











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkLos helechos arbóreos, al tener requerimientos abióticos específicos de humedad y temperatura, son indicadores del estado de salud y conservación de los bosques (Briones & Riaño 2014). Son ecológicamente importantes en el sotobosque porque pueden ser abundantes y afectar a la regeneración de los árboles, influenciar los ciclos de nitrógeno y fósforo, y proporcionar hábitats para el establecimiento de epífitas y hemiepífitas (Briones & Riaño 2014). Dos de las familias más comunes de helechos arbóreos en el Neotrópico son Dicksoniaceae y Cyatheaceae (PPG I 2016).
La familia Dicksoniaceae posee 40 especies en tres géneros: Colochalaena, Dicksonia y Lophosoria, de los cuales sólo Dicksonia y Lophosoria se distribuyen en Bolivia (Lehnert & Kessler 2018a). El género Dicksonia posee alrededor de 20 especies y está distribuido en bosques montanos húmedos y nublados de los trópicos, sobre pendientes o barrancos a lo largo de arroyos, entre 1.500 y 2.500 m de altitud, pudiendo llegar hasta los 3.500 m (Lehnert & Kessler 2018a). Dicksonia alcanza alturas de hasta 10 m, con tallos erectos, hojas con tricomas densos y muchas raíces fibrosas cercanas al ápice (Carreño- Rocabado 2000). También son características del género un manto denso, a veces corto en la base del pecíolo, con soros marginales (Lehnert & Kessler 2018a).
La familia Cyatheaceae incluye las especies de helechos arbóreos más altos, poseen troncos que pueden alcanzar hasta 20 m de altura, con escamas en la base de los pecíolos en lugar de pelos, lo que las distingue de las especies de Dicksoniaceae (Large & Braggins 2004). Se estima que Cyatheaceae incluye más de 600 especies distribuidas en el Neotrópico, en las montañas de Antillas Mayores, Centroamérica y América del Sur (Lehnert 2012, Lehnert & Kessler 2018b). Cyatheaceae posee una distribución cosmopolita, pero el género Cyathea es dominante principalmente en bosques montanos (Tryon 1976). En los Andes Centrales, Cyatheaceae presenta dos picos de riqueza a nivel altitudinal, uno a 1.000 m y otro a 2.100 m, y su diversidad decrece al aumentar la latitud (Tejedor 2017). Algunas especies de Cyathea colonizan áreas abiertas y promueven la regeneración de los bosques montanos (Arens & Baracaldo 1998). Las especies de Cyathea tienen diferentes preferencias ecológicas, desde ambientes abiertos y cálidos hasta hábitats muy húmedos y sombreados; algunas especies están adaptadas a hábitats muy específicos como dunas de arena o fondos de quebradas (Lehnert & Kessler 2018b).
Los helechos arbóreos son elementos muy comunes en los bosques montanos de los Yungas bolivianos (Navarro 1997), incluyendo varias especies de Dicksonia y Cyathea, entre ellas Dicksonia karsteniana (Alfonso-Moreno et al. 2011). D. karsteniana es una especie principalmente andina que habita en bosques de montaña desde el sur de Venezuela hasta la parte central de Bolivia (Noben et al 2018). D. karsteniana fue previamente considerada como una variedad de D. sellowiana, pero actualmente se considera como una especie separada (Noben et al. 2018). Las especies de Cyathea y Dicksonia están amenazadas por la fragmentación de su hábitat a causa del cambio de uso del suelo y la comercialización de sus partes, principalmente del tallo (Mallmann et al. 2019). En el caso de especies sudamericanas del género Dicksonia, todas están listadas por la convención CITES (Martins 2009, Lehnert & Kessler 2018a).
Las diferentes especies de helechos arbóreos pueden diferir en su tolerancia a la sombra, requisitos de fertilidad de suelos, requerimientos de humedad, lo cual se puede reflejar en diferentes patrones espaciales. El análisis de los patrones espaciales es una herramienta muy útil en el entendimiento de los procesos que determinan el establecimiento y desarrollo de los individuos en un ambiente dado, y son el resultado de procesos que promueven la coexistencia de las especies (Salinas-Melgoza 2016). Por ejemplo, un patrón de distribución agregado puede resultar de variaciones edáficas relacionadas con la topografía, variaciones en la disponibilidad de luz o una dispersión de semillas espacialmente limitada (Traissac & Pascal 2014).
Los estudios de distribución espacial de especies de helechos arbóreos en bosques húmedos montanos y su relación con variables ambientales son escasos. La presente nota contribuirá a mejorar la comprensión de los patrones de distribución de estos taxa, con alto interés en conservación y ecología del paisaje (Antúnez et al. 2022). En este contexto, el objetivo de este trabajo es comparar la densidad y distribución de Dicksonia karsteniana con especies de Cyathea en un bosque yungueño, para contribuir al conocimiento de su estado poblacional.
El estudio se llevó a cabo en la comunidad de Pataloa, colindante a la Estación Biológica Santiago de Chirca, ubicada en la provincia Sud Yungas del departamento de La Paz, Bolivia (16°23'57,19" S - 67°34'29,91" W). La zona presenta parches de bosques montanos bien conservados dominados por Lauraceae y Melastomataceae, bosques secundarios y vastas zonas degradadas por fuego. El muestreo se realizó en un parche de bosque primario que cubre aproximadamente 300 ha y atraviesa el camino precolombino Yunga Cruz, a una elevación de 2.240 m (Fig. 1a).
Para determinar los patrones espaciales se aplicó el método de cuadrantes o parcelas contiguas (Krebs 1998). Se establecieron 20 parcelas de 25 x 8 m a lo largo del sendero principal que atraviesa el área de estudio, cubriendo un área de 4000 m2. Las parcelas se colocaron de forma continua, para cubrir todo el parche de bosque con el fin de observar la distribución de ambos géneros. En cada parcela se muestrearon todos los helechos arbóreos mayores a 1.5 m de altura. Entre los helechos muestreados, se comparó Dicksonia karsteniana con diferentes especies de Cyathea. No se registraron otros géneros de helechos arbóreos en las parcelas de muestreo. Para cada individuo, se registró el punto sobre los 25 m donde fue observado (Fig. 1b). En cada parcela se tomaron datos de la inclinación, exposición y distancia al arroyo que se encontraba en el fondo de la quebrada. Además, en tres puntos a lo largo de cada parcela se midió la profundidad de la hojarasca y se tomaron muestras de suelo superficial (inmediatamente por debajo de la hojarasca), obteniéndose un total de 60 muestras de suelo, durante el mismo día. De cada muestra de suelo se obtuvo el peso húmedo, peso seco (después de secar la muestra por 10 días a temperatura ambiente) y se calculó el porcentaje de humedad del suelo.
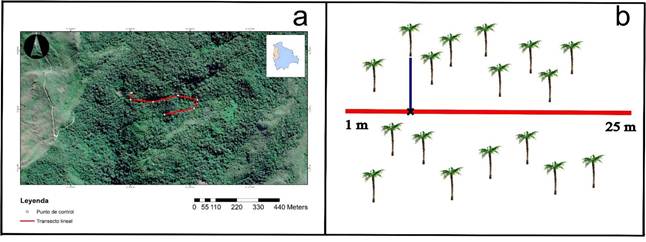
Figura 1. Localización del sitio. a. Ubicación del sendero y puntos de muestreo, b. método de cuadrantes contiguos, donde la X representa el punto registrado para cada helecho sobre la línea de 25 m.
La densidad de helechos se calculó a partir del número total de individuos de cada género, dividido entre el área total de la parcela (4.000 m2). Para analizar el efecto de las variables abióticas en la abundancia de los dos géneros, se utilizaron modelos lineales generalizados, para cada género por separado, suponiendo una distribución de errores Poisson para las variables dependientes. Para analizar el patrón de distribución espacial de cada género se calculó el índice de dispersión de Morisita (1962), que varía entre 0 y n: en patrones de distribución uniforme se obtiene un valor entre 0 y 1, en patrones agregados se obtiene un valor entre 1 y n, y en patrones aleatorios I d = 1. Para analizar si existe segregación entre los dos géneros se calculó el C-score y se lo comparó con una distribución aleatoria de la comunidad obtenida mediante 1000 simulaciones (Begon et al. 2006). Todos los análisis se realizaron en la plataforma R, versión 4.1.0 (R Core Team 2021).
La densidad de helechos se calculó a partir del número total de individuos de cada género, dividido entre el área total de la parcela (4.000 m2). Para analizar el efecto de las variables abióticas en la abundancia de los dos géneros, se utilizaron modelos lineales generalizados, para cada género por separado, suponiendo una distribución de errores Poisson para las variables dependientes. Para analizar el patrón de distribución espacial de cada género se calculó el índice de dispersión de Morisita (1962), que varía entre 0 y n: en patrones de distribución uniforme se obtiene un valor entre 0 y 1, en patrones agregados se obtiene un valor entre 1 y n, y en patrones aleatorios I d = 1. Para analizar si existe segregación entre los dos géneros se calculó el C-score y se lo comparó con una distribución aleatoria de la comunidad obtenida mediante 1.000 simulaciones (Begon et al. 2006). Todos los análisis se realizaron en la plataforma R, versión 4.1.0 (R Core Team 2021).
En el caso de Cyathea la inclinación y la distancia al arroyo tuvieron efectos positivos en su abundancia; es decir, a mayor inclinación y mayor distancia al arroyo, mayor densidad de Cyathea (Tabla 1). La afinidad de Cyathea por terrenos inclinados podría estar relacionada con su preferencia por sitios con mayor incidencia de luz (Lehnert & Kessler 2018b). Los sitios con mayor inclinación suelen tener mayor apertura de dosel, lo cual implica mayor penetración de luz al suelo y resulta en mayor estrés hídrico (Dignan & Bren 2003). La relación positiva de la abundancia de Cyathea con la distancia al arroyo coincide con lo hallado para C. delgadii, que tiene preferencia por sitios con mayor radiación solar y temperatura en bosques atlánticos subtropicales y del NE de Brasil (Dos Santos-Silva et al. 2019, Schwartz & Gasper 2020). Sin embargo, es importante considerar que las diferentes especies muestreadas de Cyathea probablemente tienen requerimientos microclimáticos distintos; por lo tanto, se recomiendan más estudios sobre los patrones espaciales y afinidades abióticas de Cyathea a nivel de especie.
Tabla 1. Tablas de selección de modelos para evaluar los efectos de la humedad del suelo, inclinación, profundidad de la hojarasca y distancia a un arroyo en la abundancia de Dicksonia y Cyathea obtenidas mediante modelos lineales generalizados con distribución Poisson. Leyenda: In = Intercepto, DA = distancia arroyo, HS = humedad suelo, Ic. = inclinación, PH = profundidad hojarasca, gl = grados de libertad
| In | DA | HS | Icl | PH | gl | log link | AICc | Delta AICc | Peso |
|---|---|---|---|---|---|---|---|---|---|
| Dicksonia | |||||||||
| -12.91 | -0.034 | 0.282 | 0.274 | 4 | -11.39 | 33.4 | 0 | 0.853 | |
| -12.11 | -0.037 | 0.296 | -0.034 | 0.282 | 5 | -11.34 | 37 | 3.5 | 0.147 |
| 1.16 | 1 | -162.29 | 326.8 | 293.4 | 0 | ||||
| Cyathea | |||||||||
| -2.52 | 0.004 | 0.076 | 3 | -52.57 | 112.6 | 0 | 0.617 | ||
| -3.36 | 0.004 | 0.018 | 0.083 | 4 | -52.09 | 114.8 | 2.2 | 0.204 | |
| 1.25 | 1 | -64.48 | 131.2 | 18.6 | 0 | ||||
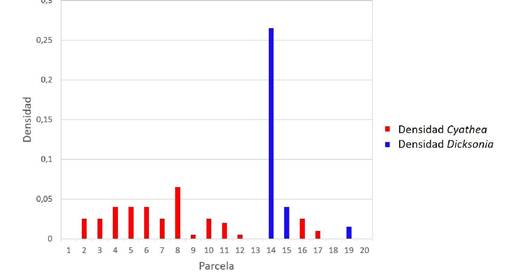
Al analizar los patrones de distribución, se obtuvo un patrón fuertemente agregado para Dicksonia (13.97; Fig. 2) y agregado para Cyathea (1.81). Ambos valores fueron significativamente diferentes al modelo nulo con distribución al azar (x 2, p<0.0001). La distribución altamente agregada de Dicksonia, puede ser explicada por sus requerimientos abióticos, que coinciden con la elevada humedad del suelo (Hunt et al. 2002), causada en nuestro caso por la cercanía al arroyo. Nuestras observaciones coinciden con el estudio de Martins (2009) en la Mata Atlántica, donde se vieron patrones de colonias agregadas de D. sellowiana en llanuras y pendientes húmedas; estas agrupaciones se atribuyeron a la acumulación de sedimentos y humedad. La estructura espacial agregada en D. karsteniana también coincide con lo reportado para D. sellowiana en bosques de Araucaria en Brasil (Marques & Krupek 2014, Lerner et al. 2021).
El valor promedio de C-score de estos géneros fue significativamente menor a lo esperado por azar (C- score=39 ±4.1, p = 0.033), lo que indica que existe segregación entre estos géneros de helechos en nuestro sitio de estudio. Esto sugiere que Dicksonia y Cyathea no coexisten en un mismo sitio a escala fina en nuestra área de estudio, lo cual coincide con lo reportado por Dignan & Bren (2003) para D. antarctica y C. australis en bosques australianos. Según Volkova et al. (2009), D. antarctica y C. australis tienen diferentes preferencias a nivel de micrositios, relacionadas con sus diferencias morfológicas y de capacidad fotosintética. Lehnert & Kessler (2018a) también indican que la presencia de Dicksonia está relacionada con alta cobertura de dosel, elevada humedad y baja exposición a la luz. Sin embargo, se necesitan más estudios para corroborar la agregación y segregación encontradas en el presente trabajo.
En este estudio comparamos la densidad y patrones de distribución de Dicksonia karsteniana con varias especies de Cyathea. La densidad de D. karsteniana fue similar a la alcanzada por varias especies de Cyathea en el sitio de estudio. Así mismo, nuestros resultados destacan el patrón de distribución fuertemente agregado de D. karsteniana, con una concentración del 95% de los individuos en solo dos parcelas (10% del área muestreada, Fig. 2). Dicho patrón estuvo relacionado positivamente con la humedad del suelo y la profundidad de la hojarasca y negativamente con la distancia a un arroyo. Nuestros resultados destacan la necesidad de conservar los ambientes cercanos a los arroyos para mantener las poblaciones de esta especie.

