









 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO  uBio
uBio 
 Permalink
PermalinkEl uso de herbicidas en Bolivia se ha incrementado en gran medida en las últimas décadas y forma parte de todas aquellas amenazas a la biodiversidad boliviana producto del proceso de expansión agrícola (Vos et al. 2020). Constituye también una amenaza para la salud de los productores, situación que se agudiza debido a la aplicación inadecuada que realizan los agricultores (Peñafiel & Kammerbauer 2001). Si bien los herbicidas no tienen tanta importancia en el cultivo de cacao (Theobroma cacao, Malvaceae) como los fungicidas e insecticidas (Azembouh et al. 2021), existe un amplio uso de estos en los países que producen cacao en grandes volúmenes (Nkamleu et al. 2007, Fahmid et al. 2018, Aminu & Edun 2019).
En un cultivo, además de las especies cultivadas, residen las plantas espontáneas, que son aquellas que forman parte del cultivo, pero no fueron sembradas. Generalmente se trata de hierbas y/o subarbustos en términos de importancia, pero se puede encontrar también una alta riqueza de especies de plántulas o rebrotes de leñosas en muy bajas densidades (Marconi & Armengot 2020). La aplicación de herbicidas puede eliminar y limitar a algunas especies de plantas espontáneas, así como favorecer a otras. En algunos casos algunas poblaciones se tornan tolerantes o resistentes a estos agroquímicos con el paso de las generaciones (Baek et al. 2021).
En anteriores estudios encontramos en monocultivos convencionales de cacao un grupo específico de hierbas que en sistemas sin aplicación de herbicidas se encontraban en bajas densidades o prácticamente ausentes (Marconi & Armengot 2020, Marconi et al. 2022). Estas hierbas suelen tener una amplia distribución geográfica, a diferencia de aquellas que aparecían en los sistemas agroforestales más complejos y orgánicos, demostrando que el manejo intensivo promueve la homogenización biótica de las hierbas (Marconi & Armengot 2020). La presencia de estas especies es producto de una dinámica más acentuada que en otros sistemas, ya que se encontró mayor cantidad de especies inmigrantes (es decir, que ingresan en la parcela) y de extinciones locales (especies que desaparecen), en comparación con los monocultivos orgánicos (Marconi et al. 2022).
El presente trabajo busca describir (con mayor detalle que en anteriores trabajos) el cambio composicional de hierbas y de otras plantas espontáneas en los monocultivos convencionales, resaltando especies que suelen estar emparentadas con especies con poblaciones resistentes al paraquat y al glifosato, y evaluando cómo los cambios en el manejo se relacionan con los patrones temporales de presencia de algunas especies.
Este trabajo, al igual que las anteriores dos publicaciones (Marconi & Armengot 2020, Marconi et al. 2022), se realizó en la estación experimental de Sara Ana (15°27’36”S-67°28’17”O, 400 m) en Alto Beni. En esta estación se han realizado comparaciones de diversos aspectos agronómicos, grupos de biodiversidad y condiciones ambientales (ver lista de estudios en Ripa & Naoki 2021) entre sistemas agroforestales y monocultivos, que combinan tres tipos de manejo orgánico, convencional y sucesional. Las cuatro parcelas de monocultivos convencionales establecidas (cada una de 48 x 48 m) corresponden al manejo más intensivo dentro de la estación, donde se aplican agroquímicos y no existe ningún tipo de cobertura arbórea (salvo la sombra temporal de los plátanos que hubo en los primeros tres años).
Las estrategias de deshierbe han ido variando con los años (Fig. 1). Químicamente se aplicaron glifosato y paraquat, que son los herbicidas más utilizados en la producción de cacao a nivel mundial (Azembouh et al. 2021). Se rociaron 2.671 ± 1.094 g x ha-1 x año-1 de glifosato y 1.686 ± 686 g x ha-1 x año-1 de paraquat. Mecánicamente se trabajó lo equivalente a 7.6 ± 7.2 jornadas de ocho horas anuales por hectárea de deshierbe manual y 4.5 ± 3.4 usando moto- deshierbadora. Las diferentes actividades realizadas y herbicidas aplicados fueron categorizadas en cuatro periodos distintos (Fig. 1) En el primer periodo, 2010-2011, existió mayor trabajo manual y aplicación de otros herbicidas además de glifosato y paraquat: 2.4-D (3707g x ha-1), fomesafen (280 g x ha-1) y haloxyfop (156 g x ha-1). En el segundo periodo se aplicaron ambos herbicidas (paraquat y glifosato) y los esfuerzos fueron principalmente con moto-deshierbadora al igual que en el tercer periodo, 2015-2016, en el cual únicamente se aplicó Paraquat. En el cuarto periodo, 2017 en adelante, se usó prácticamente sólo glifosato y los tiempos laborales se redujeron en gran medida, posiblemente debido a la sombra y hojarasca que genera el cacao.
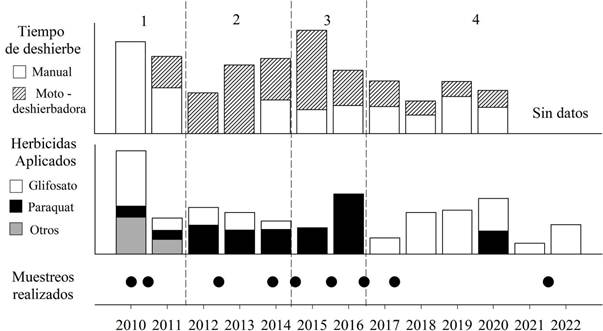
Figura 1. Herbicidas aplicados y actividades realizadas para el control de hierbas en monocultivos convencionales de cacao. En la parte inferior se muestran los herbicidas aplicados: glifosato en blanco, paraquat en negro y otros herbicidas en gris. En la parte superior las barras indican la cantidad de horas trabajadas por hectárea por persona. El patrón rayado implica el uso de moto-deshierbadora y el blanco, el deshierbe manual. Los valores fueron normalizados en base a sus respectivas medias y desviaciones estándar para su comparación. Los círculos en la parte inferior corresponden a cada uno de los muestreos de plantas espontáneas. Las líneas verticales en guiones y los números indican la clasificación del manejo en cuatro periodos distintos.
Se utilizaron datos de un total de nueve muestreos Braun-Blanquet de plantas espontáneas en el estrato herbáceo en los cuatro monocultivos convencionales que existen en el ensayo de Sara Ana (Fig. 1). Siete fueron la base del trabajo de Marconi et al. (2022) y adicionalmente se incluye el muestreo de junio-2014, que no fue tomado en cuenta antes, ya que solo se realizó en sistemas convencionales y enero-2022. Cada muestreo Braun Blanquet se realizó sobre una superficie de 256 m2 (16m x 16m), hacia el centro de cada una de las cuatro parcelas de monocultivos convencionales y utilizando los puntajes clásicos del método (Podani 2006).
Las diferencias de composición se analizaron con una ordenación no métrica de escalamiento multidimensional (Kindt & Coe 2005). Para conocer la importancia del tiempo y el sitio (parcela) para la composición se utilizó respectivamente un test de mantel y un análisis de similitudes, conocido como “anosim” en R (Kindt & Coe 2005). Adicionalmente se realizó un análisis permutacional multivariado de varianza utilizando matrices de distancia (“adonis.2” en R) para comparar la variación relacionada/explicada por ambos factores (Anderson 2001). En todos estos casos se utilizó el índice de disimilitud Bray-Curtis (Kindt & Coe 2005). Los análisis de composición se realizaron utilizando el paquete “vegan” de R, el lenguaje y entorno de computación estadística.
Para conocer qué especies se asociaban a un periodo en específico o a una combinación de estos se utilizó un test de especies indicadoras para grupos y combinaciones de grupos (De Cáceres et al. 2010). Este análisis se encuentra en el paquete “indicspecies” de R. Para todos los análisis de composición se utilizó una versión numérica de los puntajes de Braun Blanquet: “r”=1, “+”=2, “1”=3, “2”=4 y “3”=5, de tal manera que los datos se utilizaron a modo de escala, como lo sugiere Podani (2006). Por otro lado, para describir las abundancias de las especies y para calcular el grado de exclusividad dentro de los sistemas convencionales se realizó otro tipo de conversión de los puntajes Braun Blanquet, en este caso hacia valores de cobertura: “r” = 0.1%, “+”= 1%, “1”= 5%, “2”= 15% y “3”=37.5%. Este tipo de transformación es igual al realizado por la función “bb2tonum” del paquete “simba” de R.
Durante 12 años se registraron 172 especies distintas en los monocultivos convencionales, 77 corresponden a especies herbáceas o subarbustos y el resto son rebrotes o plántulas de especies leñosas (ver Anexo 1) A nivel de parcela se registraron 28 ± 9 especies de plantas espontáneas por parcela por año. Podemos observar que las especies más abundantes variaron entre los distintos periodos (Tabla 1). Por un lado, existen especies comunes en todo el ensayo de Sara Ana, como Lycianthes asarifolia y Euphorbiapoeppigii. Estas especies solo representan, respectivamente, 14 y 23% de la abundancia total (grado de exclusividad) de especies de los sistemas convencionales con aplicación de agroquímicos.
Tabla 1. Lista de cinco especies con mayor cobertura en cada uno de los periodos. El dato a continuación del nombre científico es el promedio de la cobertura, y entre paréntesis la cobertura máxima registrada en ese periodo. En negrita aparecen aquellas especies de las cuales más del 80% cuya suma de coberturas se encuentra dentro de sistemas con la aplicación de agroquímicos en el ensayo experimental de Sara Ana.
| 1 | 2 | 3 | 4 |
| Momordica charantia 13.63% (37.5%) | Leptochloa virgata 5.75% (375%) | Cyperus aggregatus 7.71% (37.5%) | Commelina obliqua 8.25% (15%) |
| Rottboellia cochinchinensis 3.5% (15%) | Boerhavia difusa 5.45% (37.5%) | Lycianthes asarifolia 5.33% (15%) | Pilea microphylla 5.25% (15%) |
| Euphorbia poeppigii 2.63% (15%) | Amaranthus viridis 4.6% (15%) | Leptochloa virgata 4.58% (15%) | Laportea aestuans 2.15% (15%) |
| Pombalia attenuata 2.51% (5%) | Euphorbia hirta 4% (15%) | Paspalum paniculatum 3.93% (15%) | Cyperus aggregatus 1.75% (5%) |
| Marsypianthes chamaedrys 2.5% (15%) | Lycianthes asarifolia 3.4% (5%) | Axonopus compressus 2.58% (15%) | Corchorus hirtus 1.53% (5%) |
La composición de plantas espontáneas de una parcela depende del tiempo (Mantel R = 0,66, P=0.001) y de la parcela (‘Anosim’ R = 0.28, P = 0.001). Estos factores son visibles en ambos ejes (stress= 0.20) de una manera ortogonal. El primer eje explica las diferencias en el tiempo (correlación Pearson entre NMDS1 y el tiempo: R= 0.92; Fig. 2) y el segundo eje explica las particularidades de cada parcela. Similarmente, encontramos que el tiempo explica el 53% de la variación de la composición y el tipo de parcela un 21%.
En el primer muestreo en total existen 58 especies de leñosas, que se van reduciendo linealmente a un ritmo de - 5.4 especies por año hasta llegar a ocho especies en 2022 (R2=0,81 P=0.0009, modelo lineal generalizado). Esto concuerda con el hecho de que 13 de las 22 especies asociadas al primer periodo son plántulas o rebrotes de especies leñosas comunes en los barbechos aledaños de acuerdo con la prueba de especies indicadoras. Algunos ejemplos de estas especies son Stigmaphyllon florosum (Malpighiaceae), Dolichandra uncata (Bignoniaceae), Guarea guidonia (Meliaceae) y Acalypha diversifolia (Euphorbiaceae). Entre las hierbas asociadas al primer periodo resaltan especies como Momordica charantia (Cucurbitaceae), Costus arabicus (Costaceae) y Bidens pilosa (Asteraceae).
En los posteriores años encontramos otras especies (Fig. 2, Tabla 1), como Leptochloa virgata (Poaceae), que es la segunda con mayor cantidad de registros a lo largo del tiempo en los monocultivos convencionales y la más abundante sumando todas las coberturas. En México, poblaciones de esta especie lograron tener resistencia al glifosato gracias a que disminuyeron la cantidad de glifosato que absorben, debido a que adquirieron una mutación en la enzima EPSPS y aumentaron la actividad de la vía metabólica en la que esta enzima participa (Alcántara de la Cruz et al. 2016).
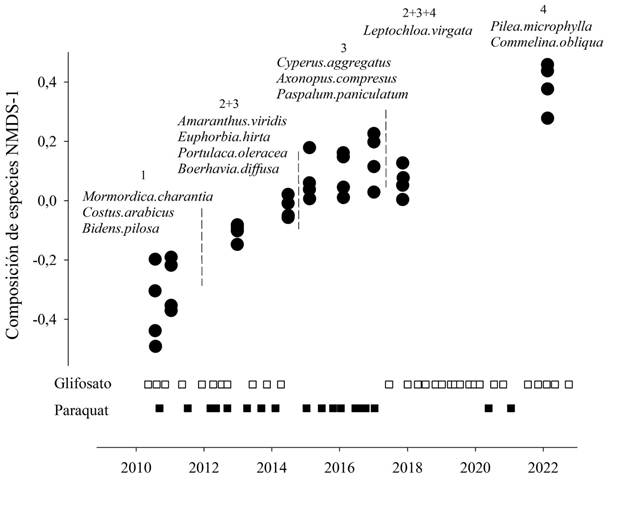
Figura 2. Relación entre la composición de especies y el tiempo. El eje vertical representa el primer eje de la ordenación no métrica de escalamiento muldimensional de la composición de plantas espontáneas en monocultivos convencionales. Cada redondo negro indica una parcela en un año específico. Las líneas verticales en guiones indican la clasificación del manejo en cuatro periodos distintos. Las especies dentro del gráfico son aquellas que obtuvieron un valor significativo para alguno de los periodos o combinación de estos en la prueba de especies indicadoras. Los cuadrados indican un evento de deshierbe en el tiempo donde se aplicó paraquat, cuadrados negros, o glifosato, cuadrados blancos.
Amaranthus viridis (Amaranthaceae) se asocia al segundo y tercer periodo de este estudio. Se sabe que recientemente se han encontrado poblaciones resistentes al glifosato de esta especie en Brasil (Alcántara de la Cruz et al. 2020). Poblaciones de otra especie en este género fueron controladas con paraquat, después de haber adquirido resistencia al glifosato, el cual elimina a la planta pero no su re-emergencia (Crow et al. 2015). El patrón de abundancia de Cyperus cf. aggregatus (Cyperaceae) muestra una meseta en el tercer periodo en el cual solo se aplicó paraquat. Esto se relaciona con estudios que han encontrado que otra especie del mismo género es fácilmente controlada por glifosato, pero no por paraquat (Iqbal et al. 2012).
Commelina obliqua (Commelinaceae) aumenta con el tiempo, pero tiene un leve descenso en su patrón de abundancia en los años de aplicación de paraquat. Se conoce que una especie relacionada, Commelina beghalensis, es poco controlada por glifosato, pero si puede ser eliminada por el paraquat (Maneechote et al. 2011). Por otro lado, C. obliqua no era en los primeros estudios una especie relacionada de manera exclusiva con los monocultivos convencionales. Tomando en cuenta los datos de todos los años, únicamente el 47% de su abundancia se encuentra dentro de los sistemas con aplicación de agroquímicos. El incremento dentro de estos es un evento reciente y quizá podría ser un indicio de un cambio en las poblaciones dentro de los cultivos convencionales. En otro estudio se encontró que el glifosato genera cambios estructurales en las ramificaciones de Commelina erecta (Panigo et al. 2012), los cuales se relacionan con cierta tolerancia al glifosato. En Sara Ana se observó que las formas de las hojas de Commelina obliqua varían entre las parcelas con aplicación de glifosato y aquellas sin su aplicación (Fig. 3); no obstante, es necesario un estudio específico para confirmarlo.
El trabajo en conjunto muestra que las especies que aparecen en cultivos de cacao con aplicación de herbicidas en Sara Ana son aquellas especies o parientes cercanos de estas que en otros países poseen poblaciones que han logrado resistencia o tolerancia al glifosato y paraquat. Este fenómeno de cambio cobra mayor importancia ya que la escala en la que este fenómeno ocurre en este estudio es pequeña (0.92 ha) e inmersa en una matriz de bosques secundarios y cultivos orgánicos. Consideramos importante, dentro de las actividades de promoción de la agricultura que existen, considerar la fuerza transformadora que poseen los herbicidas sobre los ecosistemas en los que se aplican.