










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkActa Nova
versión On-line ISSN 1683-0789
RevActaNova. v.6 n.1-2 Cochabamba mar. 2013
ARTÍCULO CIENTÍFICO
Ocurrencia de Cyclotella meduanae Germain (Bacillariophyta) en la laguna Alalay, un cuerpo de agua salobre y eutrófico (Cochabamba, Bolivia)
On the occurrence of Cyclotella meduanae Germain (Bacillariophyta) in Alalay Pond, a saline and eutrophic water body (Cochabamba, Bolivia)
Eduardo A. Morales1, Sinziana F. Rivera1, Rosemary Ayala2 & Francisca Acosta2
1Herbario Criptogámico, Universidad Católica Boliviana San Pablo. Calle M. Márquez esq. Plaza Jorge Trigo s/n, P.O. Box 5381, Cochabamba, Bolivia
2Unidad de Limnología y Recursos Acuáticos (ULRA), Universidad Mayor de San Simón. P.O. Box 471, Cochabamba, Bolivia
Recibido: 17 de agosto 2011; Aceptado: 19 diciembre 2011
Resumen: Se reporta la ocurrencia en el año 2005 de la diatomea céntrica Cyclotella meduanae Germain en la laguna Alalay, un ecosistema salobre y altamente eutrofizado dentro de la ciudad de Cochabamba, Bolivia. Los tamaños naturalmente pequeños de las células de esta diatomea no permiten su identificación confiable a los aumentos proporcionados por la microscopía fotónica, haciéndose necesaria la utilización de microscopía electrónica de barrido (MEB) para la visualización de las características diagnósticas que la diferencian de taxones morfológicamente similares: cara valvar con el área central casi plana y sin procesos reforzados; el perímetro y zona marginal de la cara valvar ondulados, procesos reforzados con tres poros satélite y estrías no alveoladas. Se discuten registros previos en Sudamérica y la ecología de la especie en función de la literatura disponible.
Palabras clave: Bacillariophyta, Bolivia, diatomeas céntricas, distribución, laguna Alalay, Sudamérica, taxonomía
Abstract: The occurrence in year 2005 of the centric diatom Cyclotella meduanae Germain in Alalay Pond, a saline, highly eutrophic ecosystem in the city of Cochabamba, Bolivia is reported. The naturally small sizes attained by the cells of this taxon do not allow their reliable identification at magnifications given by the light microscope. Therefore, the use of scanning electron microscopy (SEM) is required to observe the diagnostic features that separate C. meduanae from morphologically similar taxa: the relatively flat central area lacking strutted processes, the undulated valve face perimeter, presence of strutted processes with three satellite pores and presence of non-alveolate striae. Previous South American records of the taxon are discussed, as is its ecology in the context of available literature.
Keywords: Alalay Pond, Bacillariophyta, Bolivia, centric diatom, distribution, South America, taxonomy
1. Introducción
Cyclotella meduanae Germain es una pequeña diatomea céntrica que ha sido reportada en Sudamérica sólo recientemente [9]. Originalmente, el taxón fue descrito por Germain a partir de muestras del río Mayenne, Francia, y lo mencionó también para el río Maine, un tributario del río Loire [11]. Posteriormente, esta diatomea ha sido reportada en varias regiones geográficas del mundo, incluyendo Alemania [17], Austria [17], España [7][17][26][28], Hungría [17][18], Gran Bretaña [32], Holanda [17], Japón [22][24][30], Canadá [14], Estados Unidos [2] y ahora Brasil [9].
Es posible que haya sido confundida con otros taxa morfológicamente similares y que co-ocurren con ella, tales como Cyclotella atomus Hust., Cyclotella katiana Sala & Ramirez y Cyclotella meneghiniana Kütz. y por esta razón no haya sido reportada más amplia y frecuentemente en cada continente. Como fue expuesto por Cavalcante et al. [9], para la identificación de C. meduanae es necesario utilizar microscopía electrónica debido a que los caracteres diagnósticos sólo pueden ser observados a grandes aumentos. A nivel de microscopía fotónica, estas características no son claramente perceptibles, facilitando la confusión.
Se presenta aquí un análisis de MEB de muestras de la laguna Alalay colectadas dentro del marco del proyecto MANLAKE de la Unidad de Limnología y Recursos Acuáticos, Universidad Mayor de San Simón, Cochabamba. Este proyecto multidisciplinario tuvo como objetivo generar herramientas que contribuyeran al manejo de lagunas interandinas eutrofizadas, siendo la más importante de ellas la laguna Alalay por su valor paisajístico, su extensión y la diversidad de aves acuáticas que llegan o habitan en esta laguna [1][4].
Discutimos la morfología y ecología de este taxón en función de la literatura disponible.
2. Metodología
La laguna Alalay (17º 23' 43'' S, 66º 09' 35'' O) tiene una extensión de 230 ha, una profundidad de 0,5 a 3 m y una altitud de 2.560 m s.n.m. Es el cuerpo de agua léntico de agua dulce más grande en la ecorregión, conocida como el Valle Seco Interandino Sudamericano, que está caracterizada por una época cálida y lluviosa (de Octubre a Abril) y otra fría y seca (de Mayo a Septiembre). Topográficamente, la ecorregión presenta altitudes que oscilan entre 1.200-3.500 m s.n.m. y está flanqueada por laderas escarpadas y áreas de sedimentación aluvial de numerosos ríos intermitentes alimentados por aguas de montaña. En el departamento de Cochabamba, los Valles Secos Interandinos tienen temperaturas medias de 5 a 24° C en la época seca y de 12 a 24° C en la época lluviosa [19][23].
La vegetación actual de la laguna está compuesta principalmente por macrófitas emergentes (Schoenoplectus californicus subsp. tatora (Kunth) T. Koyama y Typha dominguensis Pers., sumergidas (Myriophyllum verticillatum L. y Potamogeton pectinatus L.) y flotantes (Azolla filiculoides Lam. y Pistia stratiotes L.) [5][8][10].
En aproximadamente las últimas cuatro décadas, la laguna ha presentado un estado eutrófico y contaminado debido a descargas domésticas, de fábricas de plásticos y ropa, así como a escorrentía cargada de desechos orgánicos [31]. La laguna también recibe grandes descargas de nutrientes, materia orgánica y desechos industriales de curtiembres y lavaderos de automóviles a través del río Rocha, el principal sistema lótico que atraviesa la ciudad de Cochabamba [6][19]. Estudios recientes muestran que las descargas de agua hacia la laguna contienen trazas de metales pesados, principalmente cobre, cromo, mercurio, plomo y zinc [25]. A pesar de esto, la laguna presenta una alta diversidad de macrófitas, algas, insectos, aves y algunos anfibios y reptiles, lo que la convierte en uno de los últimos ecosistemas de su tipo en los Valles Secos Interandinos [3][8][20][21][27].
Para el análisis de MEB, las muestras de fitoplancton fueron colectadas una vez de cada mesocosmos en Marzo de 2006, entre las 14 y 16 horas. Se extrajeron 20 L de muestra mediante un tubo colector de plancton. De este volumen, 500 mL fueron destinados al análisis de fitoplancton y 150 mL fueron filtrados mediante un equipo de filtración al vacío, utilizando filtros Watman GF/F; los que fueron luego debidamente preservados a temperaturas menores a -1º C. Los filtros fueron luego digeridos en 20 mL de ácido nítrico 70%, llevando a ebullición sobre una hornilla tipo hotplate por 20 min. Siete enjuagues con agua destilada y decantación después de 5 h de reposo produjeron suspensiones finales neutras que contenían las paredes celulares de diatomeas libres de materia orgánica. Aproximadamente 2 a 3 mL de cada muestra oxidada se dejaron secar a temperatura ambiente sobre pedazos de papel aluminio, pequeños retazos de los cuales se montaron sobre tacos metálicos utilizando cinta adhesiva de doble cara. Los tacos fueron luego recubiertos con una capa de oro-paladio de aproximadamente 135 Å de espesor, utilizando un metalizador Polaron E5100 por 1 min a 1,8 kV. Para las observaciones se utilizó un microscopio electrónico de barrido de emisión de campo Leo-Zeiss 982DSM Gemini, cuyas imágenes fueron directamente capturadas en forma digital y luego montadas con el programa Adobe Photoshop CS3 para la construcción de láminas que mostraran las características morfológicas principales de C. meduanae. La terminología morfológica sigue a Houk [15] y Houk & Klee [16].
Al tiempo de colecta de las muestras biológicas, se midieron in situ transparencia de Secchi (con un disco de Secchi), conductividad, pH, y oxígeno disuelto (OD) (con un medidor multiparámetro portátil) en la laguna.
3. Resultados y Discusión
Durante Marzo de 2006, la laguna Alalay presentó una transparencia de 23 cm, conductividad de 1.019,50 µS cm-1 y valores de pH y OD de 8,2 y 3,84 mg O2 L-1, respectivamente. La concentración de nutrientes en la laguna durante ese mes fue de 0,44 mg P L-1 de fósforo total y de 0,13 mg NO3 L-1 de nitratos [6], que evidencian el estado eutrófico de la laguna. No se tienen datos de salinidad medida como tal, ni de cloruros para el año 2006. Sin embargo, sí se tienen datos de cloruros de Junio de 2013, los cuales muestran valores altos que oscilan entre 300 a 330 mg Cl L-1. La alcalinidad total en este mes es también alta y se halla entre 524 y 689 mg CaCO3 L-1.
Cyclotella meduanae fue registrada como Cyclotella sp. durante los análisis de fitoplancton y su ocurrencia en el periodo 2003-2007 se restringió a una sola muestra del año 2005 colectada en un mesocosmos sometido a un tratamiento que no incluía peces. La densidad del taxón en esta muestra fue baja (83 células L-1) (Ayala et al., en prep.). Es posible que densidades menores se hayan producido en este u otro mesocosmos durante los experimentos de biomanipulación de la Laguna Alalay dentro del proyecto MANLAKE [1][5][6] pero debido a los bajos aumentos del microscopio fotónico usado en estos análisis, el taxón pudo haber pasado inadvertido. En los años 2011-2013 los estudios de diatomeas de bentos y plancton en Alalay han sido intensivos (p.e., Morales & Rivera [20]) y se analizaron varias decenas de muestras de ambas comunidades analizadas en detalle con MEB, pero C. meduanae no fue hallada nuevamente. Las razones por las cuales la especie apareció una sola vez y luego desapareció completamente son desconocidas, así como lo es su procedencia y la manera en la que llegó a Alalay.
El MEB muestra que los frústulos de C. meduanae son cilíndricos y de poca profundidad (alrededor de 2 µm) (Fig. 1C). Se observaron cadenas cortas de 2 a 3 células (Fig. 1C). Las valvas son circulares y miden entre 5 y 7 µm de diámetro (Figs 1A, 1D). El área central de la valva es casi plana tanto en vista externa como interna (Figs 1A, 1D). El perímetro y la zona marginal de la cara valvar son ondulados, con las estrías localizadas en las partes elevadas y las costillas en las depresiones y desprovistas de otras estructuras (Figs 1A, 1B). Las costillas son mucho más angostas en comparación con las estrías y pueden estar ligeramente elevadas en vista interna (Fig. 1E). Las estrías tienen más o menos la misma longitud desde el área central hasta la unión entre la cara valvar y el manto y continúan sin interrupción por el manto terminando cerca de su borde (Figs 1A-C). Estas estrías no son alveoladas y están compuestas externamente por varias filas desordenadas de areolas (Figs 1A-F), ocluidas internamente por una sola placa cribosa. La densidad de estrías varía entre 12 y 14 en 10 µm. Los procesos reforzados (fultoportulae) están localizados cada segunda o tercera costilla y ubicados en la zona de transición entre la cara y el manto valvar (Figs 1D, 1E). Cada uno de estos procesos tiene tres poros satélite (Figs 1D, 1E) y se abre externamente mediante un tubo muy corto (Figs 1B, 1C). En la zona central de la valva no hay procesos reforzados (Figs 1A, 1D). El único proceso labiado se halla en el anillo de procesos reforzados (Fig. 1D). Este proceso tiene labios muy pequeños y un pedicelo corto (recuadro en Fig. 1D). Externamente, se abre mediante un tubo más estrecho que los de los procesos reforzados (no mostrado aquí). No existen espinas pero sí pequeñas protuberancias en el manto valvar que resultan de la elevación de los bordes de areolas contiguas (Figs 1A-C). Las bandas conectivas son abiertas (Fig. 1F) y están perforadas por diminutos poros desordenados (Fig. 1C).
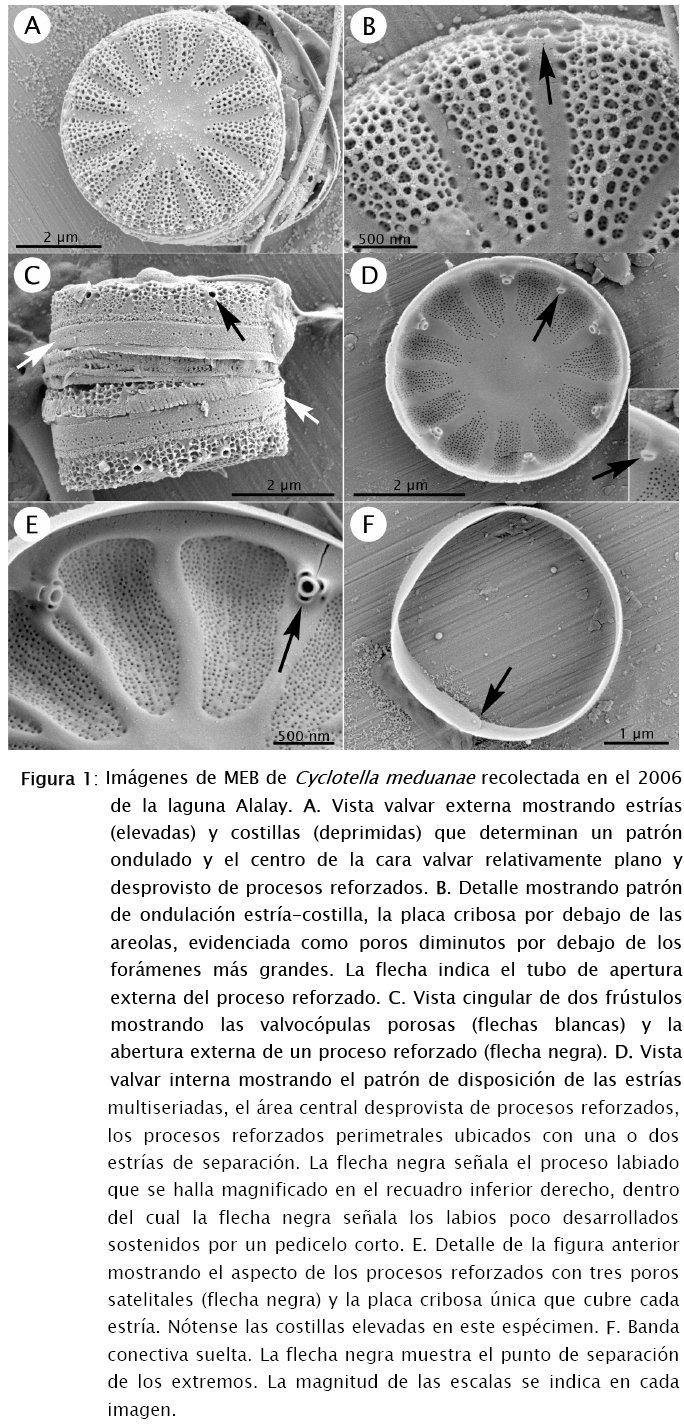
Las características que diferencian a C. meduanae de C. atomus, C. katiana y C. meneghiniana son el área central casi plana, el perímetro de la cara valvar ondulado, la presencia de procesos reforzados con tres poros satélite, las estrías areoladas, no alveoladas, y el área central desprovista de procesos reforzados [11][17][18][29].
El diámetro valvar y el número de estrías en los especímenes de Alalay está dentro de los rangos indicados para el material tipo de Francia (5-7 µm y 12-17 en 10 µm, respectivamente). Las características de los procesos reforzados y labiados son similares a los que figuran en la literatura, excepto los de la población japonesa estudiada por Tanaka [30] en la que el proceso labiado presenta labios mucho más desarrollados y un pedicelo más grande. Sin embargo, en estos especímenes japoneses, las costillas también están más desarrolladas y elevadas en vista interna y las estrías parecen ser más gruesas, lo que resulta en su densidad ligeramente más baja (10-12 en 10 µm). Es posible que las diferencias en el grado de silicificación entre la población de Alalay y la japonesa se deban a diferencias en las condiciones químicas del agua pero esto requiere una mayor investigación. Procesos labiados más desarrollados también están presentes en células de especímenes canadienses [14] aunque la identidad de estos individuos todavía no ha sido establecida, ya que en el resumen y las leyendas de las figuras 23 y 24 en Håkansson & Kling [14], pp. 289 y 291], aparece como Cyclotella meduanae, pero en sus observaciones, las autoras se refieren al mismo taxón como Cyclotella cf. meduanae, argumentando que existían leves diferencias con el material descrito por Germain [11].
Así como ocurrió con los ejemplares japoneses, la densidad de estrías también es baja tanto en los de Brasil (Cavalcante et al. [9]; 8-13 en 10 µm) como en las poblaciones de Alemania y España estudiadas por Houk et al. [17] (8-10 en 10 µm). Al no existir diferencias notorias en la combinación de las características diagnósticas mencionadas anteriormente, se puede sostener que las poblaciones de Alalay, Brasil, Japón, España y Alemania son conspecíficas, a pesar de la ligera diferencia de densidad de estrías y las características del proceso labiado. Cabe también mencionar que ninguna de esas poblaciones tiene espinas, estructuras que fueron erróneamente mencionadas como presentes en C. meduanae por Cavalcante et al. [9], p. 243]. Lo que estos autores muestran en sus figuras 7L-N corresponden en realidad a lo que Kiss et al. [18] denominan espínulas y gránulos, que son ontogenética y estructuralmente diferentes de las espinas.
Contrariamente a lo sostenido por Håkansson [13], Håkansson & Kling [14] y Sala & Ramírez [29], nosotros afirmamos que C. meduanae es un taxón fácilmente distinguible de otros morfológicamente similares, siempre y cuando se disponga de imágenes de MEB de alta resolución. De las características combinadas mencionadas con anterioridad, la presencia de procesos reforzados con tres poros satélites y la ausencia de estrías alveoladas deben ser determinados utilizando esta tecnología mientras que el área central casi plana, el perímetro ondulado de la cara valvar y la ausencia de procesos reforzados en el área central pueden (discutiblemente) ser observados con microscopios fotónicos bien calibrados.
La mención de que existen poblaciones japonesas de C. meduanae con procesos reforzados centrales (Ogawa [24]; discutida por Tanaka [30]), probablemente corresponde a la aplicación de un concepto demasiado amplio de este taxón. La evidencia en la literatura citada en el presente trabajo muestra que la metapoblación de C. meduanae posee una combinación de caracteres diagnósticos que varía muy poco y que las variaciones están más bien presentes en otros atributos como la densidad de estrías y las características estructurales de los procesos labiados. Por tanto, es muy posible que los individuos con procesos reforzados centrales mencionados por Ogawa [24] pertenezcan más bien a un taxón similar que se deberá determinar a través de un re-estudio del material analizado por esta autora.
Hasta el presente reporte, la ocurrencia de C. meduanae en Sudamérica parece haber estado restringida a Brasil. Previamente, existe un único reporte para el río de La Plata, Argentina [12] pero que necesitaría ser verificado (N. Maidana, 2013, com. pers.). Contrario a lo aseverado por Cavalcante et al. [9], Cyclotella meduanae no ha sido reportada para Colombia. El presente trabajo constituye, entonces, el segundo reporte verificado en el continente de esta diatomea céntrica pero, como se dijo anteriormente, es muy posible que haya sido hallada previamente y mencionada bajo distintos nombres debido a las dificultades que existen en su identificación.
Tomando en cuenta la información publicada sobre C. meduanae, resulta difícil determinar las razones de su distribución actual, es decir, no se puede concluir si esta distribución es natural o si es el producto de una facilitación por el incremento en el número de ecosistemas eutrofizados a nivel mundial. Asimismo, ante la ausencia de registros a largo plazo, no se puede determinar si su ocurrencia en Sudamérica es reciente o si ya estaba presente en este continente desde hace miles de años. Es importante hacer notar, sin embargo, que en Brasil [9] y Bolivia el taxón se halló en ambientes eutróficos, como ocurrió en varias localidades europeas y asiáticas. Con la degradación paulatina que están sufriendo los ecosistemas acuáticos a nivel mundial, es posible que se establezcan nuevas oportunidades para C. meduanae, especialmente en ecosistemas salobres. Esta observación se fundamenta además en el hecho de que el grueso de los reportes de la especie se hizo en las últimas dos décadas y para ecosistemas afectados en mayor o menor grado por actividad antrópica.
Kiss et al. [18] consideran que C. meduanae es típicamente potamoplánctica y que generalmente se desarrolla mejor en trechos de ríos con corrientes bajas. En el caso de Alalay, existe un movimiento constante de masas de agua inducido por vientos diariamente recurrentes, especialmente en horas de la tarde, y es posible que esto haya favorecido el desarrollo de C. meduanae en el 2006, aunque no existen datos que muestren que este año fue particularmente diferente en estas condiciones a años anteriores o posteriores. Con respecto a la salinidad, el río Mayenne, la localidad tipo donde la abundancia de C. meduanae fue alta [11] y el río Maine tienen tenores salinos relativamente elevados pero no como para ser considerados como salobres. Kiss et al. [18] aseveran que en ríos de Hungría donde se halló la especie el tenor salino es variable, por lo que este factor no parece influir sobre la distribución de C. meduanae. Especialmente en Japón, se han reportado altas densidades producidas en condiciones eutróficas o hipertróficas y con alcalinidades también altas [24][30].
En las aguas salobres de Alalay, las abundancias de C. meduanae no fueron muy altas (como se vió anteriormente) posiblemente debido al alto tenor salino.
Con relación al estado trófico, puede que los factores de alta disponibilidad de fósforo, nitrógeno y elevado pH en Alalay hayan favorecido la aparición aislada de C. meduanae.
4. Conclusiones
Cyclotella meduanae es un taxón con valvas muy pequeñas que sólo puede ser identificado con certitud bajo MEB. Las especies morfológicamente más cercanas a C. meduanae son C. atomus, C. katiana y C. meneghiniana, de las cuales se diferencia por la combinación del área central casi plana, el perímetro ondulado de la cara y margen valvar, la presencia de procesos reforzados con tres poros satélites, de estrías no alveoladas y el área central desprovista de procesos reforzados. La ocurrencia de C. meduanae en Bolivia se resume a una sola muestra colectada en el 2005 de mesocosmos construidos dentro de la laguna Alalay, pero es posible que haya pasado inadvertida porque la mayoría de los análisis del fitoplancton se realizaron a bajos aumentos de microscopía fotónica. Sin embargo, en estudios recientes con MEB de las comunidades del bentos y fitoplancton tampoco se ha encontrado esta especie. La única aparición de C. meduanae en Alalay puede haberse debido a las condiciones eutróficas de la laguna combinadas con algún factor como la corriente y otro desconocido en los mesocosmos mientras que la alta salinidad podría haber sido el factor que en definitiva no permitió, ni permite actualmente, su sobrevivencia. El presente constituye el segundo registro confirmado de la presencia de esta especie en Sudamérica.
Agradecimientos
Agradecemos la colaboración de Luc Ector (Public Research Centre – Gabriel Lippmann, Luxemburgo) y Vaclav Houk (Academy of Sciences of the Czech Republic) quienes nos ayudaron en la identificación inicial de C. meduanae y proveyeron parte de la literatura utilizada en este trabajo. Agradecemos también la ayuda brindada durante el análisis de microscopía electrónica por Marie Cantino y Jim Romanow (Q.E.P.D.) (University of Connecticut Electron Microscopy Laboratory, EUA). El análisis de MEB fue realizado con el patrocinio de Donald Charles (The Academy of Natural Sciences of Drexel University, Filadelfia, EUA). Agradecemos de manera especial a Patricio Rivera (Universidad de Concepción, Chile), Nora Maidana (Universidad de Buenos Aires) y Silvia Sala (Universidad Nacional de La Plata) por datos de la presencia de C. meduanae en Chile y Argentina. El presente trabajo conmemora los 5 años de actividades del Herbario Criptogámico de la Universidad Católica Boliviana en Cochabamba, Bolivia.
Referencias
[1] Acosta, F. & Ayala, R. 2007. Hacia la gestión de lagunas urbanas: La experiencia en la Laguna Alalay (Cochabamba – Bolivia). En: Castro, M. & Fernández, L. (Eds). Gestión Sostenible de Humedales. CYTED-Programa Internacional de Interculturalidad. Chile. 409 pp. [ Links ]
[2] ANSP-TaxaService. 2013. Página: Cyclotella meduanae (NADED 20060) (http://diatom.ansp.org/taxaservice/ShowTaxon1.ashx?naded_id=20060). The Academy of Natural Sciences of Drexel University, Phycology Section: ecology and taxonomy of freshwater algae, particularly diatoms. Consultada Agosto 10, 2013. [ Links ]
[4] Arias, S. 1998. Proyecto: Conservación de la ornitofauna de la laguna Alalay. Centro Universitario de Ecología, Medio Ambiente y Desarrollo (CUEMAD), Universidad Mayor de San Simón (UMSS). 57 pp. [ Links ]
[5] Ayala, R.; Castro, M.; Bayro, V.; Acosta, F. & Rejas, D. 2006. Interacciones fitoplancton – zooplankton en una laguna eutrofizada del valle de Cochabamba (Bolivia). Revista de Ciencia y Tecnología 5: 43-50 [ Links ]
[6] Ayala, R.; Acosta, F.; Mooij, W.M.; Rejas, D. & Van Damme, P.A. 2007. Management of Laguna Alalay: a case study of lake restoration in Andean valleys in Bolivia. Aquatic Ecology 41: 621-630. [ Links ]
[8] Cadima, F.M. 1998. Algas y macrófitas de la laguna Alalay (Cochabamba-Bolivia). Revista Boliviana de Ecología y Conservación Ambiental 3: 35-46. [ Links ]
[9] Cavalcante, K.P.; Tremarin, P.I. & Ludwig, T.A.V. 2013. Taxonomic studies of centric diatoms (Diatomae): unusual nanoplanktonic forms and new records for Brazil. Acta Botánica Brasilica 27(2): 237-251. [ Links ]
[10] de la Barra, N. 2003. Clasificación ecológica de la vegetación acuática en ambientes lacustres de Bolivia. Revista Boliviana de Ecología y Conservación Ambiental 13: 65-93. [ Links ]
[12] Gómez, N. & Bauer, D.E. 2000. Diversidad fitoplanctónica en la franja costera sur del Río de la Plata. Biología Acuática 19: 7-26. [ Links ]
[13] Håkansson, H. 2002. A compilation and evaluation of species in the general Stephanodiscus, Cyclostephanos and Cyclotella with a new genus in the family Stephanodiscaceae. Diatom Research 17: 1-139. [ Links ]
[14] Håkansson, H. & Kling, H. 1994. Cyclotella agassizensis nov. sp. and its relationship to C. quillensis Bailey and other prairie Cyclotella species. Diatom Research 9: 289-301. [ Links ]
[15] Houk, V. 2003. Atlas of freshwater centric diatoms with a brief key and descriptions. Part I. Melosiraceae, Orthosiraceae, Paraliaceae and Aulacosiraceae. Czech Phycology 1(suppl.): 1-109. [ Links ]
[16] Houk, V. & Klee, R. 2007. Atlas of freshwater centric diatoms with a brief key and descriptions. Part II. Melosiraceae and Aulacosiraceae (suppl. to Part I). Fottea 7(2): 85-255. [ Links ]
[17] Houk, V.; Klee, R. & Tanaka, H. 2010. Atlas of freshwater centric diatoms with a brief key and descriptions, Part III: Stephanodiscaceae A, Cyclotella, Tertiarius, Discostella. Fottea 10 (suppl.): 1–498. [ Links ]
[18] Kiss, K.T.; Klee, R.; Ector, L. & Ács, É. 2012. Centric diatoms of large rivers and tributaries in Hungary: morphology and biogeographic distribution. Acta Botanica Croatica 71(2) 1-53. [ Links ]
[19] Maldonado, M.; Van Damme, P. & Rojas, J. 1998. Contaminación y eutrofización en la Cuenca del Río Rocha (Cochabamba). Revista Boliviana de Ecología y Conservación Ambiental 3: 3–9. [ Links ]
[20] Morales, E.A. & Rivera, S.F. 2012. Choice of macrophyte substrate in the use of diatoms as indicators of pond water quality assessment: preliminary data on the case of Alalay Pond (Cochabamba, Bolivia). Lakes, Reservoirs and Ponds 6: 20–42. [ Links ]
[21] Muñoz, H.; Vallejos, A.; Ayala, R. & Acosta, F. 2007. Estructura y dinámica de la comunidad de peces de la laguna Alalay, Cochabamba-Bolivia. En: Meyen, J., Aguirre, L.F. & Moraes R.M. (Eds). Congreso Internacional sobre Desarrollo, Medio Ambiente y Recursos Naturales: Sostenibilidad a Múltiples-Niveles y Escalas Cochabamba - Bolivia. Vol. II: 1166-1171. [ Links ]
[22] Nagumo, T. & Kobayasi, H. 1985. Fine structure of three freshwater and brackish water species of the genus Cyclotella (Bacillariophyceae), C. atomus, C. caspia and C. meduanae. Bulletin of the Plankton Society of Japan 32: 101-109. [ Links ]
[24] Ogawa, K. 1990. Thalassiosiraceae collected from Lake Teganuma, the most hypertrophic lake in Japan. Diatom 5: 59-68. [ Links ]
[25] Pérez, M. 2005. Especiación de metales pesados en sedimentos de la laguna Alalay. Tesis de Maestría en Ingeniería Ambiental. Universidad Mayor de San Simón, Cochabamba, 90 pp. [ Links ]
[26] Pérez, M.C.; Maidana, N.I. & Comas, A. 2009. Phytoplankton composition of the Ebro River estuary, Spain. Acta Botanica Croatica 68: 11-27. [ Links ]
[28] Sabater, S. & Klee, R. 1990. Observations on centric diatoms of the River Ebro Phytoplankton, with special interest on some small Cyclotella. Diatom Research 5: 141-154. [ Links ]
[29] Sala, S. & Ramírez-R, J. 2008. Cyclotella katiana sp. nov. from La Reina swamp, Parque Nacional Los Ratíos, Colombia. Diatom Research 23: 147-157. [ Links ]
[30] Tanaka, H. 2007. Taxonomic studies of the genera Cyclotella (Kützing) Brébisson, Discostella Houk et Klee and Puncticulata Håkansson in the Family Stephanodiscaceae Glezer et Makarova (Bacillariophyta) in Japan. Bibliotheca Diatomologica 53: 1-205. [ Links ]
[31] Van Damme, P.A.; Romero, A.M.; Goitia, E.; Rojas, J.; Cadima, M.; Arce, O. & Romero, M. 1997. Evaluación de estrategias actuales y alternativas para la recuperación de la laguna Alalay (Cochabamba). Centro de Medio Ambiente y Recursos Renovables CEMAR, Universidad Mayor de San Simón, Cochabamba. 91 pp. [ Links ]
[32] Whitton, B.A.; John, D.M.; Kelly, M.G. & Haworth, E.Y. 2003. A coded list of freshwater algae of the British Isles. Second Edition. Centre for Ecology and Hydrology, Natural Environment Research Council. http://www.ceh.ac.uk/data/algae/algae_index.html. Consultada Agosto 9, 2013.

