










Servicios Personalizados
Revista
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkActa Nova
versión On-line ISSN 1683-0789
RevActaNova. v.6 n.1-2 Cochabamba mar. 2013
ARTÍCULO CIENTÍFICO
Surirella antioquiensis sp. nov. and S. rafaelii sp. nov. (Bacillariophyta) from Colombia
Surirella antioquiensis sp. nov. y S. rafaelii sp. nov. (Bacillariophyta) de Colombia
Silvia E. Sala1, John J. Ramírez2, Amelia A. Vouilloud1 & Yasmín Plata-Díaz C.3
1División Científica Ficología Dr. Sebastian Guarrera, Facultad de Ciencias Naturales y Museo. Paseo del Bosque s/n. 1900 La Plata, Argentina.
2Instituto de Biología, Universidad de Antioquia. Apartado 1226 Medellín, Colombia.
3Grupo de Estudios en Biodiversidad, Departamento de Biología, Universidad Industrial de Santander. Apartado 678 Bucaramanga, Colombia.
e-mail: sesala@museo.fcnym.unlp.edu.ar
Recibido: 20 de octubre 2013; Aceptado: 7 de diciembre 2013
Summary: This article is part of a series of studies to produce and inventory of the diatom flora of Colombia. Two Surirella taxa -S. antioquiensis S.E. Sala, J.J. Ramírez, Plata-Díaz & Vouilloud and S. rafaelii J.J. Ramírez & S.E. Sala- from lotic and lentic aquatic systems in the Department of Antioquia (Colombia) are proposed as new species. Both new species were analyzed with light and scanning electron microscopy. Each taxon is described and illustrated and information about environmental conditions of sampling sites is given.
Key Words: Andean region, Colombia, diatoms, lentic systems, lotic systems, Surirella, Tropics
Resumen: Este artículo forma parte de una serie de estudios que se llevan a cabo para realizar un inventario de la flora diatomológica de Colombia. En él se describen dos especies del género Surirella -S. antioquiensis S.E. Sala, J.J. Ramírez, Plata-Díaz & Vouilloud y S. rafaelii J.J. Ramírez & S.E. Sala- provenientes de sistemas lénticos y lóticos localizados en el Departamento de Antioquia, Colombia. Ambas especies fueron analizadas con microscopio de luz con contraste de fases y microscopio electrónico de barrido. Cada taxón es descrito e ilustrado y se brinda información acerca de las condiciones ambientales de los sitios en que fueron colectados.
Palabras clave: Colombia, diatomeas, región Andina, sistemas lénticos, sistemas lóticos, Surirella, trópicos
1. Introduction
Although it is widely assumed that microalgae are ubiquous and cosmopolitan, there are evidences that this is an erroneous assumption especially in tropical and subtropical areas. One of the most consulted diatom references around the world is the Süβwasserflora von Mitteleuropa [25][27][28][29], but as Metzeltin & García-Rodríguez [36] pointed out, it should be used only in Central Europe and with caution in other zones. This limitation owes to the fact that the species of Central Europe could be quite similar to those of other regions, but they are not always the same from other latitudes [1][12][13][14][34][35][39], especially those from the circumtropical areas. We are convinced that researchers of these latitudes must invest a great effort to generate their own information. The present study is part of a series of investigations carried out in order to inventory the diatom flora of Colombia.
The genus Surirella Turpin is the founding member of the family Surirellaceae, established by Kützing [30]. According to Fourtanier & Kociolek [20], Surirella comprises around 1,700 species. Peragallo & Peragallo [41], divided the genus in three subgenera groupings -Fastuosae, Robustae and Pinnatae- which differ in the morphology of the keel and associated supporting structures [6]. Ruck & Kociolek [47] held a preliminary analysis of the phylogeny of the Family Surirellaceae and found that Surirella is not a monophyletic group, but until more studies are held it is useful to follow the classic system of clustering.
While most representatives of the Fastuosae group live in marine and brackish habitats, have keels directly attached to the valve, opening internally through portulae, the Robustae group lives predominantly in freshwaters and the major part of the species have keels and wings connected to the valve by alar canals separated by fenestrae. The Pinnatae group can be separated into two subgroups, one comprising the big forms with morphology similar to S. ovalis Bréb.; and the second group -conformed around S. minuta Bréb. ex Kütz.- that comprises more delicate forms [26]. The species of the Pinnatae group do not have wings or alar canals, but have costate fibulae and portulae that connect the raphe canal with the inner part of the valve. The costae are transverse and the striae are generally multiseriate; both interrupted at valve centre by a hyaline area. The frustule is heavily silicified, isopolar or heteropolar, sometimes twisted. The valves are linear, elliptic or ovate, plane or concave.
Most Surirella species have a subcosmopolitan distribution -defined by O´Sullivan & Reynolds [40], as species that occur throughout the world, but always in specialized environments- and a highest species richness in circumtropical environments. According to Karthick et al. [24], Surirella species from tropical freshwaters have received much attention due to the high percentage of endemisms reported across different continents. Many Surirella species from tropical regions were described during the 19th and early 20th centuries by Ehrenberg, Müller and West [12][14]. Karthick et al. [24] pointed out that Ehrenberg [19], reported the first Surirella species from the Indian subcontinent and was followed by Dickie [16], and West & West [59], among others. Cocquyt & Jahn [9][11][12][13] considered that Surirella species are typical components of the diatom flora of the African Great Lakes (Malawi, Nyassa, Tanganika and Victoria), with nearly 50 taxa, several of them endemic from Eastern and Central Africa. In the Malili Lakes located in the Sulawesi Island in Indonesia, Hustedt (in Huber-Pestalozzi [21]) identified 46 taxa of Surirella, many of them endemic, and later Bramburger et al. [6] studied 35 species from the same lakes, eleven of which were new to science. According to these latter authors, the high number of Surirella species found in the lakes of Malili put these sites at the top as the world hotspot of diversity for this genus. Cocquyt [8] found 36 Surirella taxa in the northern basin of Lake Tanganyika. In Papua New Guinea, Vyverman [56] registered 17 taxa and many unknown taxa. These findings place these zones as areas with a diversity comparable to that of the Malili Lakes. In the tropics of America, Krasske described S. arctissima var. muscicola Krasske from Brazil and S. columbiensis Krasske from Colombia and Brazil [31]. More recently, Metzeltin & Lange-Bertalot [34] reported S. citrus Metzeltin & Lange-Bert. from Kaiereteur Falls, Potaro River (Guyana), S. rotti Metzeltin & Lange-Bert. from Iguassu River (Brazil), S. rumrichorum Metzeltin & Lange-Bert. from Laguna Negra (Venezuela), S. tenerisilex Metzeltin & Lange-Bert. from Essequibo River (Guyana) and S. susanae Metzeltin & Lange-Bert. from Laguna Agua Azul (México). Wydrzycka & Lange-Bertalot [58] registered S. bryophila Lange-Bert., Wydrzycka & Metzeltin from the Agrio River in Costa Rica, while Blanco et al. [5] described S. moralesii S. Blanco, Álvarez-Blanco & Cejudo-Figueiras from Lake Honda in Bolivia.
Due to their large and heavy frustules, Surirella species are rarely euplanktonic and live in littoral habitats, being part of the phycoperiphyton, the metaphyton, the episammon or the epilithon [6]. Besides, Surirella species are sometimes common in acid waters in high-altitude lakes and peats and they develop in waters moderately rich in organic matter, but they are more common in waters with high temperatures, alkaline pH values, and relatively high conductivity [6].
In Colombia, although there are numerous records of diatoms in studies on periphyton ecology [15][18][33][38][45][60], paleoecological investigations [32], and studies on phytoplankton ecology [3][42][43][44], among other fields, the knowledge on taxonomy of freshwater algae in general -and particularly for the diatoms- is scarce. Identifications are mostly made by ecologists, with the consequent burden of mistakes that this practice generates. In the majority of the mentioned papers, Surirella taxa were identified up to the genus level and the only reports at the species level are S. columbiensis Krasske [17] and S. guatimalensis Ehrenb. [37].
In a preliminary study held in lentic and lotic systems from the Antioquia Department, almost 90 diatom taxa were identified (unpublished data). Approximately 30% of them could not be assigned to recognized taxonomic entities, so they are presumably new for science. In this article, we describe two Surirella taxa found in this Colombian region.
2. Methodology
Samples containing the two new species described here were collected from lotic and lentic environments in the Department of Antioquia (05º 26' 20'' N, 08º 52' 23'' N and 73º 53' 11'' W, 77º 07' 16'' W), which is located in the Andean mountainous system in Colombia.
Epilithon samples from La Vega Stream (San José del Nus Municipality) were collected between August, 2001 and April, 2002 at three sampling places. The sample from El Carmen de Viboral was collected squeezing plants of Nymphoides sp., the dominant macrophyte of a small pond in November, 2003.
Samples were fixed with 10% Lugol or 6-8% formaline and were treated to eliminate organic matter following the method described in CENT 230 [7]. Samples for light microscopy (LM) were mounted in Naphrax ®; and for scanning electron microscopy (SEM) they were mounted on glass stubs and coated with gold-palladium in a Jeol FINE COAT ION SPUTTER JFC-1100. Observations were conducted with a Wild M20 LM and a Leica equipped with DM 2500 phase contrast for LM and a Jeol JSM-T100 for SEM at the Servicio de Microscopía Electrónica del Museo de La Plata.
The terminology used is that suggested in Anonymous [2], Barber & Harworth [4], Ross et al. [46] and Ruck & Kociolek [47].
Type material, holotypes and isotypes are deposited in the Herbario, Departamento Científico, Ficología, Museo de Ciencias Naturales de La Plata, Argentina (LPC). Although we analyzed several samples from Antioquia, we mention only those containing the new species:
- LPC10003: La Vega Stream, San José del Nus Municipality, Department of Antioquia, Colombia. April, 2002. Collector: Esnedy Hernández.
- LPC10011: El Carmen de Viboral Municipality, Department of Antioquia, Colombia. November, 2003. Collector: Yimmy Montoya.
3. Results and Discussion
3.1. Surirella antioquiensis S.E. Sala, J.J. Ramírez, Plata-Díaz & Vouilloud sp. nov. (Figs 1-2)
Holotype: Designated here: LPC 10011 (Slide1, Finder: P 34-2). El Carmen de Viboral Municipality, November, 2003. Collector: Yimmy Montoya. The valve representing the holotype is illustrated in Figs 1B-C.
Type locality: El Carmen de Viboral Municipality, Department of Antioquia, Colombia (6° 05' 09'' N, 75° 20' 19'' W).
Etymology: the specific epithet antioquiensis refers to the Department of Antioquia, the geographic region where the type locality is located.
Description. Light Microscope. Cells isopolar, lanceolate with cuneate poles; the longer specimens with a more or less linear outline. Valve face without concentric undulations, transapical costae strongly visible reaching the conspicuous axial area, parallel at valve center radial to the poles (Figs 1A-G).
Scanning Electron Microscope. Valves slightly curved towards the girdle at the poles. Valve face in external view slightly transapically undulated due to the distinct and irregular groupings of striae and Type II costae separated by 1-2 small sunken costae that extend out to the axial area, a straight hyaline line (Figs 1H-L). Each group has 3-4 uniseriate striae composed of circular areolae exceptionally transapically elongated (Fig. 1K). Striae and costae are radial towards the poles and parallel at the valve center (Figs 1H-J). The mantle is shallow with the same height alongside and the same structure of the valve face (Figs 1J, L).

In internal valve view, fibulae and portulae are evenly spaced (Figs 2A-D). Fibulae type III, reaching the sternum, are robust and broaden at the basis. The interfibular spaces present one elliptical portula. The fibulae correspond to the externally sunken costae (Figs 2C-D). Raphe canal smooth only slightly elevated above the valve surface. Raphe interrupted at both poles externally, with straight and distant ends without terminal pores (Fig. 1L).

Morphometric data (n=30): length: 14-32 µm, width: 6.4-7.6 μm, L/W: 2.7-4.5, striae: 26-27 in 10 μm: fibulae: 60-82 in 100 μm, areolae: 53-65 in 10 μm.
Ecological data: epiphythic taxon on Nymphoides sp. plants in a small pond located at 2,200 m a.s.l. This species is abundant and coexists with small diatoms of the genera Gomphonema Ehrenb., Achnanthidium Kütz., Planothidium Round & Bukhtiyarova and Nitzschia Hassall.
Distribution: This species is only known from a small pond at El Carmen de Viboral Municipality.
Remarks: the studied material belongs to the Pinnatae group; it is similar in size and structure to S. minuta, but clearly differs in the isopolar valve outline. Surirella angusta Kütz., a small isopolar species, is the closest described taxon. Information about the valve morphology of this widely distributed species is scarce, so comparisons were done against the type material photographed with LM in Krammer & Lange-Bertalot [29]. Our material differs from S. angusta in its smaller size and also in valve outline, lanceolate in S. antioquiensis and with parallel margins in S. angusta (Table 1).
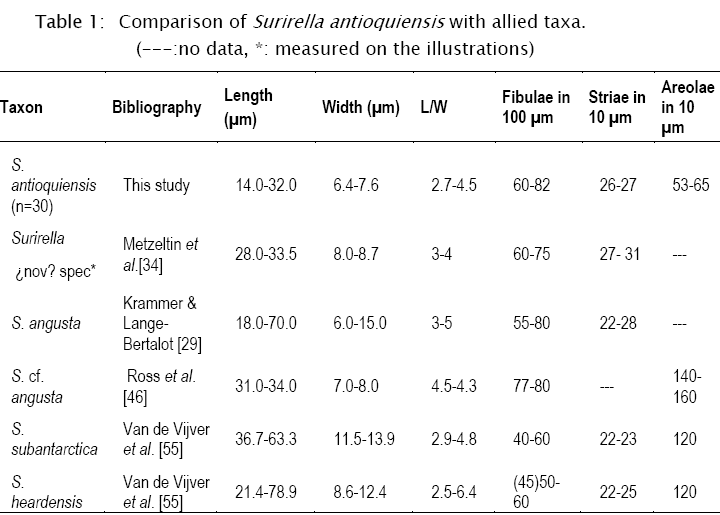
The studied material is also similar to materials from Crater Lake, Oregon (CAS 622015) designated as S. cf. angusta in [47]. LM photographs [47] show that this taxon has straight margins in valve view, slightly constricted at midvalve, and parallel costa alongside. Besides, under SEM, they differ in striae structure, multiseriate in S. cf. angusta and uniseriate in S. antioquiensis. The lower costae are connected to the grouped costa, by few and fine siliceous elements in S. antioquiensis (Fig. 1K), while in S. cf. angusta these elements are numerous and conspicuous [47]. The studied material differs also in valve structure with the North American valves in their wider axial area and the parallel and short costae all alongside [47]. These two taxa also differ in fibulae and areolae density (Table 1). Although, in external view these taxa can be confused, differences in internal view are conspicuous as the valves in S. cf. angusta are more robust, fibulae are short and thick and interfibular spaces are triangular.
The specimens described in [34] from Brazil, Iguazú Falls and Río Tec (Venezuela) as Surirella (nov?) spec. coincide with S. antioquiensis in dimensions, general LM and SEM appearance, and South American tropical distribution, but other details of this taxon are insufficient to be sure if they are conspecific (Table 1).
Other taxa with similar valve morphology are S. subantarctica Van de Vijver & Cocquyt and S. heardensis Van de Vijver & Cocquyt [55]. These taxa differ from the studied material in dimensions (Table 1), but also in the multiseriate structure of the striae, the number of portulae: a large one with two small ones on each side, among other morphological details.
Surirella rafaelii J.J. Ramírez & S.E. Sala sp. nov. (Figs 3-4)
Holotype: Designated here: LPC 10003 (slide 1, Finder: O28-4). La Vega Stream, Municipality of San José del Nus, April, 2002. Collector: Esnedy Hernández. The valve representing the holotype is illustrated in Figs 3B-C.
Type locality: Municipality of San José del Nus, Department of Antioquia, Colombia. (6° 29' 30'' N, 74° 49' 30'' W).
Etymology: the specific epithet rafaelii refers to the Zoologist Rafael Urrejola, Universidad Nacional de La Plata, Buenos Aires, Argentina.
Description. Light Microscope. Frustules isopolar in valve and girdle view (Figs 3A-F, 4A). Valves parallel with a short pervalvar axis (Fig. 4A). Valves linear-lanceolate with parallel margins in the smaller specimens (Figs 3A-B) and slightly constricted at midvalve in larger specimens (Figs 3 C-F), apices bluntly cuneate. Valve face strongly transpically undulated, axial area straight slightly elevated with a conspicuous median ridge not reaching the apices (Figs 3 A-F).
Scanning Electron Microscope. Valve face regularly corrugated; axial area sligthly elevated in external view with a thin irregular ridge not reaching the valve apices (Figs 3G-H). The entire valve face is transapically striated. Striae are uniseriate with circular areolae difficult to see even with SEM (Figs 3H-J). In external view, the interstriae are covered with small granules arranged in transapical rows, more densely disposed near the central keel (Fig. 3H). Alar canals slightly wider than the fenestrae (Fig. 3G). Wings well developed (Fig. 4B), fenestrae short with 3-6 fenestral bars (Figs 3H, J, 4 B-D). Tendrils solitary or eventually grouped (2-3), ribbon-like with sharp ends placed at the base of the fenestrae (Figs 3J, 4B-D, F). Raphe canal with transversal ribs (Fig. 3H-J), raphe externally interrupted at both ends, curved towards the mantle and with terminal small pores (Figs 3 I-J, 4F). Mantle completely perforated with areolae irregularly arranged (Fig. 4E). Girdle with few broad bands, valvocopula interrupted near midvalve (Figs 4B, G). Internally, the alar canals open in simple portulae (Figs 4 E, H-I) and the mantle and the valve face have the same structure (Figs 4I).


Morphometric data (n=40): length: 56-112 µm, width: 14-19 µm, L/W: 3-7, pervalvar axis: 15.7-17.8 µm, alar canals: 21-30/100 µm, striae in 10 µm: 30-48, areolae in 10 µm: approximately 60.
Ecological data: elevation: 651 m a.s.l., water temperature: 21.4-26.3º C, pH: 6.64-7.90, dissolved oxygen: 6.9-8.0 mg L-1. In the study area this species is rare and coexists with diatoms of the genera Cocconeis Ehrenb., Navicula Bory and other small naviculoids.
Distribution: this species is only known from a stream at San Jose del Nus Municipality.
Remarks: this taxon belongs to the Robustae group and was compared with isopolar taxa with median ridge as Surirella engleri O. Müll., S. sublinearis Hust., S. sublinearis var. suggesta Bramburger & P.B. Hamilton, S. angusticostata Hust., S. susanae Metzeltin & Lange-Bert., S. feuerborni Hust., S. biseriata var. splendida f. constricta Hust., S. gradifera Hust. and S. acuminata Hust. (Table 2). Surirella engleri, S. susanae, S. feuerbornii, S. biseriata var. splendida f. constricta and S. gradifera differ from the studied material in dimensions and/or alar canal densities.
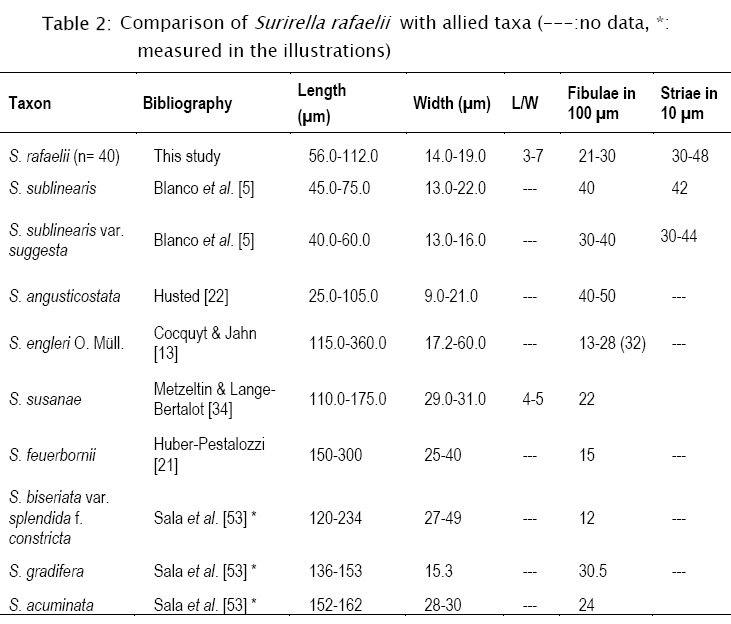
Besides, S. rafaelii resembles S. engleri in the valve outline and the axial area, but the type material studied by Cocquyt & Jahn [13] differs from our material in the siliceous granules only distributed on the depressions of the valve face, the elongated fenestrae, the absence of tendrils and the raphe channel with longitudinal costae. The studied material differs from S. sublinearis and S. sublinearis var. suggesta [6] in the numerous alar canals (see Table 2), in the absence of a silica ridge or keel along the axial area, and the long and grouped filiform tendrils. Besides, it differs from var. suggesta in the spinules arranged in parallel rows across the valve face becoming irregular dispersed in the central area and also in the raphe endings. All of these species were described from tropical areas, but only S. susanae from South America.
4. Conclusions
This study was held within the framework of a research project to produce an inventory of the Colombian diatom flora. The research is carried out in different regions of the country such as Antioquia, Santander, Chocó, Córdoba and Amazonia, as well as low and high-altitude waterbodies. While the genus Surirella is highly diversified in tropical zones of Africa and Asia, few species have been described from the neotropics until now. In the Colombian regions studied thus far, the genus was poorly represented both from qualitative and quantitative points of view. The species analyzed here were only found in Antioquia, a Department that belongs to the Antioquian Mountain subregion located in the Andean region. These aquatic systems are located in sites with different elevation. While La Vega stream is placed between 1,250 and 790 m a.s.l. in the subtropical level, El Carmen de Viboral is located between 3,340 and 800 m a.s.l. in the mesothermic or temperate level.
Surirella antioquiensis is close to S. angusta and S. cf. angusta in Ruck & Kociolek [47] and Surirella (¿nov?) spec. in Meltzeltin & Lange-Bertalot [34]. The latter species, collected in Brazil and Venezuela, is similar in external valve morphology to the studied material, but data given by the authors are insufficient to establish if they are conspecific.
Surirella rafaelii, an isopolar species, was compared with related taxa with prominent median ridge that belongs to the Robustae group. Although there are several species that can be confused under LM, details of the valve morphology allow its distinction from other species. Filiform tendrils are present in other Surirella species [6], but ribbon like tendrils have not been described for any species in the genus up to this date.
The results of this study are consistent with those obtained in previous studies about the genera Brachysira Kütz., Cyclotella Kütz. ex Bréb., Encyonema Kütz., Urosolenia Round & R.M. Crawford, from continental water bodies of Colombia, where near one third of the taxa could not be assigned to known entities [48][49][50][51][52][53][57]. This situation allows us to conclude that Colombia has a richer diatom flora than presumed to date, and that it is necessary to generate literature for identification of local taxa, especially if the intention is to apply this information to bioindication, paleoecology, and/or limnology.
Acknowledgments
We thank the Institute of Biología of Universidad de Antioquia, Colciencias, and the Red Latinomericana de Botánica (RLB) for the fellowships given to Yasmin Plata Díaz.
References
[1] Álvarez-Blanco, I.; Cejudo-Figueiras, C.; de Godos I.; Muñoz, R. & Blanco, S. 2011. Las diatomeas de los salares del Altiplano boliviano: singularidades florísticas. Boletín De la Real Sociedad Española de Historia Natural Sección Biología 105: 67-82. [ Links ]
[2] Anonymous. 1975. Proposals for standardization of diatom terminology and diagnoses. Nova Hedwigia 53: 232-354. [ Links ]
[3] Asprilla, S.; Ramírez, J.J. & Roldán, G. 1998. Caracterización limnológica de la ciénaga de Joutaudó (Chocó, Colombia). Actualidades en Biología 20: 87-107. [ Links ]
[4] Barber, H.G. & Haworth, E.Y. 1981. A guide to the morphology of the diatom frustule. Freshwater Biological Association 44, Ambleside, 112 pp. [ Links ]
[5] Blanco, S.; Álvarez-Blanco, I.; Cejudo-Figueiras, C.; de Godos, I.; Bécares, E.; Muñoz, R.; Guzmán, H.O.; Vargas, V.A. & Soto, R. 2013. New diatom taxa from high-altitude Andean saline lakes. Diatom Research 28: 13-27. [ Links ]
[6] Bramburger, J.; Haffner, G.D.; Hamilton, P.B.; Finz, H. & Hehanussa, P.E. 2006. An examination of species within the genus Surirella from the Malili lakes, Sulawesi Island, Indonesia, with descriptions of 11 new taxa. Diatom Research 21: 1-56. [ Links ]
[8] Cocquyt, C. 1998. Diatoms from the Northern Basin of Lake Tanganyika. Bibliotheca Diatomologica 39: 1-275 [ Links ]
[9] Cocquyt, C. & Jahn, R. 2005a. Rare Surirella taxa (Bacillariophyta) from East Africa described by Otto Müller: typifications, recombinations, new names, annotations and distributions. Willdenovia 35: 359-371. [ Links ]
[10] Cocquyt, C. & Jahn,R. 2005b. Surirella capensis Ehrenberg ex Cocquyt et R. Jhan sp. nov., a rare and historical diatom species from South Africa. Crytogamie Algologie 26: 149-153. [ Links ]
[11] Cocquyt, C. & Jahn,R. 2007a. Surirella nyassae O. Müller, S. malombae O. Müller and S. chepurnovii Cocquyt & R. Jahn sp. nov. (Bacillariophyta) – typification and variability of three closely related East African diatoms. Nova Hedwigia 84: 529-548. [ Links ]
[12] Cocquyt, C. & Jahn,R. 2007b. Taxa related to Surirella ovalis from Tanzania, East Africa: typification of O. Müllers taxa and description of a new species. Crytogamie Algologie 28: 107-116. [ Links ]
[13] Cocquyt, C. & Jahn,R. 2007c. Surirella engleri O. Muller – a study of its original infraspecific types, variability and distribution. Diatom Research 22: 1-16. [ Links ]
[14] Cocquyt, C. & Kusber, W.H. 2010. Reinvestigation of West African Surirellaceae (Bacillariophyta) described by Foged from Ghana. Nova Hedwigia 91: 111–136. [ Links ]
[15] Díaz, C.A. & Rivera, C. 2004. Diatomeas de pequeños ríos andinos y su utilización como indicadoras de condiciones ambientales. Caldasia 26: 381-394. [ Links ]
[16] Dickie, G. 1882. Notes on algae from the Himalayas. Journal of Linnean Society (Botany) 19: 230. [ Links ]
[17] Donato, J.C. 2010. Phytoplankton of Andean Lakes in northern South America (Colombia). Composition and environment distribution factors. In: Witkowski A. (Ed.). Diatom Monographs. Gantner Verlag, Germany. 188 pp. [ Links ]
[19] Ehrenberg, C.G. 1845. Novorum Generum et Specierum brevis definitio. Zusätze zu seinen letzten Mit-theilung über die mikroskopischen Lebensformen von Portugall und Spanien, Süd-Afrika, Hinter-Indien,Japan und Kurdistan, und legte die folgenden Diagnosen u.s.w. Bericht über die zur Bekanntmachung geeigneten Verhandlungen der Königlich-Preussischen Akademie der Wissenschaften zu Berlin 1845: 357– 377.
[20] Fourtanier, E. & Kociolek, J.P. 2011. Catalogue of Diatom Names. Available online at http://research.calacademy.org/research/diatoms/names/index.asp
[21] Huber- Pestalozzi, G. 1942. Das Phytoplankton des Susswassers; Systematik und Biologie. In: Thienemann, A. (Ed.). Die Binnengewasser. 2 Teil. 2 Halfte. Diatomeen. Stuttgart, E. Schweizerbartsche Verlagsbuchhandlung. pp 367-549. [ Links ]
[22] Hustedt, F. 1935. Die fossile diatomeenflora in den Ablagerungen des Tobasees auf Sumatra. Archiv für Hydrobiologie, Supplement 14: 143-192.
[23] Hustedt, F. 1938. Systematische und ökologische Untersuchungen über die Diatomeen flora von Java, Bali und Sumatra nach dem Material der Deutschen Limnologischen Sunda_Expedition. Teil I. Archiv für Hydrobiologie, Supplement 15, 393-506.
[24] Karthick, B.; Hamilton, P.B. & Kociolek, J.P. 2012. Taxonomy and biogeography of some Surirella Turpin (Bacillariophyceae) taxa from Peninsular India. Nova Hedwigia, Beiheft 141: 81–116. [ Links ]
[25] Krammer, K. & Lange-Bertalot, H. 1986. Bacillariophyceae. In: Süsswasserflora von Mittleuropa. Band 2, Teil 1. Naviculaceae. Gustav Fisher, Jena. 876 pp.
[26] Krammer, K. & Lange-Bertalot, H. 1987. Morphology and taxonomy of Surirella ovalis and related taxa. Diatom Research 2: 77-95. [ Links ]
[27] Krammer, K. & Lange-Bertalot, H. 1991a. Bacillariophyceae. 3. Teil: Centrales, Fragilariaceae, Eunotiaceae. In: Ettl, H.; Gerloff, J.; Heynig, H. & D. Mollenhauer (Eds). Süsswasserflora von Mitteleuropa, Band 2/3. Gustav Fischer Verlag, Jena. 576 pp. [ Links ]
[28] Krammer, K. & Lange-Bertalot, H. 1991b. Bacillariophyceae. 4. Teil: Achnanthaceae, Kritische Ergänzungen zu Navicula (Lineolatae) und Gomphonema. In: Ettl, H.; Gerloff, J.; Heynig, H. & D. Mollenhauer (Eds). Süsswasserflora von Mitteleuropa, Band 2/4. Gustav Fischer Verlag, Stuttgart, Jena. 437 pp.
[29] Krammer, K. & Lange-Bertalot, H. 1997. Bacillariophyceae. 2. Teil: Bacillariaceae, Epithemiaceae, Surirellaceae. In: Ettl, H.; Gerloff, J.; Heynig, H. & D. Mollenhauer (Eds). Süsswasserflora von Mitteleuropa, Band 2/2. Gustav Fischer, Jena. 611 pp.
[30] Kutzing, F.T. 1844. Die Kieselschaligen bacillarien oder Diatomeen. Nordhausen, 152 pp. [ Links ]
[31] Lange-Bertalot, H.; Külbs, K.; Lauser, T.; Nörpel-Schempp, M. & Willman, M. 1996. Diatom taxa introduced by Georg Krasske Documentation and Revision. Iconographia Diatomologica. Annotated Diatom Micrographs. Volume 3. Koeltz Scientific Books, Königstein. 358 pp. [ Links ]
[32] Lozano, C.G.; Parra, L.N.; Moro, R.S. & Ramírez, J.J. 1999. Inferencias paleolimnológicas del Holoceno con base en diatomeas en la laguna Puente Largo, Páramo de Frontino, Antioquia, p.57-116. En: M.T. Flórez & C.G. Lozano (Eds) Silicofósiles altoandinos. Colciencias, Universidad Nacional de Medellín, Comité de Investigaciones Universidad de Antioquia, Medellín. 116 pp. [ Links ]
[33] Martínez, E.L. & Donato, J.C. 2003. Efectos del caudal sobre la colonización de algas en un río de alta montaña tropical (Boyacá, Colombia). Caldasia 25: 337-354. [ Links ]
[34] Metzeltin, D. & Lange-Bertalot, H. 1998. Tropical Diatoms of South America I. Koeltz Scientific Books, Germany. 695 pp. [ Links ]
[35] Metzeltin, D. & Lange-Bertalot, H. 2002. Diatoms of the island continent Madagascar. In: Lange-Bertalot, H. (Ed.). Iconographia Diatomologica. Vol. 11. Koeltz Scientific Books, Koenigstein. 877 pp. [ Links ]
[36] Metzeltin, D. & García-Rodríguez, F. 2003. Las Diatomeas Uruguayas. D.I.R.A.C. Facultad de Ciencias, Montevideo. 208 pp. [ Links ]
[37] Modesto-Iregui, M.; Guillot, G.; Donato, J.C. & Obregón, M.T. 2002. Dimensión fractal y relación área superficial/volumen de algas del fitoplancton de lagos colombianos. Caldasia 24: 121-134. [ Links ]
[38] Montoya-M, y.; Ramírez-Restrepo, J.J. & Segecin-Moro, R. 2008. Diatomeas perifíticas de la zona de ritral del río Medellín (Antioquia) Colombia. Actualidades Biológicas 30: 181-192. [ Links ]
[39] Morales, E.A.; Fernández, E. & Kociolek, P.J. 2009. Epilithic diatoms (Bacillariophyta) from cloud forest and alpine streams in Bolivia, South America 3: diatoms from Sehuencas, Carrasco National Park, Department of Cochabamba. Acta Botanica Croatica 68: 263-283. [ Links ]
[40] OSullivan, P.E. & Reynolds C.S. (Eds). 2004. The Lakes Handbook. Vol.1. Limnology and limnetic ecology. Blackwell Science Ltd., Oxford. 699 pp
[41] Peragallo, H. & Peragallo, M. 1897-1908. Diatomées marines de France et des districts maritime s voisins. Grez-sur-Loing, Paris. 492 pp. [ Links ]
[42] Plata, Y. 2000. Distribución horizontal, biomasa y productividad primaria de la comunidad fitoplantónica de la ciénaga de Chucurí (Santander, Colombia). Trabajo de grado. Universidad Industrial de Santander, Bucaramanga. 125 pp. [ Links ]
[43] Ramírez, J.J. & Díaz, A. 1994. Caracterización limnológica y estructura de la comunidad fitoplanctónica en la laguna del Parque Norte, Medellín, Colombia. Hoehnea 21: 7-28. [ Links ]
[44] Ramírez, J.J.; Bicudo, C.E.; Roldán, G. & García, L.C. 2000. Temporal and vertical variations on the phytoplankton community structure and its relation with some morphometric factors in four Colombian reservoirs. Caldasia 22: 108-126. [ Links ]
[45] Ramírez, A.M. & Plata-Díaz, Y. 2008. Diatomeas perifíticas en diferentes tramos de dos sistemas lóticos de alta montaña (Páramo de Santurbán, Norte de Santander, Colombia) y su relación con las variables ambientales. Acta Biológica Colombiana 13: 199-216. [ Links ]
[46] Ross, R.; Cox, E.J.; Karayeva, D.G.; Mann, D.G.; Padock, T.B.; Simonsen, R. & Sims, P.A. 1979. An emended terminology for the siliceous components of the diatom cell. Nova Hedwigia 64: 513-533. [ Links ]
[47] Ruck, E.C. & Kociolek, J.P. 2004. Preliminary phylogeny of the Family Surirellaceae (Bacillariophyta). J. Cramer, Berlin. 236 pp. [ Links ]
[48] Sala, s.E.; Duque, S.R.; Núñez-Avellaneda, M. & Lamaro, A.A. 1999. Nuevos registros de diatomeas (Bacillariophyceae) de la Amazonía Colombiana. Caldasia 21: 26-37. [ Links ]
[49] Sala, S.E.; Duque, S.R.; Núñez-Avellaneda, M. & Lamaro, A.A. 2002a. Diatoms from the Colombian Amazonia. Cryptogamie Algologie. 23: 75-99. [ Links ]
[50] Sala, S.E.; Duque, S.R.; Núñez-Avellaneda, M. & Lamaro, A.A. 2002b. Diatoms from the Colombian Amazon: some species of the genus Eunotia (Bacillariophyceae). Acta Amazonica 32: 589-603. [ Links ]
[51] Sala, S.E. & Ramírez, J.J. 2008. Cyclotella katiana nov. sp. (Bacillariophyceae) from La Reina Swamp, Parque Nacional Natural Los Katíos, Colombia. Diatom Research 23: 147-157. [ Links ]
[52] Sala, S.E.; Núñez –Avellaneda, M. & Vouilloud, A.A. 2008a. Ultrastructure of the frustule of Urosolenia species from the Colombian and Peruvian Amazon: U. delicatissima spec. nov., U. amazonica spec. nov. and U. braunii (Hustedt) Rott & Kling. Diatom Research 23: 159-169. [ Links ]
[53] Sala, S.E.; Ramírez, J.J. & Plata, Y. 2008b. Diatoms from lentic and lotic systems in Antioquia, Chocó and Santander Departments in Colombia. Revista de Biología Tropical (International Journal of Tropical Biology) 56: 1159-1178. [ Links ]
[54] Simonsen, R. 1987. Atlas and catalogue of the diatoms types of Friedrich Hustedt. Vol. I: Catalogue. 525 pp. Vol 2: Atlas, Taf. 1-395. Vol. 3: Atlas, Taf. 396-772. J. Cramer, Berlin.
[55] Van de Vijver, B.; Cocquyt, C.; De Haan, M.; Kopalová, K. & Zidarova, R. The genus Surirella (Bacillariophyta) in the sub-Antartic and maritime Antartic region. Diatom research 28: 93-108. [ Links ]
[56] Vyverman, W. 1991. Diatoms of Papua Guinea. Bibliotheca Diatomologica 22: 1-224 [ Links ]
[57] Vouilloud, A.A.; Sala, S.E.; Núñez-Avellaneda, M. & Duque, S.R. 2010. Diatoms from the Colombian and Peruvian Amazon: the Genera Encyonema, Encyonopsis and Gomphonema. Revista de Biología Tropical (International Journal of Tropical Biology) 58: 45-62. [ Links ]
[58] Wydrzycka, U. & Lange-Bertalot, H. 2001. Las diatomeas (Bacillariophyceae) acidófilas del río Agrio y sitios vinculados con su cuenca, Volcán Poás, Costa Rica. Brenesia 55-56: 1-68. [ Links ]
[59] West, W. & West, G.S. 1907. Fresh-water algae from Burma including a few from Bengal and Madras. Annals of the Royal Botanic Gardens, Calcutta 6 (2): 175-260. [ Links ]
[60] Zapata, A.M. & Donato, J.C. 2005. Cambios diarios de las algas perifíticas y su relación con la velocidad de corriente en un río tropical de montaña (Río Tota - Colombia). Limnetica 24: 327-338. [ Links ]

