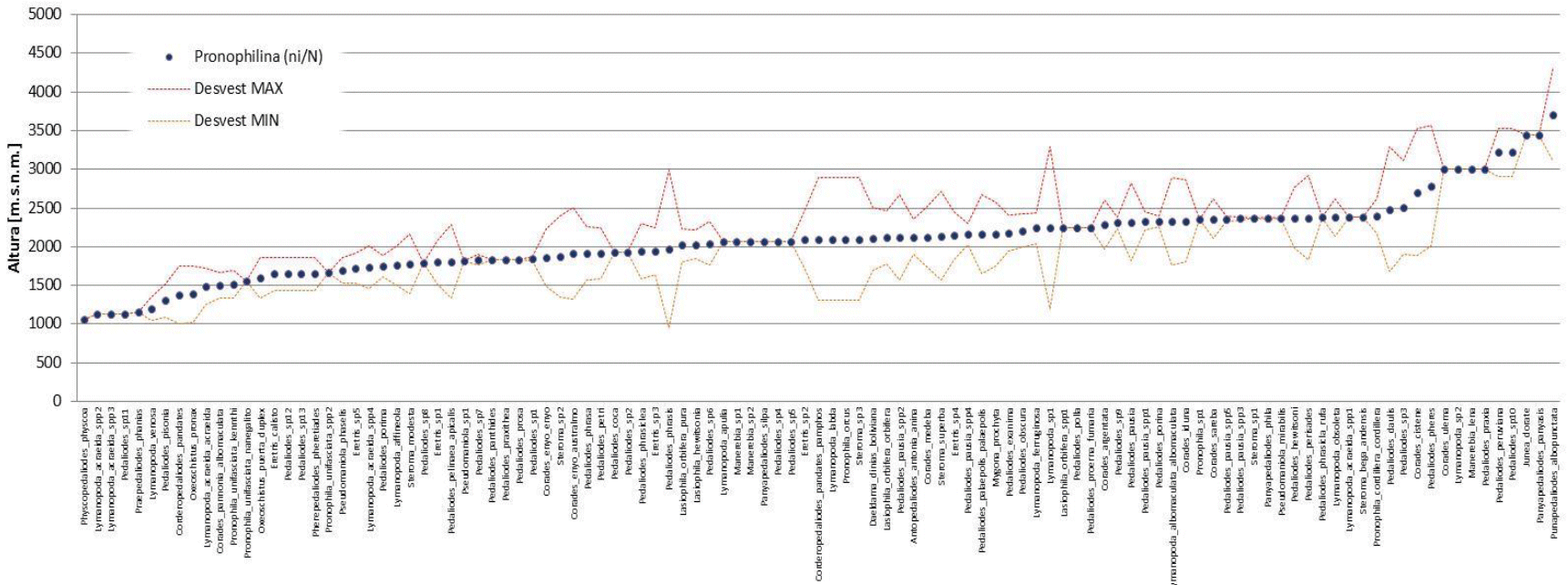
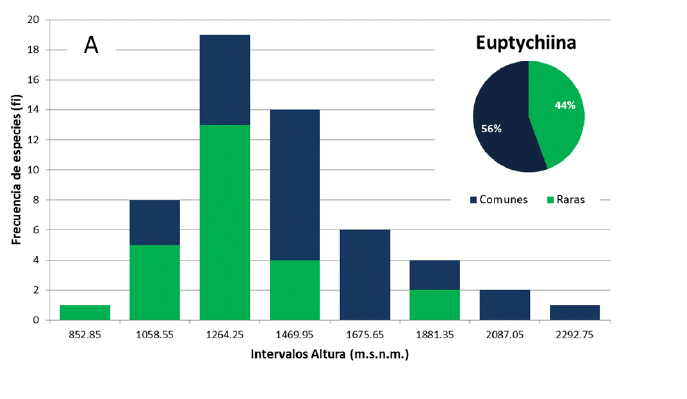
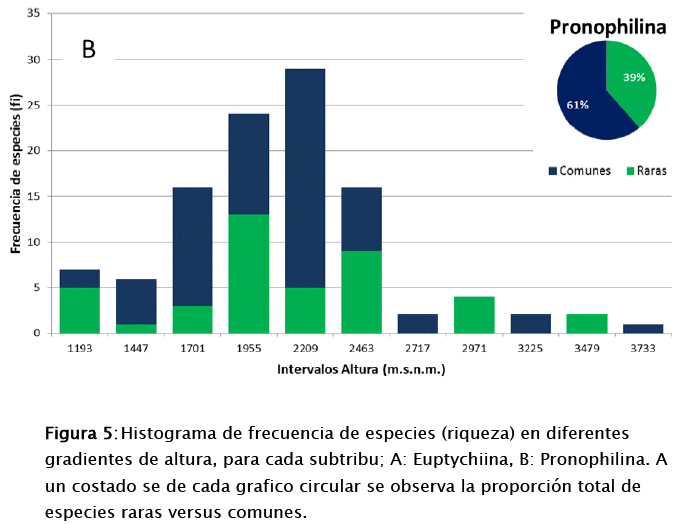
En base al registro en función la altura, existen especies que no presentan un rango amplio (desvió estándar) y otros que sí. De las 55 especies de Euptychiina, se pudo determinar que el 56% de las especies son comunes y un 44% son raras (Fig. 5a). Y para Pronophilina al respecto presenta un 61% son comunes y 39% son raras (Fig. 5b). Esta diferenciación de especies raras y comunes en ambas subtribus, denota que el número de especies comunes son más dominantes con respecto a las raras en los gradientes definidos (intervalo de altura). Además, la distribución de frecuencias posibilito identificar gradientes donde se concentra el mayor número de especies. Para Euptychiina se halló una riqueza de 19 especies a los 1 264 m s.n.m. y 10 especies a los 1 469 m s.n.m (siendo lo valores más altos). Pronophilina se observa dos picos altos de riqueza con 29 especies a los 2 209 m s.n.m. y 24 especies a los 1 955 m s.n.m.
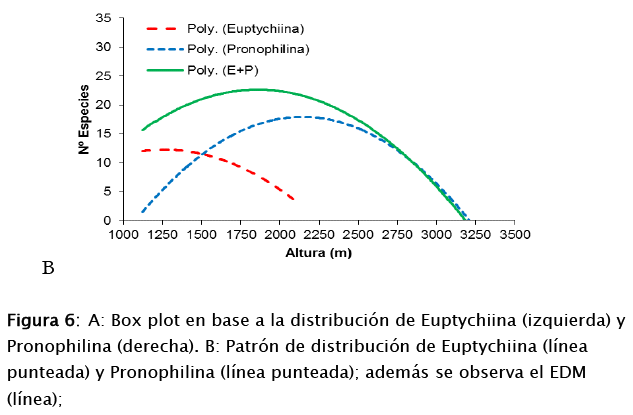
Las curvas de distribución de ambas subtribus presentan forma de joroba. Euptychiina tiene amplitud con mayor número de especies entre los 1 000 a 2 000 m s.n.m., y Pronophilina en cambio abarca la amplitud mayor de Euptychiina entre los 1 200 a 2 900 m s.n.m (Fig. 6a). Al sobreponerse ambas curvas considerando todas las especies, se pudo identificar otra curva en forma de joroba, donde la mayor concentración de número de especies es a los 1875 m s.n.m. (ver figura 6b).
Con la amplitud de distribución de ambas subtribus (E+P), se obtuvo correlaciones con las variables climáticas (precipitación, temperatura máxima y mínima). Del cual se observa que en función de un modelo cuadrático (ver figura 7), la precipitación es una variable tiene un efecto significativo (p=0,001; r2=0,77), con un 90% de confianza sobre la riqueza de especies. En segundo lugar la altura (como gradiente) tiene efecto significativo (p=0,03; r2=0,5), como lo demuestran los resultados de arriba. La temperatura mínima no tiene un efecto significativo (p=0,054; r2=0,44), de la misma manera la, la temperatura máxima no tuvo efecto (p=0,12; r2=0,36).
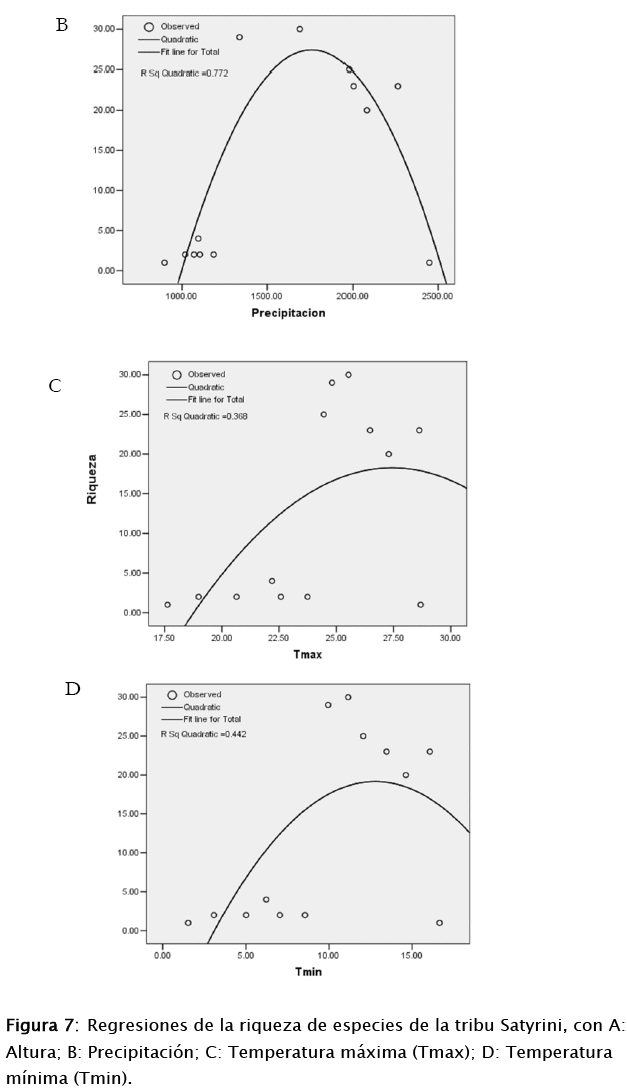
4. Análisis y discusión
La variación de la riqueza de especies a lo largo de gradientes altitudinales, es un tema central de interés biogeográfico y ecológico (Sanders et al. 2003, Gotelli et al. 2009). Se conoce que la riqueza de especies declina con incremento de la elevación (MacArthur 1972), sin embargo, en los bosques nublados de los Andes Tropicales se ha demostrado que la riqueza de especies tiende a mostrar otro patrón de distribución, en forma de joroba (hump-shaped), es decir, a elevaciones medias existe una mayor concentración de especies (Brehm et al. 2007). Estudios realizados en los bosques nublados tropicales con: Acanthaceae, Araceae, Bromeliaceae, Melastomataceae, Palmae, y Pteridophyta (Kessler 2001), en aves (Herzog et al. 1999; Katan & Franco 2004; Herzog et al. 2005), mariposas nocturnas Geometridae (Brehm & Fiedler 2003; Axmacher et al. 2004; Brehm & Fiedler 2004; Brehm et al. 2007)han demostrado estar presentes este tipo de distribución en forma de joroba.
Con respecto a las mariposas de la tribu Satyrini, en base a los resultados también presenta este mismo patrón de distribución donde a elevaciones medias existe una alta la riqueza de especies. Pyrcz (2009) también demostró que existe una congruencia muy notable en cuanto a los patrones de diversidad altitudinal para mariposas de Pronophilina, en varias áreas de la cordillera Andina del Ecuador y Venezuela, con rangos donde presentan una alta concentración de especies, aproximadamente entre los 2 300 a 2 900 m s.n.m (Pyrcz & Wojtusiak 2002; Pyrcz et al. 2009), similar resultados obtenidos en este trabajo. Con respecto a Euptychiina, se conoce que la ocurrencia de especies es mas en tierras bajas (Murray & Prowell 2005), pero esta subtribu también presenta una buena cantidad de especies en los yungas. Y posiblemente la riqueza de especies sea relativamente mayor o se equipare a las tierras bajas.
Los patrones de riqueza de especies a medias elevaciones puede diferir entre taxones o como dentro los taxones, y entre diferentes regiones o en la misma región (Rahbek 1995). Aunque, en áreas montañosas la elevación es un factor decisivo para los patrones de diversidad, porque presenta los cambios en la disponibilidad de recursos. Además, otro tipo de factores pueden afectar la distribución de la riqueza de especies en gradientes altitudinales, como los gradientes climáticos (temperatura, precipitación, humedad, etc.) (Brehm et al. 2007). Nuestros resultados demuestran que la precipitación es un factor que influye bastante. Aunque nosotros no realizamos una relación con la composición florística, estudios confirman que si existe bastante correspondencia entre la diversidad de fauna con la estructura de la vegetación en gradientes altitudinales (Jankowski et al. 2012, Joshi et al. 2012)
5. Conclusiones
La riqueza de especies es bastante alta, donde se ha podido caracterizar dos categorías, especies raras y comunes. Siendo la categoría de rara de prioridad por la posibilidad de haber identificado especies endémicas de rangos altitudinales. Como es el caso para Euptychiina con el 44% de especies raras que equivalen a 24 especies y para Pronophilina se han identificado 42 especies (39%).
Estas dos subtribus de tribu Satyrini, se encuentran distribuidas en diferentes rangos de distribución. Para la subtribu Euptychiina, es que dominan rangos inferiores en un gradiente altitudinal. En cambio Pronophilina, se encuentra dominando rangos altitudinales mayores, entre los 1 750 a 2 750 m s.n.m., rango donde se encuentra aproximadamente el 71% de las especies.
En base a los resultados mencionado anteriormente, el solapamiento de los patrones de distribución de las subtribus Euptychiina y Pronophilina, son definidos por la riqueza de especies presentes en cada rango, mostrando un patrón de distribución en forma de joroba. Dentro de ese rango, existen especies que tiene una amplitud de distribución ≤100 m, y otras que alcanzan desde ≤ 200 m hasta 1 000 m. La subtribu Euptychiina presenta un 60% de las especies tienen rangos entre 100 m y 500 m, y Pronophilina, el 62% de las especies, tienen una amplitud entre 100 m y 1 000 m. Este tipo de análisis son la clave para identificar áreas importantes de conservación en montañas, y nos proporciona criterios básicos e información sobre cuales con las causas de concentración de especies a determinadas alturas.
Agradecimientos
Agradecemos al Proyecto Diversidad de las Mariposas Tropicales (Tropical Andean Butterfly Diversity Project) y al programa de becas Werner Hannagarth de la Fundación PUMA, por haber contribuido al desarrollo de esta investigación desde sus inicios. Además al Museo de Historia Natural Alcide d'Orbigny de Cochabamba, por su apoyo incondicional. A Ricardo Sahonero & Gilberth Lachaume amigos que nos brindaron la oportunidad de comenzar en los caminos de la investigación.
Bibliografía
[1] AGUIRRE, Daniela. Estructura de la comunidad de cuatro familias de mariposas a lo largo de un gradiente altitudinal (correspondiente a cinco pisos bioclimaticos) en la cuenca oeste del río Ichilo, Cochabamba. Trabajo de Titulación (Licenciado Biologia) Cochabamba, Bolivia. Universidad Mayor de San Simón. 2004.
[2] ALIAGA, Marcelo. & CÉSPEDES, Ariel. Biogeografia de la tribu Pronophilini (Satyrinae) en los yungas del Parque nacional Carrasco (Bolivia). Informe Técnico de Proyecto. Bolivia. Tropical Andean Butterfly Diversity Project. 2007.
[3] Andrade-C., M.G. 1998. Utilización de las mariposas como bioindicadoras del tipo de hábitat y su biodiversidad en Colombia. Rev. Acad. Colomb. Cienc., 22(84): 407-421. [ Links ]
[4] Axmacher, J.C.; Hotmann, G.; Scheuermann, L.; Brehm, G.; Müller-Hohenstein K. & Fiedler, K. 2004. Diversity of geometrid moths (Lepidoptera: Geometridae) along an Afrotropical elevational rainforest transect. Diversity and Distributions, 10: 293-302. [ Links ]
[5] Benavides, J. 2011. El fascinante mundo de las mariposas. Fundación Amazonia de Pie. La Paz, Bolivia. 162 pp.
[6] Brehm, G.; Colwell, R.K. & Kluge, J. 2007. The role of environment and mid-domain effect on moth species richness along a tropical elevational gradient. Global Ecology and Biogeography, 2 (16): 205–219. [ Links ]
[7] Brehm, G. & Fiedler, K. 2004. Faunal composition of geometrid moths changes with altitude in an Andean montane rain forest. Journal of Biogeography, 30: 431-440. [ Links ]
[8] Brehm, G. & Fiedler, K. 2003. Bergmann´s rule does not apply to geometrid moths along an elevational gradient in an Andean montane rain forest. Global Ecology and Biogeography, 13: 7-14 [ Links ]
[9] CÉSPEDES, Ariel, & ALIAGA, Marcelo. Estado de Conservación del linaje Satyroide (Nymphalidae) de la yunga de Cochabamba (Bolivia). Informe Técnico de Proyecto. La Paz, Bolivia. Fundación PUMA. 2008.
[10] Conservation International (C.I.) 2013. Tropical Andes. Disponible en: http://www.conservation.org/. Fecha de consulta: 17 de septiembre del 2013.
[11] Garzón, O.I., & Miranda, E.D. 2005. Las relaciones filogenéticas dentro Nymphalidae usando experimentos basados en el análisis de sensibilidad. Laboratorio de Sistemática & Biogeografía, Universidad Industrial de Santander. Página web: http://tux.uis.edu.co/labsist. [ Links ]
[12] Gareca Y., Forno, E.; Pyrcz, T.; Willmott, K. & Reichle, S. 2006. Lista preliminar de mariposas diurnas de Bolivia. En: Gareca, Y. & S. Reichle (eds.) 2006. Mariposas diurnas de Bolivia. PROMETA. Santa Cruz – Bolivia. 26-31. [ Links ]
[13] Gotelli N.; Anderson, M.J.; Arita, H.T.; Chao, A.; Colwell, R.K.; Connolly, S.R.; Currie, D.J.; Dunn, R.R.; Graves, G.R.; Green, J.L.; Grytnes, J.; Jiang, Y.; Jetz, W.; Lyons, S.K.; McCain, C.M.; Magurran, A.E.; Rahbek, C.; Rangel, T.; Soberón, J.; Webb, C.O. & Willig, M.R. 2009. Patterns and causes of species richness: a general simulation model for macroecology. Ecol. Lett. 12, 873–886. [ Links ]
[14] Herzog, S. K.; Kessler M. & Bach, K. 2005. The elevational gradient in Andean bird species richness at the local scale: a foothill peak and a high-elevation plateau. Ecography 28:1–14. [ Links ]
[15] Hijmans, R.J., Cameron, S.E.; Parra, J.L.; Jones, P.G. & Jarvis, A. 2005. Very high resolution interpolated climate surfaces for global land areas. International Journal of Climatology 25: 1965-1978. [ Links ]
[16] Japan Space Systems. 2012. ASTER GDEM. Visitado el 21 de diciembre del 2010. Pagina web: http://www.jspacesystems.or.jp/ersdac/GDEM/E/ [ Links ]
[17] Jankowski, J.E.; Merkord, C.L.; Rios, W.F.; Cabrera, K.G.; Revilla, N.S. & Silman, M.R. 2013. The relationship of tropical bird communities to tree species composition and vegetation structure along an Andean elevational gradient. Journal of Biogeography, 40: 950–962. [ Links ]
[18] Joshi, K.K.; Bhatt, D. & Thapliyal, A. 2012. Avian diversity and its association with vegetation structure in different elevational zones of Nainital district (Western Himalayan) of Uttarakhand. International Journal of Biodiversity and Conservation, 4(11): 364-376. [ Links ]
[19] Katan, G. H., & Franco, P. 2004. Bird diversity along elevational gradients in the Andes of Colombia: area and mass effects. Global Ecology and Biogeography 13: 451-458 [ Links ]
[20] Kessler, M. 2001. Patterns of diversity and range size of selected plant groups along an elevational transect in the Bolivian Andes. Biodiversity and Conservation, 10: 1897-1921. [ Links ]
[21] Lamas, G. 2004. Atlas of Neotropical Lepidoptera. Checklist: Part 4A Hesperoidea – Papilionoidea. Association for Tropical Lepidoptera. EEUU.
[22] MacArthur, R.H. 1972. Geographical Ecology. New York: Harper & Row [ Links ]
[23] Müller R., Briancon, A.; Hinojosa, I. & Ergueta, P. 2003. Prioridades de conservación en los Yungas Bolivianos. Editorial F.A.N., Santa Cruz de la Sierra, Bolivia.
[24] Murray, D. & Prowell, D.P. 2005. Molecular phylogenetics and evolutionary history of the neotropical satyrine subtribe Euptychiina (Nymphalidae: Satyrinae). Molecular Phylogenetics & Evolution, 34: 67-80. [ Links ]
[25] Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.B. & Kent, J. 2000. Biodiversity hotspots for conservation priorities. Nature, 203: 853–858. [ Links ]
[26] Navarro, G., & Ferreira, W. 2007. Mapa de Vegetación de Bolivia. The Nature Conservancy. Bolivia. [ Links ]
[27] Navarro, G.; Ferreira, W.; Antezana, C.; Arrázola, S. & Vargas, R. 2004. Bio-Corredor Amboró Madidi, Zonificación Ecológica. Ed. FAN. Santa Cruz. Bolivia.
[28] Navarro, G., & Maldonado, M. 2002. Geografía ecológica de Bolivia: Vegetación y ambientes acuáticos. Ed. Fundación Simon I. Patiño. Bolivia. pp. 278-342. [ Links ]
[29] Pyrcz, T. W. & Viloria, A.L. 1999. Contribution to the knowledge of Ecuadorian Pronophilini, Part 1; new pedaliodines (Lepidoptera: Nymphalidae: Satyrinae). Genus, 10 (1): 117-150. [ Links ]
[30] Pyrcz, T.W. & Wojtusiak, J. 2002. The vertical distribution of Pronophiline butterflies (Nymphalidae, Satyrinae) along an elevational transect in Monte Zerpa (Cordillera de Merida, Venezuela) with remarks on their diversity and parapatric distribution. Global Ecology & Biogeography, 11:211-221. [ Links ]
[31] Pyrcz, T W. 2004. Pronophiline butterflies of the highlands of Chachapoyas in northern Peru: faunal survey, diversity and distribution patterns (Lepidoptera, Nymphalidae, Satyrinae). Genus, 15 (4): 455-622. [ Links ]
[32] Pyrcz, T.W. & Viloria, A.L. 2005. Adiciones a la fauna de mariposas de la reserva forestal de tambito (colombia): descripción de cuatro subespecies nuevas de la subtribu Pronophilina (Nymphalidae, Satyrinae). Boletín Científico - Centro de Museos - Museo de Historia Natural, 9: 258-277. [ Links ]
[33] Pyrcz, T.W.; Wojtusiak J. & Garlacz, R. 2009. Diversity and Distribution Patterns of Pronophilina Butterflies (Lepidoptera: Nymphalidae: Satyrinae) along an Altitudinal Transect in North-Western Ecuador. Neotropical Entomology, 38(6):716-726. [ Links ]
[34] Rahbek, C. 1995. The elevational gradient of species richness – a uniform pattern. Ecography, 18: 200–205. [ Links ]
[35] Sanders, N. J. 2002. Elevational gradients in ant species richness: area, geometry, and Rapoport´s rule. Ecography, 25: 25-32 [ Links ]
[36] Soria, A.R.W. & Kessler, M. 2007. The influence of sampling intensity on the perception of the spatial distribution of tropical diversity and endemism: a case study of ferns from Bolivia. Diversity and Distributions, Journal compilation Blackwell Publishing Ltd.: 1- 8. [ Links ]
[37] Sreekumar, P.G. & Balakrishnan, M. 2001. Habitat and altitude preferences of butterflies in Aralam Wildlife Sanctuary, Kerala. Tropical Ecology, 42(2): 277-281. [ Links ]
[38] Wahlberg, N.; Braby, M.F.; Brower, A.V.Z.; de Jong, R.; Lee, M.M.; Nylin, S.; Pierce, N.E.; Sperling, F.A.H.; Vila, R.; Warren, A.D. & Zakharov. E. 2005. Synergistic effects of combining Morphological and molecular data in resolving the phylogeny of butterflies and skippers. Proceedings of the Royal Society of London, 272: 1577–1586. [ Links ]
[39] Wahlberg, N.; Weingartner, E. & Nylin, S. 2003. Towards a better understanding of the higher systematics of Nymphalidae (Lepidoptera: Papilionoidea). Molecular Phylogenetics and Evolution, 28: 473–487. [ Links ]
[40] Wallis DeVries, M.F. & Raemakers, I. 2001. Does extensive grazing benefit butterflies in coastal dunes? Restoration Ecology, 9:179– 188. [ Links ]
[41] Wells, S.; Pyle, R. & Collins, M. 1985. The IUCN Invertebrate Red Data Book. Ed. IUCN Gland Switzerland. Unwin Brothers Limited. Cambridge U.K. [ Links ]
[42] Young, B. 2007. Distribución de las especies endémicas en la vertiente oriental de los Andes en Perú y Bolivia. Nature Serve, Arlington, Virginia, EE UU. 90 pp. [ Links ]
[43] Zhang M.; Cao, T.; Jin, K.; Ren, Z.; Guo, Y.; Shi, J.; Zhong, Y. & Ma, E. 2008. Estimating divergence times among subfamilies in Nymphalidae. Chinese Science Bulletin, 53 (17): 2652-2658. [ Links ]
Lista de especies de la tribu Satyrini (Nymphalidae) identificados en los yungas de Cochabamba.
Notas
[1] Ariel Cespedes, scotigera@gmail.com

