











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkEcología en Bolivia
versión impresa ISSN 1605-2528versión On-line ISSN 2075-5023
Ecología en Bolivia vol.56 no.2 La Paz set. 2021
Artículo
Variación en rasgos reproductivos de roedores silvestres a diferente elevación en los Andes bolivianos
Reproductive traits variation in wild rodents along an elevational gradient in the Bolivian Andes
Susana G. Revollo-Cadima1,2*, Adriana Rico-Cernohorska2 & Jorge Salazar-Bravo2,3,4
1International Center for Arid and Semi Arid Land Studies, Texas Tech University, Lubbock, TX 79409-5004, USA
2Colección Boliviana de Fauna, Instituto de Ecología, Carrera de Biología, Universidad Mayor de San Andrés, Campus Universitario, Calle 27 Cota Cota, Casilla 10077-Correo Central, La Paz, Bolivia
3Department of Biological Sciences, Texas Tech University, Lubbock, TX 79409-3131, USA
4Instituto Nacional de Biodiversidad, Quito, Ecuador.
*Autora de correspondencia: sussygab@gmail.com
Recibido: 01.11.20, Aceptado: 11.03.21.
Resumen
En varias especies de roedores, la reproducción está regulada por las características del ambiente. En este sentido, la elevación puede ser una variable importante, ya que a mayor elevación los recursos fluctúan con una mayor tendencia a la estacionalidad. Por ello, se espera que la reproducción se ajuste a estas condiciones, siendo más estacional a mayor elevación por el costo energético que representa. En esta contribución ponemos a prueba esta hipótesis, analizando los patrones reproductivos (tamaño de camada, número de embriones, largo testicular en adultos) en dos géneros de roedores sigmodontinos que se distribuyen a lo largo de la Coordillera Oriental en Bolivia: Akodon y Oligoryzomys. Los datos fueron analizados con un Modelo Lineal Generalizado (GLM). Para ambos géneros se tuvo mayor cantidad de hembras preñadas en época de transición (Akodon: χ2=6.26, g.l.= 1, p = 0.01; Oligoryzomys: χ2=8.10, g.l.= 1, p = 0.004). El tamaño testicular en Akodon en época de transición fue mayor a mayor elevación (W = 239, p = 0.02), mientras que en Oligoryzomys fue mayor a menor elevación (W=256, p ≤ 0.01). El tamaño de la camada, sólo en Oligoryzomys, fue más grande en época seca a menor elevación (W=60.5, p = 0.01). La elevación y la época del año afectan los patrones reproductivos y su efecto varía según los grupos estudiados. Estas diferencias deben estar relacionadas a la plasticidad de las especies en respuesta a la variación intrínsica de los recursos alimenticios con la estación y en función de los requerimientos energéticos especie-específicos.
Palabras clave: Época reproductiva, camadas, Roedores sigmodontinos, Tamaño testicular.
Abstract
In several rodent species, reproduction is regulated by habitat characteristics. From this perspective, elevation can be an important variable, since at higher elevations resources fluctuate and become more seasonal. Reproductive characteristics are expected to adjust to these conditions (i.e., seasonality), being more seasonal at higher elevation because of the energetic cost it represents. In this contribution we test this hypothesis by analyzing the reproductive patterns of two genera of sigmodontine rodents that are distributed along the eastern coordillera in Bolivia: Akodon and Oligoryzomys. The data were analyzed with a Generalized Linear Model (GLM). For both genera there were more pregnant females in the transition period (Akodon: χ2=6.26, g.l.= 1, p = 0.01; Oligoryzomys: χ2=8.10, g.l.= 1, p = 0.004). Akodon registered larger testicles in transition period at higher elevations (W = 239, p = 0.02), whereas in Oligoryzomys testicles were bigger at lower elevations (W = 256, p = 0.01). Litter size was larger in the dry season at lower elevations only in Oligoryzomys (W = 60.5, p = 0.01). Elevation and time of year affect reproductive patterns and their effect varied among the groups studied. These differences must be related to the plasticity of the species in responding to the variation in food resources availability and species-specific energetic requirements.
Key words: Reproductive season, Litter size, Sigmodontine rodents, Testicular size.
Introducción
Las estrategias reproductivas de las especies silvestres, incluidos los roedores, están influenciada por las características del hábitat, la estructura de la vegetación, los recursos alimentarios y el refugio, los cuales están determinados por factores ambientales (Morgan et al. 2000, Cameron & Scheel 2001, Rohling et al. 2005, Bumrungsri et al. 2007, Guidobono 2013). En las zonas de montaña los factores que afectan las características ambientales son elevación y época (Bumrungsri et al. 2007).
En lugares de menor elevación, donde la temperatura y la humedad son mayores, la disponibilidad de alimentos es continua durante las diferentes estaciones del año (Murphy 1992). En estos lugares se esperarían eventos reproductivos constantes en las especies de roedores, asociados a la continua disponibilidad de los recursos alimenticios (Shine & Brown 2008, Rocha et al. 2017) con camadas más grandes, por una mayor masa corporal del adulto por la cantidad y calidad de alimentos (Gentile et al. 2000, Bronson 2009, Guidobono 2013, Rübensam et al. 2015).
Por otro lado, en lugares de mayor elevación con temperaturas bajas, marcada diferencia de las estaciones y vegetación menos compleja, los recursos alimenticios serán menos diversos y su disponibilidad, cantidad y calidad, será variable por épocas (Secor 2001, Rezende et al. 2005). Por tanto, estas presiones ambientales producirán un aumento o disminución de la reproducción de las especies según la cantidad y/o calidad de recursos (del Valle & Busch 2003, Rübensam et al. 2015). En roedores, estas estrategias pueden estar basadas en plasticidad fenotípica (Pigliucci et al. 2006, Whitman & Agrawal 2009) con cambios morfológicos, fisiológicos, de desarrollo o comportamentales como una respuesta adaptativa (Sabat & Bozinovic 2000, Nespolo et al. 2002, Pigliucci 2005, Naya et al. 2008, del Valle et al. 2006, Storz et al. 2010, Cheviron et al. 2013). Las respuestas pueden incluir variación en la cantidad de crías, como camadas más grandes y un aumento de los eventos reproductivos en épocas de mayor temperatura (Trillmich et al. 2019), relacionado con una mayor producción primaria y; por tanto, un aumento en la masa corporal de los individuos (Wynne-Edwards 1998, Bears et al. 2008).
Bolivia es un país tropical con montañas de gran elevación en la parte oeste que recorren el país de norte a sur. El bosque montano en esta área tiene un gran rango altitudinal, que va desde los 1.100 hasta más de los 5.000 m, con una topografía variable y un mosaico complejo de plantas (Ribera-Aristmendi 1995) y una gran diversidad de vertebrados (Villalpando 2004, Swenson et al. 2012, Montaño-Centellas & Garitano-Zavala 2015, Vargas et al. 2018). La zona presenta dos estaciones bien marcadas a lo largo del año: seca (que coincide con el invierno) y húmeda (que coindice con el verano), así como otras dos que son las épocas de transición entre ambas, lo que produce una fluctuación en sus competentes bióticos (Ribera-Aristmendi 1995). El conocimiento de riqueza y distribución de roedores en el sitio de estudio es cada vez mayor (Aguirre et al. 2019) y aunque se reportan algunos datos de las características reproductivas de ciertas especies de roedores, no hay estudios que reporten los ciclos reproductivos, épocas de reproducción o tamaños de camada, mucho menos de manera comparativa, ya sea entre condiciones ambientales o entre especies.
Este conocimiento es necesario porque muchas especies de roedores brindan servicios ecosistémicos importantes, desde la dispersión de semillas y polinización, hasta la modificación física del ambiente al formar galerías que mejoran la calidad del suelo, pero también son un importante recurso alimenticio, al ser la base de la cadena alimentaria. Por otro lado, existen especies de roedores que pueden ser plagas y/o vectores de enfermedades (algunas especies del género Oligoryzomys), por tanto, conocer y comprender su biología reproductiva es relevante para comprender la fluctuación de sus poblaciones, y poder trabajar planes de control de plagas y alerta de zoonosis.
En ese sentido, nuestro objetivo es determinar si las estrategias reproductivas de dos géneros de roedores sigmodontinos varía según la elevación en la cordillera central de los Andes de Bolivia, en dos épocas diferentes. Los géneros estudiados son Akodon (A. aerosus, A. dayi y A. mimus) y Oligoryzomys (O. destructor destructor, O. microtis y O. occidentalis). Esperamos que las especies a menor elevación tengan varios eventos reproductivos con camadas numerosas durante todo el año, mientras que a mayores elevaciones los animales deberían aumentar los eventos reproductivos y el tamaño de sus camadas en épocas con mayor cantidad de recursos.
Área de estudio
Para el presente trabajo se evaluaron especímenes colectados en el departamento de La Paz, en los proyectos: PBE4 I (2014) y II (2017), Diversidad de roedores en Cotapata (2007-2008), Ipiri (2013), Palos Blancos (2015) y Sara Ana (2015), llevados en diferentes épocas del año, pero los especímenes que entraron en este estudio corresponden a las épocas seca y de transición seco a húmeda. Todos los especímenes se conservaron en alcohol al 96% y ninguno fue procesado antes del presente estudio.
Los animales fueron separados en dos grupos, según las características de la vegetación de las diferentes áreas (Según Navarro 2011). Los animales provenientes del sector biogeográfico amazónico de las estribaciones andinas, área que se encuentra entre 150 y 1.300 m, denominada en el presente trabajo como "menor elevación". Esta zona presenta un clima cálido casi todo el año con cortos periodos fríos, que corresponden a los meses de junio a agosto (época seca). Las estaciones lluviosas son normalmente largas produciendo alta humedad, donde las mayores precipitaciones se dan entre noviembre y febrero (época húmeda). Estas características producen bosques con un alto nivel de dosel, diferentes estratos, gran diversidad de plantas, lo que da diferentes recursos tanto alimenticios como de refugio (Navarro 2011). Un segundo grupo de animales corresponden al sector biogeográfico de Yungas de Beni, área que se encuentra por encima de 1.300 hasta 5.600 m, denominado en el presente trabajo de ahora en adelante como "mayor elevación". En este sector la temperatura disminuye y la humedad incrementa a medida que aumenta la elevación, lo que genera cambios en el tamaño del dosel y número de estratos (Navarro 2011), abarcando seis pisos ecológicos que van desde el piso nival con picos nevados (5.600 m) hasta bosques de Yungas (1.100 m) (Ribera Arismendi 1995). Para el presente trabajo, los animales analizados provienen de tres primeras zonas: Bosque de Yungas (1.200 y 2.400 m), bosque nublado (2.400-3.300 m) y Páramo yungueño (3.300-3.800 m) (Ribera Arismendi 1995). Esta región presenta una gran variación en la temperatura, siendo en la zona de trabajo mesotérmico húmedo, con lluvias de verano e inviernos secos y cálidos, pero con mejor temperatura que en verano, especialmente durante las noches (Bosque nublado y bosque de Yungas) (Ribera Arismendi 1995), generando una época seca y una húmeda.
Para el presente trabajo se evaluaron muestras provenientes de la época seca (de junio a agosto), caracterizado por las temperaturas bajas, y de transición seca a húmeda, denomina de ahora en adelante como época de transición (de agosto hasta octubre), con un incremento paulatino de la temperatura e inicio de las lluvias. No se trabajó en las otras épocas debido a que no todos los sitios contaban con muestreos en las cuatro épocas del año o había baja cantidad de especímenes colectados en las mismas.
Métodos
La comparación se realizó entre las especies de los géneros Oligoryzomys y Akodon, los dos grupos de elevación (menor y mayor) y dos épocas (transición de seca-húmeda y seca). Según los dos grupos de elevación, el número de individuos de las especies evaluadas para cada elevación fueron: A menor elevación las especies Akodon dayi (n = 57) y Oligoryzomys microtis (n = 350). A mayor elevación: Akodon aerosus (n = 32), Akodon mimus (n = 70) Oligoryzomys destructor destructor (n = 10) y Oligoryzomys occidentalis (n = 23). Para la comparación estadística las especies fueron agrupadas por géneros Akodon y Oligoryzomys.
Trabajo de laboratorio: Edad: Para separar los individuos juveniles (categoría dental de edad I and II, Myers y Patton, 1989) de los adultos (categoría dental de edad III, IV y V Myers & Pattonl989) se determinó la edad de los roedores por desgaste dentario (Myers & Patton 1989) utilizando un estereomicroscopio. De esta manera se excluyó a los individuos juveniles.
Proporción del tamaño de los testículos: Se midió el largo del testículo (en mm) (medido con un vernier LINIO modelo PIERECAL), sin tomar en cuenta el epidídimo debido a que una gran mayoría de los individuos al haber estado preservados en alcohol tenían las gónadas aplastadas. El dato obtenido se dividió por la raíz cubica del peso, para obtener la proporción del largo de los testículos según el peso de los individuos. Reproducción en las hembras: Las hembras preñadas se determinaron por la presencia o ausencia de fetos. Tamaño de la camada: se determinó por el número de fetos por hembra (Mills et al. 1992, Mallmann et al. 2011).
Análisis estadístico
Para determinar si existe alguna influencia de la época del año (seco y de transición), la elevación en la Cordillera Central de los Andes de Bolivia (menor y mayor) y el género (Akodon y Oligoryzomys), sobre las estrategias reproductivas de los individuos colectados, se utilizaron modelos lineales generalizados (GLM). Además, se realizó una comparación entre los dos grupos de elevación (alta y baja) y las épocas dentro de cada género, utilizando el análisis de chi cuadrado para el caso de las hembras y U de Mann Whitney (debido a la cantidad de datos) para el número de crías y tamaños de los testículos. Todos los análisis se los realizó en Rstudio con R versión 3.6.0 (R Core Team 2018).
La cantidad de hembras preñadas fueron analizadas siguiendo una distribución binomial. Para el tamaño de la camada, tomando en cuenta sólo hembras gestantes, utilizamos una distribución de Poisson con enlace logarítmico. Mientras que, para los machos se realizó el GLM con una distribución beta (normal). En todos los casos se incluyó la interacción entre las tres variables.
Para la distribución binomial y Poisson se usó la función "glm" para el caso de las hembras y tamaño de camada, para el caso de los machos se usó la función "betareg" del paquete "BETAREG" (Cribari-Neto y Zeileis 2010). Para la elección de los mejores modelos se usó el criterio de información Akaike con la corrección para muestras pequeñas (AICc), usando la función "dredge" del paquete "MuMin" (Barton 2020) y la clasificación por peso.
Resultados
Número de hembras preñadas : El número de hembras preñadas está influenciado por dos de las variables (Tabla 1), con una importancia de 1 para época y de 0.98 para elevación.
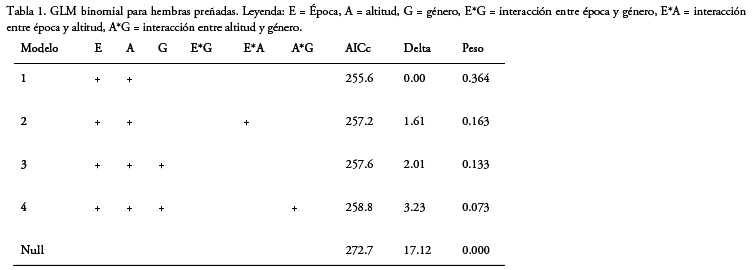
Para el género Akodon (n=173), no se registró una diferencia significativa entre elevaciones. Considerando únicamente la época, se registró una mayor cantidad de hembras preñadas en la época de transición que en la seca en ambas elevaciones, pero sólo a mayor elevación se tuvo una diferencia significativa (χ2=6.26, g.l.= 1, p = 0.01) y no así a menor elevación (Fig. 1).

Para el género Oligoryzomys (n=188), aunque más de la mitad de las hembras preñadas fueron colectadas a menor elevación, la diferencia entre mayor y menor elevación no fue significativa. A menor elevación más de la mitad de hembras preñadas se registró en época de transición (χ2=8.10, g.l.= 1, p =0.004). Mientras que, a mayor elevación, se colectaron dos hembras preñadas, por lo cual no es posible determinar diferencias entre las épocas (Fig. 1).
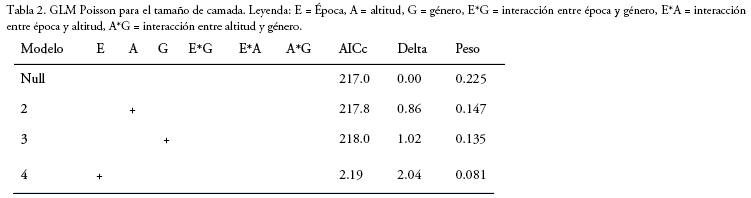
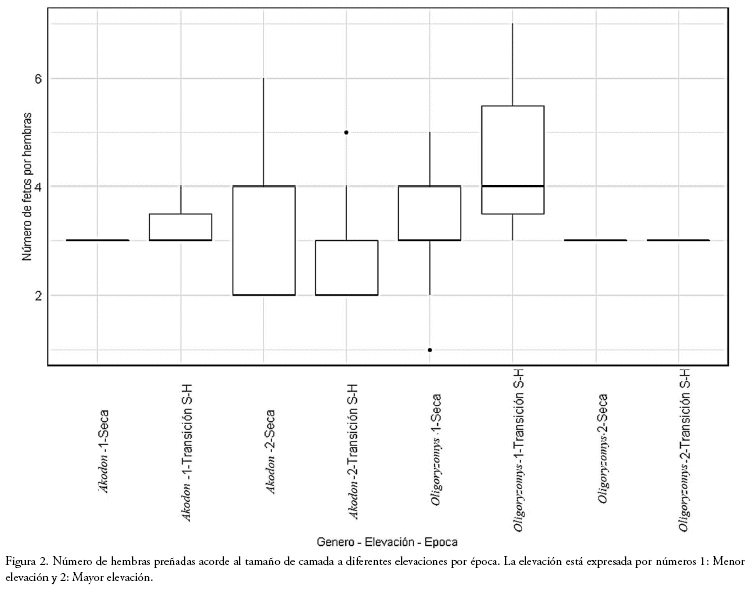
Tamaño de la camada: El GLM muestra que ninguna de las variables tiene una fuerte relación con el número de fetos por hembra. Aunque la época y elevación, tienen una importancia de 0.42 y 0.44, respectivamente, no son determinantes (Tabla 2). Sin embargo, a menor elevación se registraron camadas más grandes (W = 544, p = 0.008). Para el género Akodon (n=19) las camadas más frecuentes fueron de tres fetos a menor elevación con un incremento a cuatro en época de transición (Fig. 2). Mientras que, a mayor elevación las camadas fueron más variables, siendo la mayor de seis fetos registrado en época seca (Fig. 2). La cantidad de fetos no varió entre épocas a ninguna elevación. Para el género Oligoryzomys (n=63) a menor elevación, las hembras en época de transición presentan camadas más grandes (de hasta siete fetos) que en época seca (W=60.5, p =0.01; Fig. 2), donde tres fetos es el tamaño más frecuente (44%). No se realizó una comparación a mayor elevación debido a la baja cantidad de hembras preñadas.
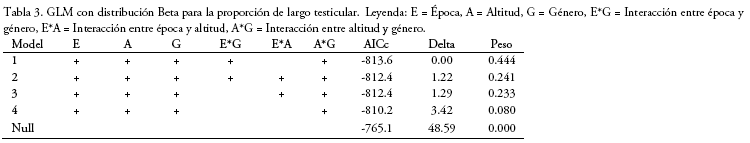
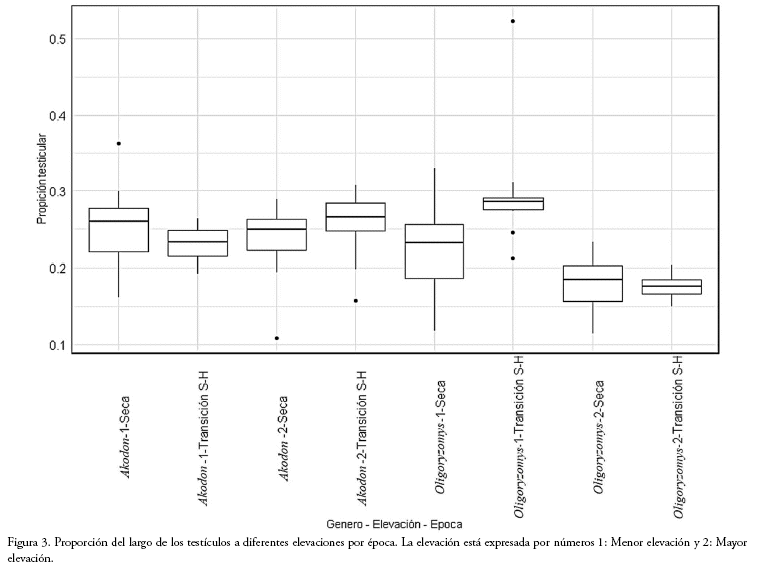
Proporción testicular de los machos: Para la proporción de los testículos, las tres variables independientes están presentes en el mejor modelo (Tabla 3) con una importancia de 1 para cada una, al igual que las interacciones entre género época (importancia de 0.69) y género elevación (1). El tamaño de los testicular no presentó diferencias entre las dos elevaciones. Para el género Akodon (n=116), el tamaño de los testículos presenta una gran variación, sin una diferencia significativa entre épocas a menor elevación, pero sí a mayor elevación (W = 239, p = 0.02; Fig. 3), presentando testículos más grandes en época de transición. Para el género Oligoryzomys (n=246) el tamaño de los testículos presenta diferencia significativa a menor elevación (W=256, p = 0.00002), presentando testículos más grandes en época de transición, y no así a mayor elevación (Fig. 3).
Discusión
En el presente estudio se pudo determinar que la elevación y la época del año afectan los patrones reproductivos en hembras y en machos, para el caso de los machos este efecto varía según el género, de acuerdo a lo obtenido en el GLM.
Esto se puede deber a que las características ambientales cambian con la elevación y la época. Por tanto, los recursos disponibles varían dando una cantidad diferente de masa y energía a los animales, los cuales deberán equilibrar las mismas para los procesos metabólicos, modificando sus estrategias reproductivas (del Valle et al. 2006, Cheviron et al. 2013). Las diferencias encontradas entre los géneros se pueden deber a las estrategias particulares de cada especie para adaptarse a sus hábitats y/o a la plasticidad fenotípica o adaptación genética (Storz et al. 2010).
Las diferencias observadas entre las elevaciones, pueden ser una respuesta a la cantidad y/o calidad del recurso alimenticio presente, siendo estos menores a mayor elevación (del Valle et al. 2006, Carnegie et al. 2011, Sassi et al. 2017). Así, a menor elevación la cantidad y la diversidad de recursos a lo largo del año es mayor y más constante, dando mejores posibilidades para tener mayor masa corporal y por tanto mayor cantidad de hembras preñadas (Akodon, Oligoryzomys) y eventos de reproducción (Akodon, Oligoryzomys) y camadas más grandes (Oligoryzomys), lo que podría verse reflejado en el crecimiento de los testículos en la época de transición de seca a húmeda (Oligoryzomys).
Estos resultados indican que la proporción de hembras preñadas varía entre épocas, en ambas elevaciones. Este se debe a la variación en temperatura, que afectará la disponibilidad de recursos alimenticios, sobre todo en época seca (Messina & Fox 2001), que se traduce en una menor cantidad de hembras preñadas en época seca, como se evidenció en el estudio (exceptuando para Oligoryzomys a mayor elevación). Si bien no se tienen datos para las otras dos épocas, se esperaría un incremento de la reproducción en la época húmeda.
Para el tamaño de camada por hembra, el GLM no identificó una variable de importancia entre elevación, época o género, lo que se puede deber a que esta variable depende de rasgos particulares de cada especie (p.e. masa corporal). Aunque sí se observaron camadas más grandes en la época de transición, particularmente a menor elevación para cada género, que es lo que se esperaba gracias a una mayor cantidad de nutrientes a menor elevación (Secor 2001, Rezende et al. 2005). Es posible que, al incrementar el tamaño de la muestra y en el esfuerzo de captura por épocas, estas diferencias se hagan más evidentes.
El tamaño de los testículos presenta una variación dependiente al género por elevación (Fig. 3), mostrando un aumento del tamaño para Akodon (mayor elevación época de transición) y Oligoryzomys (menor elevación época de transición) que puede estar relacionado al incremento de hembras receptivas. La variación en el tamaño testicular entre los géneros y entre las elevaciones, puede deberse a que las características del hábitat (menores temperaturas o menor cantidad de oxígeno a mayor elevación) afectan su fisiología y por tanto, la cantidad de espermas (Reyes et al. 2012).
Analizando los resultados por género, para Akodon en ambas elevaciones se incrementa la proporción de hembras preñadas en época de transición, lo que sería una respuesta al aumento de recursos en esta época (González-Chávez et al. 2017). Además, a mayor elevación, los machos presentaron un incremento en el tamaño de sus testículos, lo que puede ser resultado de la presencia de hembras receptivas o un reflejo del tipo de sistemas de reproducción de la especie. Un mayor tamaño testicular, puede darse por una alta competencia intrasexual (Liao & Lu. 2011), aumentando así la cantidad de esperma y las posibilidades de fecundar a más hembras. No se conoce la estrategia reproductiva de las especies de Akodon estudiadas en este trabajo y es algo aún pendiente de determinar.
La variación en el tamaño de la camada para Akodon, podría responder a las características del hábitat y las características anatómicas y morfológicas de cada especie (Fig. 2) esto explicaría la mayor variación, de dos a seis crías, en el tamaño de las camadas en lugares a mayor elevación. A menor elevación, la variación en las épocas que muestra Akodon dayi (tres fetos en época seca y de tres a cuatro fetos en época de transición) puede ser explicada por la disponibilidad y calidad de alimento, dándose un aumento de energía dando la posibilidad de tener camadas más grandes en época de transición. A mayor elevación, el tamaño de la camada fue más variable para A. aerosus (n = 2-6) que para A. mimus (n = 2). Esta variación puede ser explicada por las condiciones ambientales dónde se encuentra cada especie: A. aerosus se encuentra entre los 1.300 a 2.400 msnm, donde las condiciones climáticas y la disponibilidad de alimento pueden ser menos adversas que para A. mimus, que habita por encima de los 3.000 msnm, donde la baja cantidad de fetos puede ser el resultado de la presión ambiental adversa (temperatura, cantidad de oxígeno, entre otras) a esta elevación. Sin embargo, se espera que en la época húmeda haya mejores condiciones ambientales y el tamaño de la camada aumente, al igual que el número de hembras gestantes. Esto es algo que requiere mayor trabajo de campo, en las diversas épocas.
Para el género Oligoryzomys, la cantidad de hembras preñadas y el tamaño de testículos varían con la época y la altitud, mostrando que las características ambientales son importantes para comprender su reproducción, esto está más claro a menor elevación que a mayor elevación. Oligoryzomys microtis (a menor elevación) tiene un mayor número de hembras preñadas en época de transición (83.7%) que en época seca (16.2%). No se esperaba una diferencia tan grande en la cantidad de hembras preñadas a baja elevación, ya que las condiciones climáticas al ser más constantes a lo largo del año, no afectarían la producción primaria por ende el efecto sería mínimo sobre la reproducción (Shine & Brown 2008, Rocha et al. 2017). Estos resultados indicarían que para la reproducción de esta especie en particular la calidad de recursos alimenticios es importante, no sólo la cantidad. El aumento de recursos, posiblemente con más nutrientes, coincide con la mayor proporción de hembras con camadas más grandes en la época de transición, respaldando la importancia de la cantidad de nutrientes para el embarazo (González-Chávez et al. 2017). Este incremento tanto de hembras preñadas, como del tamaño de camada puede ser por la fluctuación de los cultivos en la zona, ya que O. microtis es una especie asociada a la agricultura (llegando a ser considerada plaga), mostrando la plasticidad de la especie a condiciones ambientales para poder reproducirse. El incremento de hembras preñadas se relaciona con un aumento testicular de los machos, determinada también por una competencia intrasexual en la época (Liao & Lu 2011), generando mayor cantidad de esperma para aumentar las posibilidades de fecundación (Zebunuto et al. 2002).
Para las especies a mayor elevación (O. destructor destructor y O. occidentalis) sólo se registraron una hembra preñada para cada especie en diferentes épocas (de 15 hembras colectadas), además los testículos no presentaron una diferencia entre épocas. Ambos resultados podrían deberse a que aún no tienen los recursos necesarios para que pueda iniciarse la reproducción (Reyes et al. 2012).
Las diferencias observadas entre los géneros Akodon y Oligoryzomys, se pueden deber a la diferencia en hábitos alimenticios, calidad de nutrientes y a que la energía puede ser destinada a otras actividades de sobrevivencia (González-Chávez et al. 2017). Pero también puede ser una respuesta adaptativa por parte de las especies de Oligoryzomys que se encuentran en áreas con mejor oferta de recursos alimenticios (Secor 2001, Rezende et al. 2005). Las diferencias también pueden deberse a la masa corporal de los individuos (Happold 2011) y, por tanto, a su dieta y requerimientos de nutrientes (González-Chávez et al. 2017, Douglas de Carvalho et al. 2019), los cuales están ligados al clima, que afecta las características del hábitat (Messina & Fox 2001) y la plasticidad de los mamíferos (Hansen & Boonstra 2000), todas estas variables importantes para la reproducción.
Para comprender mejor la reproducción en las hembras, es importante incorporar factores como las características morfológicas y fisiológicas de los individuos, como peso, longitud corporal y dieta (cantidad y calidad), esta última podría explicar la cantidad de fetos por hembra (González-Chávez et al. 2017). Además, recomendamos considerar otros datos de las hembras, que deben registrarse en fresco, como la presencia del tapón vaginal, que se da en el momento que la hembra está preñada, la cantidad de heridas en los cuernos uterinos, que es la única forma para determinar si una hembra no preñada es activa reproductivamente. Mientras que, para los machos es importante tomar las medidas de los testículos en fresco (largo y ancho). No se recomienda trabajar con la información de la posición de los testículos (escrotado o abdominal) ya que estudios realizados por Boiani y colaboradores (2008) indican que debido al estrés producido al animal por la manipulación puede producir un descenso de los testículos, dando información errónea de los periodos de reproducción de las especies. Además, para estudios de reproducción en machos también es importante tomar una muestra del epidídimo para estudios de espermas. El material espermático de animales en alcohol debe ser comparado con material fresco para determinar si el material en alcohol da datos confiables.
Por último, sugerimos que se aumente la cantidad de puntos de muestreo, tanto en elevaciones estudiadas, como épocas del año, para corroborar las conclusiones generales de nuestro trabajo. Además, un muestreo más completo y que incluya el ciclo reproductivo completo de las especies de interés, permitirá una visión integral del efecto de la elevación en dichos patrones.
Conclusiones
Si bien varios puntos de la discusión son especulativos debido a la falta de información en este tema, podemos concluir que el cambio de recursos a través de las estaciones es relevante para los dos géneros estudiados, mostrando una mayor reproducción en época de transición seca a húmeda que en época seca (exceptuando Oligoryzomys a mayor elevación). Mientras que la altitud es una variable relevante, sobre todo en machos de Oligoryzomys, presentando testículos de mayor tamaño a bajas elevaciones, y en hembras de Akodon, donde la cantidad de hembras gestantes a menor elevación para ambas épocas.
Finalmente, es importante llevar a cabo más estudios de la reproducción de los roedores silvestres tomando datos con mayor detalle del ambiente y de los individuos en el momento de la colecta. De esta manera se comprenderá mejor las fluctuaciones espaciales y temporales de las poblaciones de roedores. Esta información es determinante para realizar monitoreo y control de las poblaciones de roedores en sitios y épocas determinados, que puedan ser consideradas un problema para la salud humana o la agricultura.
Agradecimientos
Este trabajo se basa en especímenes colectados en los proyectos: Diversidad de roedores en Cotapata (Fondos 14-36098G y LO1415 del MSMT - Ministerstvo Skolství, Mládeze a Télovychovy, Global Change Research Institute, República Checa), Ipiri (Beca 74031 de la Fundación John D. & Catherine T. MacArthur al Instituto de Ecología), PBE4 (fondo ASDI a la UMSA), PBE4 II, Palos Blancos y Sara Ana (fondos del Impuesto Directo a los Hidrocarburos: IDH 2013-2014, IDH 2013-2014, IDH 2015-2016, respectivamente). Agradecemos a la Colección Bolivia de Fauna (Instituto de Ecología - UMSA y Museo Nacional de Historia Natural) por la disponibilidad de espacio de trabajo, a los estudiantes y colaboradores que trabajaron en la obtención de los especímenes y a los revisores anónimos que brindaron comentarios para esta versión final.
Referencias
Aguirre L.F., T. Tarifa, R. B. Wallace, N. Bernal H.5, L. Siles, E. Aliaga-Rossel & J. Salazar-Bravo. 2019. Lista actualizada y comentada de los mamíferos de Bolivia. Ecología en Bolivia 54(2): 107-147.
Barton K. 2020. MuMIn: Multi-Model Inference. R package version 1.43.17. https://CRAN.Rproject.org/package=MuMIn [ Links ]
Bears, H., K. Martin & G.C. White. 2008. Breeding in high-elevation habitat results in shift to slower lifehistory strategy within a single species. Journal of Animal Ecology 78: 365-375.
Boiani, L., N. Berois & G. D’Elía. 2008. Annual male reproduction cycle of a hantavirus reservoir, the long-tailed mouse Oligoryzomys flavescens (Rodentia; Cricetidae, Sigmodontinae) from Uruguay. Mastozoología Neotropical 15(1): 23-32.
Bumrungsri, S., W. Bumrungsri & P.A. Racey. 2007. Reproduction in the short-nosed fruit bat in relation to environmental factors. Journal of Zoology 272:73-81.
Bronson, F. 2009. Climate change & seasonal reproduction in mammals. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences 364: 3331-3340. [ Links ]
Cameron, G. & D. Scheel. 2001. Getting Warmer: Effect of global climate change on distribution of rodents in Texas. Journal of Mammalogy 82: 652-680.
Carnegie, S.D., L.M. Fedigan & A.D. Melin. 2011. Reproductive seasonality in female capuchins (Cebus capucinus) in Santa Rosa (Área de Conservación Guanacaste), Costa Rica. International Journal of Primatology 32: 1076-1090.
Cheviron, Z.A., G.C. Bachman & J.F. Storz. 2013. Contributions of phenotypic plasticity to differences in thermogenic performance between highland and lowland deer mice. Journal of Experimental Biology 216: 1160-1166.
Cribari-Neto, F. & A. Zeileis. 2010. Beta regression in R. Journal of Statistical Software 34(2): 1-24. URL http://www.jstatsoft.org/v34/i02/.
del Valle, J.C. & C. Busch. 2003. Body composition and gut length of Akodon azarae (Muridae: Sigmodontinae): relationship with energetic requirements. Acta Theriologica 48(3): 347-357.
del Valle, J.C., C. Busch & A.A. López-Mañanes. 2006. Phenotypic plasticity in response to low quality diet in the South American omnivorous rodent Akodon azarae (Rodentia: Sigmodontinae). Comparative Biochemistry and Physiology, Part A 145: 397-405.
Gentile, R., P.S. D`Andrea, R. Cerqueira & L.S. Maroja. 2000. Population dynamics and reproduction of marsupials and rodents in a Brazilian rural area: a five-year study. Studies on Neotropical Fauna and Environment 35: 1-9.
González-Chávez, B., V. Rojas-Díaz & L. Cruz-Bernate. 2017. Demographic Parameters of the Silky Shrew-Opossum Caenolestes fuliginosus (Paucituberculata, Caenolestidae) along an Altitudinal Gradient in the Cordillera Central of the Colombian Andes. Journal of Mammal Evolution doi:10.1007/s10914-017-9417-6.
Guidobono, J.S. 2013. Dinámica poblacional de roedores en agroecosistemas y su relación con variables ambientales. Tesis de doctorado, Universidad de Buenos Aires, Buenos Aires. [ Links ]
Hansen, T.F. & R. Boonstra. 2000. The best in all possible worlds? A quantitative genetic study of geographic variation in the meadow vole, Microtus pennsylvanicus. Oikos 89: 81-94.
Happold, D.C.D. 2011. Reproduction and ontogeny of Mastacomys fuscus (Rodentia: Muridae) in the Australian Alps and comparisons with other small mammals living in alpine communities. Mammalian Biology 76: 540-548. [ Links ]
Liao, W.B. & X. Lu. 2011. Variation in body size, age and growth in a subtropical treefrog (Rhacophorus omeimontis) along an altitudinal gradient in western China. Ethology Ecology and Evolution 23: 248-261.
Mallmann, A.S., M. Finokiet, A.C. Dalmoso, G.L. Melo, V.L. Ferreira & N.C. Caceres. 2011. Dinâmica populacional e reprodução de pequenos mamíferos de uma Floresta Estacional do Maciço do Urucum, oeste do Pantanal, Brasil. Neotropical Biology and Conservation 6(2): 94-102.
Messina, F.J. & C.W. Fox. 2001. Offspring size and number. pp. 113-127. En: Fox, C.W., D.A. Roff & D.J. Fairbairn (eds.) Evolutionary Ecology: Concepts and Case Studies. Oxford University Press, Nueva York.
Mills, J.N., B.A. Ellis, K.T. McKee, J.I. Maiztegui & J.E. Childs. 1992. Reproductive characteristics of rodent assemblages in cultivated regions of central Argentina. Journal of Mammalogy 73(3): 515-526.
Montaño-Centellas, F.A. & A. Garitano-Zavala. 2015. Andean bird responses to human disturbances along an elevational gradient. Acta Oecologica 65-66: 51-60.
Morgan, E., J.H. Brown & R.R. Parmenter. 2000. Rodents, plants, and precipitation: spatial and temporal dynamics of consumers and resources. Oikos 88: 470-482.
Murphy, E.C. 1992. The effects of natural increase in food supply on a wild population of house mice. New Zealand Journal of Ecology 16(1): 33-40. [ Links ]
Myers, P. & L. Patton. 1989. A new species of Akodon from the cloud forests of eastern Cochabamba Department, Bolivia (Rodentia: Sigmodontinae). Occasional papers of the Museum of Zoology (University of Michigan) 72(1): 1-28.
Navarro, G. 2011. Clasificación de la vegetación de Bolivia. Centro Simón I. Patiño, Santa Cruz. [ Links ]
Naya, D.E., F. Bozinovic & W.H. Karasov. 2008. Latitudinal trends in digestive flexibility: testing the climatic variability hypothesis with data on the intestinal length of rodents. The American Naturalist 172(4): 122-134.
Nespolo, R.F., L.D. Bacigalupe, P. Sabat & F. Bozinovic. 2002. Interplay among energy metabolism, organ mass and digestive enzyme activity in the mouse–opossum Thylamys elegans: the role of thermal acclimation. Journal of Experimental Biology 205: 2697-2703.
Pigliucci, M., C. J. Murren & C. D. Schlichting. 2006. Phenotypic plasticity and evolution by genetic assimilation. The Journal of Experimental Biology 209: 2362-2367.
Rezende, E.L., F.R. Gomes, C.K. Ghalambor, G.A. Russell & M.A. Chappell. 2005. An evolutionary frame of work to study physiological adaptation to high altitudes. Revista Chilena de Historia Natural 78: 323-336.
Reyes, J.G., J.G. Farias, S. Henríquez-Olavarrieta, E. Madrid, M. Parraga, A.B Zepeda & R.D. Moreno. 2012. The hypoxic testicle: physiology and pathophysiology. Oxidative Medicine and Cellular Longevity Doi:10.1155/2012/929285
Ribera Arismendi, M.O. 1995. Caminos de Cotapata. Instituto de Ecología, La Paz. [ Links ]
Rocha, C.R., R. Ribeiro & J. Marinho-Filho. 2017. Influence of temporal variation and seasonality on population dynamics of three sympatric rodents. Mammalian Biology 84: 20-29.
Rohling, I., V.M. Guedes, A.P. Lima & W.E. Magnusson. 2005. Spatially explicit population dynamics in a declining population of the tropical rodent, Bolomys lasiurus. Journal of Mammalogy 86(4): 677-682.
R Core Team. 2018. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Viena. URL https://www.Rproject.org/ [ Links ]
Rübensam, K., R. Hribal, K. Jewgenowa & A. Guenther. 2015. Seasonally different reproductive investment in a medium-sized rodent (Cavia aperea). Theriogenology 84: 639-644.
Sabat, P. & F. Bozinovic. 2000. Digestive plasticity and the cost of acclimation to dietary chemistry in the omnivorous leaf-eared mouse Phyllotis darwini. Journal of Comparative Physiology B 170: 411-417.
Sassi, P.L., J. Menéndez & M.F. Cuevas. 2017. Geographic variation in life-history traits: factors shaping energy investment in reproduction in a highland dwelling rodent. Journal of Zoology doi:10,1111/jzo,12523
Secor, S.M. 2001. Regulation of digestive performance: a proposed adaptative response. Comparative Biochemical Physiology Part A 128: 565-577. [ Links ]
Shine, R. & G.P. Brown. 2008. Adapting to the unpredictable: reproductive biology of vertebrates in the Australian wet–dry tropics. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences 363: 363-373.
Storz, J.F., G.R. Scott & Z.A. Cheviron. 2010. Phenotypic plasticity and genetic adaptation to highaltitude hypoxia in vertebrates. Journal of Experimental Biology 213: 4125-4136.
Swenson, J.J., B.E. Young, S. Beck, P. Comer, J.H. Córdova. et al. 2012. Plant and animal endemism in the eastern Andean slope: challenges to conservation. BMC Ecology 12: 1-18.
Trillmich, F., E. Geißler & A. Guenther. 2019. Senescence and costs of reproduction in the life history of a small precocial species. Ecology and Evolution 9: 7069-7079.
Vargas, J., P. Flores & J. Martínez. 2018. Pequeños mamíferos en dos áreas protegidas de la vertiente oriental boliviana, considerando la variación altitudinal y la formación vegetacional. Boletín REDESMA. http://revistavirtual.redesma.org/vol2/pdf/ambiental/mamiferos.pdf
Villalpando, G. 2004. Variación altitudinal y temporal de roedores y marsupiales en el área del cerro Hornuni del Parque Nacional y Área de Manejo Integrado Cotapata. Tesis de licenciatura en biología, Universidad Mayor de San Andrés, La Paz. [ Links ]
Whitman, D.W. & A.A. Agrawal. 2009. What is phenotypic plasticity and why is it important? Pp. 1-63. En: Whitman, D.W. & T.N. Ananthakrishnan (eds.) Phenotypic Plasticity of Insects: Mechanisms and Consequences. Science Publishers, Boca Raton https://doi.org/10.1201/9780367803568
Wynne-Edwards, K.E. 1998. Evolution of parental care in Phodopus: conflict between adaptations for survival and adaptations for rapid reproduction. American Zoologist 38: 238-250. [ Links ]
Zebunuto, R.R., A.I. Vassallo & C. Busch. 2002. Social and reproductive behavior of the subterranean solitary rodent Ctenomys talarum (Rodentia: Ctenomyidae) in a seminatural enclosure. Revista Chilena de Historia Natural 75(1): 165-177.

