











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkIntroducción
La producción de estructuras reproductivas (flores y frutos) es una etapa fundamental en el ciclo de vida de las plantas ya que determinará la demografía de la población (Ratsirarson et al. 1996, Quitete-Portela et al. 2010, Lopez-Toledo et al. 2011). Para varias especies de plantas se ha encontrado una variación en el patrón temporal de producción de flores y frutos a lo largo de la vida (Sampaio & Scariot 2008). Los factores que condicionan los períodos de floración y fructificación en muchas especies de las plantas están relacionados con la temperatura y las precipitaciones (Morellato et al. 2013, Pau et al. 2013). En el caso de las palmeras, familia Arecaceae, muchas especies se encuentran en ambientes de humedales con una distribución estrechamente relacionada con el régimen de precipitaciones (Bjorholm et al. 2005, Dransfield et al. 2008). En estos ambientes de humedales las palmeras forman paisajes característicos y tienen adaptaciones que garantizan el éxito reproductivo (Lasso et al. 2016). Entre estas adaptaciones, las palmeras tienen la floración en la estación húmeda y una fructificación extendida con un pico en la estación seca, incrementando la disponibilidad de frutos para los frugívoros en una época en que las otras especies de plantas no suelen fructificar (Ragusa-Netto & Fecchio 2006, Keuroghlian & Eaton 2008, Araujo & Lobo 2020).
Copernicia alba Morong. conocida comúnmente como “palma blanca” o “caranday” (Cabral & Castro 2007), pertenece a la familia Arecaceae, subfamilia Coryphoideae (Moraes 2006) y se presenta tanto en bosques como en pastizales, donde los individuos maduros ocupan el estrato superior (Cisz 2011). Esta palmera se extiende desde el norte de Argentina, este-noreste de Paraguay, oeste de Brasil (Pantanal) y norte y este de Bolivia (Costa et al. 2022). La especie ocupa la ecorregión chaqueña en planicies aluviales inundables donde se dan ciclos entre inundaciones y largos períodos de sequía (Moraes 1991, Negrelle & Degen-Naumann 2012), resistiendo menos de 300 mm de precipitaciones por año (Moraes 2001) e incendios frecuentes en la estación seca (Torres et al. 2015). La polinización de C. alba es realizada por abejas (Moraes 1991, Cisz 2011) y por el viento (Mereles 2001).
En los diferentes países donde C. alba se encuentra distribuida, se han descrito períodos de floración y fructificación diferentes con respecto a la duración y los meses en los que ocurre, así como para algunas características de los individuos tales como el tamaño (Moraes 1991, Ragusa-Netto & Fecchio 2006, Mereles 2001). Los frutos de C. alba son un recurso clave para especies de fauna nativa (por ejemplo, Rhea americana Linnaeus, "Tayassu pécari Link y Pécari tajacu Linnaeus [Puechagut et al. 2013]) y como forraje para ganado (Castellani et al. 2014), por lo tanto, es importante contar con información de base para realizar un manejo adecuado de este recurso para la conservación de la fauna silvestre nativa. Además, es necesario determinar la relación entre las precipitaciones y la temperatura en la producción de estructuras reproductivas de C. alba considerando el actual contexto de cambio climático (Walther et al. 2007, Eiserhardt et al. 2011, Nazareno & Reis 2012). Específicamente en este trabajo se pretende describir el patrón temporal, la producción de flores y frutos y el número de frutos producidos por individuo de acuerdo al tamaño de las palmeras, las precipitaciones y la temperatura en el noroeste argentino.
Area de estudio
Este estudio se realizó en un palmar cercano a la localidad de El Talar, Provincia de Jujuy, Argentina (23°39.107’S, 64°20.522’O). Este palmar pertenece a la empresa Ledesma S.A.A.I., ocupa una superficie de 656 ha, no ha tenido fuegos frecuentes y se encuentra rodeado por un alambrado que excluye el ganado. Las precipitaciones anuales en esta zona varían entre los 650 y 750 mm y se concentran entre los meses de noviembre y abril (Bravo et al. 1999). Además de C. alba, otras especies vegetales dominantes son Geoffroea decorticans, Prosopis ruscifolia y Tabebuia nodosa (Chalukian et al. 2002).
Métodos
En el palmar estudiado se seleccionaron al azar 50 palmeras con una altura > 3 m. Cada palmera se marcó con una chapa de aluminio numerada. Se midió la altura de cada palmera desde el suelo hasta la base de la copa con un palo telescópico graduado de 15 m. Para describir el patrón temporal de floración y fructificación se determinó visualmente, con ayuda de binoculares, la presencia o ausencia de inflorescencias y/o infrutescencias de las palmeras marcadas durante 19 meses (entre septiembre de 2011 y septiembre de 2013, sin embargo, no se registraron datos en enero, marzo, abril y octubre de 2012 ni en enero y febrero de 2013 por problemas logísticos). Para contar el número de frutos producidos por individuo, se tomó una fotografía de cada palmera marcada durante los 19 meses de seguimiento. Las fotografías fueron tomadas para que incluyeran todas las infrutescencias en el marco de la foto, observando que la luz solar no interfiriera. En gabinete se superpuso una grilla de 2.500 subdivisiones sobre cada fotografía y se contaron los frutos maduros, en cada subdivisión se marcaron los frutos para evitar contar dos veces los mismos frutos. Es importante destacar que el número de frutos reportado no es la cantidad producida por mes sino la cantidad cuantificada cada mes, los mismos frutos en la misma palmera pueden haber sido contados en meses sucesivos.
Se comparó la proporción de individuos que florecieron o fructificaron (variables dependientes) a lo largo de todo el seguimiento (mes contra mes, variable predictora como factor fijo de 19 niveles), entre los dos años de muestreo: Año 1 (septiembre de 2011 - agosto de 2012) y Año 2 (septiembre de 2012 - agosto de 2013). Se utilizó el año como factor fijo con dos niveles (año 1 y año 2), y entre los dos años de seguimiento dentro de cada mes, se consideró únicamente los siete meses en los que se tomaron datos en los dos años. Se utilizaron modelos lineales generales y mixtos y el test a posteriori DGC en Infostat para determinar si existen diferencias de las variables dependientes entre meses (Di Rienzo et al. 2011, 2012, 2015). Siempre se consideró los números identificadores de cada palmera como variable aleatoria, ya que el conteo se realizó mensualmente en las mismas palmeras (Di Rienzo et al. 2011, 2012, 2015). Para determinar si existían diferencias entre meses dentro de cada año de seguimiento (factor fijo con 19 niveles), se comparó el promedio del número de frutos contados por mes (variable dependiente) utilizando modelos lineales generales y mixtos y el test a posteriori DGC, incluyendo únicamente a todas las palmeras que produjeron frutos (n= 23 palmeras) y al número identificador de cada palmera como factor aleatorio, ya que el conteo de individuos se realizó mensualmente en los mismos individuos (Di Rienzo et al. 2012, 2014).
Los datos de precipitaciones y temperatura se obtuvieron de la estación meteorológica de la empresa Ledesma S.A.A.I., localizada en el área de estudio. Se obtuvieron las precipitaciones diarias y las temperaturas máximas y mínimas diarias entre septiembre de 2011 y septiembre de 2013 (25 meses). Con estos datos se calcularon las precipitaciones mensuales, la temperatura mensual y la amplitud térmica mensual durante el tiempo de seguimiento. Para evaluar si hubo variaciones entre los 25 meses que duró el estudio en las precipitaciones mensuales, la temperatura mensual y la amplitud térmica mensual, se compararon las variables climáticas entre meses utilizando modelos lineales generales y mixtos con los meses como factor fijo con 12 niveles (Di Rienzo et al. 2011, 2012, 2015).
Para evaluar la relación entre la probabilidad de producir inflorescencias o infrutescencias y el tamaño de las palmeras marcadas, los datos de precipitaciones y de temperatura, se ajustaron modelos de regresión logística con la variable respuesta presencia o ausencia de inflorescencias/infrutescencias y enlace logit utilizando la plataforma de modelos lineales generalizados mixtos de Infostat (Di Rienzo et al. 2011, 2012, 2015). Para evaluar la relación entre la cantidad de frutos contados por mes y el tamaño de las palmeras marcadas, los datos de precipitaciones y de temperatura, se ajustaron modelos de regresión Poisson con función de enlace log utilizando la plataforma de modelos lineales generalizados mixtos de Infostat (Di Rienzo et al. 2011, 2012, 2015). El tamaño de las palmeras, las precipitaciones mensuales, la temperatura mensual y la amplitud térmica mensual son los efectos fijos del modelo y los efectos aleatorios en el modelo es el número identificador de cada palmera, el año y el mes de seguimiento anidado dentro del año ya que las observaciones se realizaron sobre los mismos individuos.
Resultados
En el primer año florecieron 30 palmeras de las 50 palmeras marcadas y en el segundo año florecieron 33 palmeras, 26 de las cuales (52% del total de palmeras marcadas) florecieron los dos años. Sin embargo, 15 palmeras (30%) no florecieron en ninguno de los dos años. No se encontraron diferencias significativas entre los meses del primer y segundo año de seguimiento en la proporción mensual de palmeras marcadas que florecieron (Año 1: 0.11±0.02 y Año 2: 0.14±0.02; F= 1.76; P= 0.18).
En los meses de mayo, junio, julio y agosto no se registraron palmeras con flores en ninguno de los dos años. La proporción de individuos con inflorescencias varió entre meses marcándose dos picos (octubre y noviembre de 2011, y noviembre y diciembre de 2012) a lo largo del tiempo de seguimiento (F= 26.59; P< 0.05; Fig. 1).
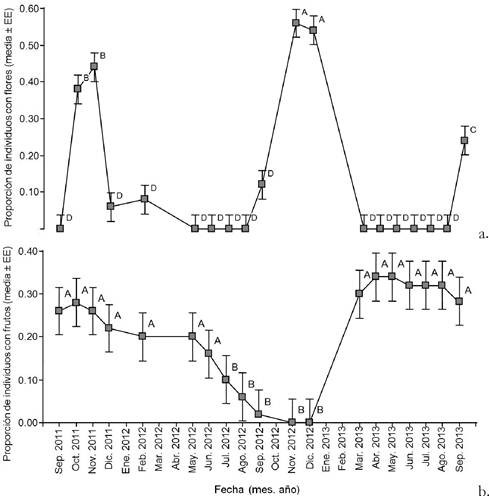
Figura 1. Proporción de individuos con flores (a.) o frutos (b.) en 50 palmeras marcadas y seguidas desde septiembre de 2011 a septiembre de 2013 en un palmar de Copernicia alba cercano a la localidad de El Talar, provincia de Jujuy, Argentina. Letras distintas indican diferencias significativas entre meses (P<0.05).
Se registraron palmeras con frutos en todos los meses de seguimiento, excepto en noviembre y diciembre de 2012. En el primer año fructificaron 16 palmeras, en el segundo año fructificaron 18 de las 50 palmeras, 11 de las cuales (22% del total de palmeras marcadas) fructificaron los dos años. Sin embargo, 27 palmeras (54% del total) no fructificaron en ninguno de los dos años. Considerando a las 30 palmeras que florecieron en el primer año, 22 de ellas llegaron a producir frutos en alguno de los dos años de seguimiento. No se encontraron diferencias significativas entre el primer y segundo año de seguimiento en la proporción mensual de palmeras marcadas que fructificaron (Año 1: 0.19±0.02 y Año 2: 0.22±0.02; F= 0.82; P= 0.36). La proporción de individuos con infrutescencias varió entre meses, presentándose un período (de julio a diciembre de 2012) con menor proporción de palmeras con frutos (F= 4.56; P< 0.05; Fig. 1).
Durante el tiempo de seguimiento de las 50 palmeras se observó un período con significativamente mayores precipitaciones (estación húmeda) y un período con significativamente menores precipitaciones (estación seca; F= 2.55; P< 0.05; Fig. 2). Las precipitaciones mensuales durante la estación húmeda en el primer año de seguimiento (109.42±39.85 mm/mes) fueron significativamente mayores a las registradas en el segundo año de seguimiento (54.58±39.85 mm/mes; F= 3.20; P< 0.05). La temperatura durante el tiempo de seguimiento fue de 22.73±0.21°C y se observó un período en que las temperaturas fueron significativamente mayores, coincidiendo aproximadamente con la estación húmeda (F= 87.84; P< 0.05; Fig. 2). La amplitud térmica diaria durante el tiempo de seguimiento fue de 14.55±0.18°C y durante la estación seca se registraron tres meses con mayor amplitud térmica (de julio a septiembre) y el resto de los meses con menor amplitud térmica (F= 11.59; P< 0.05) durante el tiempo de seguimiento.
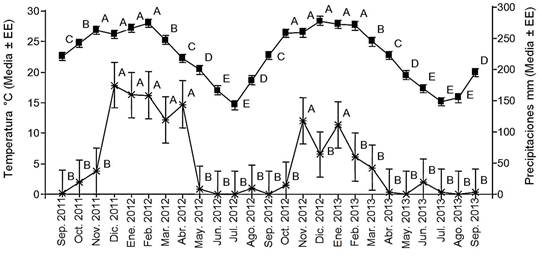
Figura 2. Temperatura (cuadrados) y precipitaciones (asteriscos) mensuales entre septiembre de 2011 y septiembre de 2013 en un palmar cercano a la localidad de El Talar, provincia de Jujuy, Argentina. Los datos fueron proporcionados por la empresa Ledesma S.A.A.I. Letras diferentes indican diferencias significativas entre meses (P<0.05).
La probabilidad de que las palmeras produzcan estructuras reproductivas estuvo relacionada con el tamaño (es decir, la altura) de la palmera y con la temperatura (Tabla 1; Fig. 3). Ninguno de los 11 individuos marcados que tenían una altura entre 3 y 5 m florecieron o fructificaron durante los dos años del seguimiento. No se registró floración por debajo de la temperatura de 22.57°C (percentil 50). No se encontró relación entre estructuras reproductivas de las palmeras y las precipitaciones acumuladas, ni con la amplitud térmica, durante el tiempo de seguimiento (Tabla 1). Se encontró una relación positiva significativa entre el número de frutos contados por palmera por mes y el tamaño de cada palmera (z= 3.53; P< 0.05) y a medida que disminuye la temperatura media mensual es mayor el número de frutos contados por palmera (z= -2.86; P< 0.05; Fig. 4). No se encontró una relación entre el número de frutos por palmera y las precipitaciones mensuales (z= - 0.78; P= 0.43), ni con la amplitud térmica (z= -1.53; P= 0.12), durante el tiempo de seguimiento.
Discusión
De acuerdo a los resultados de este trabajo, el período de floración de C. alba es sincrónico y está relacionado positivamente con la temperatura mensual. Un patrón similar ha sido encontrado en otras especies de palmeras, en las que la producción de flores se encuentra concentrada en la estación húmeda y cálida (Gorman 1996, Sampaio & Scariot 2008, Genini et al. 2009, Rocha et al. 2015, Garcia & Barbedo 2016). En general, el aumento de temperatura es el principal factor que influye positivamente en la producción de flores al aumentar la tasa de biosíntesis y procesos metabólicos (Raich et al. 1997, Pau et al. 2013).
El pico de floración de C. alba en el palmar estudiado se da algunos meses después que lo observado en el Pantanal brasileño (agosto y septiembre; Ragusa-Netto & Fecchio 2006) y el noreste de Bolivia (agosto a octubre; Moraes 1991). Esto puede deberse a variaciones climáticas en los diferentes puntos de la distribución de C. alba, ya que en el noreste boliviano la temperatura media mensual varía entre 24.3 y 27.9°C y la media anual está por encima de los 26°C y en el Pantanal brasileño la temperatura media varía entre 20°C en la estación seca y 27°C en la estación húmeda (Moraes 1991, Ragusa-Netto & Fecchio 2006). En el sudoeste de Paraguay, en donde la temperatura media anual es similar a la observada en el área de estudio, con una media anual de 23-24°C se observó el mismo período de floración que en el área de estudio (Mereles 1999, 2001).
Tabla 1. Efecto del tamaño de las palmeras, la precipitación y la temperatura registradas durante el periodo de seguimiento sobre la probabilidad de floración y fructificación de 50 palmeras marcadas y seguidas desde septiembre de 2011 a septiembre de 2013 en un palmar de Copernicia alba en el noroeste de Argentina. En negrita se muestran los resultados significativos.
| Floración | Fructificación | |||
|---|---|---|---|---|
| Variable (tiempo seguimiento) | z | P | z | p |
| Tamaño (m) | 4.26 | <0.01 | 4.73 | <0.01 |
| Precipitaciones acumuladas (mm) | -0.33 | 0.74 | 0.17 | 0.87 |
| Temperatura (°C) | 3.00 | <0.01 | -1.93 | 0.05 |
| Amplitud térmica (°C) | 1.37 | 0.17 | -1.76 | 0.08 |
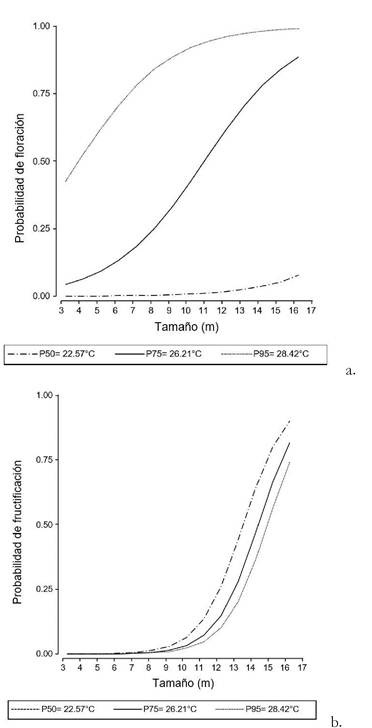
Figura 3. Probabilidad de floración (a) y de fructificación (b) en relación al tamaño de 50 palmeras marcadas y seguidas entre septiembre de 2011 y septiembre de 2013 en un palmar de Copernicia alba cercano a la localidad de El Talar, provincia de Jujuy, Argentina. Cada curva representa una temperatura, para los percentiles 50 (línea discontinua), 75 (línea continua) y 95 (línea punteada) de la distribución de temperaturas.
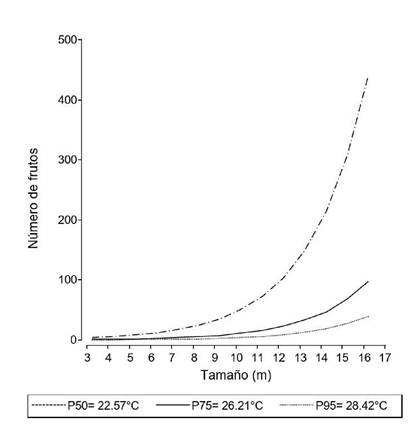
Figura 4. Número de frutos contados por mes en relación al tamaño de 23 palmeras marcadas y seguidas entre septiembre de 2011 y septiembre de 2013 en un palmar de Copernicia alba cercano a la localidad de El Talar, provincia de Jujuy, Argentina. Cada curva representa una temperatura, para los percentiles 50 (línea discontinua), 75 (línea continua) y 95 (línea punteada) de la distribución de temperaturas.
Durante el desarrollo de este estudio se encontró frutos en las palmeras en prácticamente todos los meses de seguimiento. Cuando se considera la distribución geográfica de la especie se observa que en otras regiones la fructificación también ocurre extendida por varios meses (Mereles 1999, 2001, Cisz 2011). En el Chaco paraguayo, el pico de fructificación es de febrero a agosto y los frutos caen a fines del invierno (Mereles 1999, Cisz 2011). Para el pantanal de Brasil se ha reportado que C. alba fructifica desde septiembre a mayo, coincidiendo con el período de temperaturas más altas (Ragusa Netto & Fecchio 2006, Araujo & Lobo 2020), y en el noreste de Bolivia la fructificación se da desde octubre hasta mayo (Moraes 1991). La prolongada fructificación en C. alba puede deberse, como ocurre en otras especies de palmeras, que los frutos requieren un largo período de desarrollo (Khorsand Rosa et al. 2013, Pedroso et al. 2021). La fructificación de C. alba en este estudio mostró una relación inversa con la temperatura lo cual puede deberse a que altas temperaturas producen desecamiento de los frutos impidiendo la adecuada maduración o absición prematura de los frutos (Staggemeier & Morellato 2011, Cortés-Flores et al. 2013, Tisné et al. 2020). En el pantanal de Brasil, la fructificación no mostró correlación con variables climáticas, pero los frutos inmaduros mostraron una asociación positiva con la temperatura (Araujo & Lobo 2020). Sin embargo, dado que en este estudio no se evaluaron las distintas fenofases de la fructificación es difícil interpretar este patrón (Cortés-Flores et al. 2019). Es necesario resaltar que en este estudio menos de la mitad de las palmeras marcadas fructificaron en los dos años de seguimiento, esto podría sugerir serias limitaciones en la estrategia reproductiva de C. alba en el noroeste argentino (Khorsand Rosa & Koptur 2013).
La probabilidad de floración y fructificación y la cantidad de frutos de C. alba estarían fuertemente influenciadas por el tamaño de la palmera, incrementando la producción de estructuras reproductivas a medida que aumenta de tamaño el individuo. Estudios sugieren que en C. alba la actividad reproductiva aumenta durante todo el ciclo de vida de la palmera (Oyama 1990, Mendoza & Franco 1998, Svenning 2001, Berry & Gorchov 2004). A su vez, los individuos de mayor tamaño estarían más expuestos a ser polinizados por el viento y por los insectos, por lo que producirían más frutos que los que son de menor tamaño y se encuentran por debajo del dosel (Svenning 2001). En un palmar en Beni, Bolivia, se encontró que C. alba comienza a mostrar estructuras reproductivas cuando alcanzan los 3 m de tamaño (Moraes 1991) y en dos estudios sobre la especie realizados en Paraguay se consideran adultos (por presentar estructuras reproductivas) a todos los individuos de más de 3 m de tamaño (Cisz 2011, Degen & Negrelle 2014). Aunque pareciera que en el área de estudio C. alba alcanza la madurez reproductiva con un tamaño mayor (>5 m para flores y >10 m para frutos) que en otros lugares donde la palmera se distribuye, no se han encontrado estudios analizando diferencialmente la floración y fructificación en relación al tamaño de los individuos.
Conclusiones
Los resultados de este trabajo muestran que en el noroeste argentino el período de floración de C. alba es sincrónico y menos de la mitad de las palmeras marcadas fructificaron en los dos años de seguimiento. La probabilidad de floración y fructificación y la cantidad de frutos de C. alba en el noroeste argentino incrementa a medida que aumenta de tamaño el individuo. La floración de C. alba en el noroeste argentino está relacionado positivamente con la temperatura mensual, mientras que la fructificación y la cantidad de frutos presentan una relación inversa con la temperatura. Es fundamental continuar con estudios fenológicos a largo plazo que permitan evaluar la respuesta de C. alba al aumento de la temperatura debido al cambio climático (Morellato et al. 2016).

