










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkEcología en Bolivia
Print version ISSN 1605-2528On-line version ISSN 2075-5023
Ecología en Bolivia vol.56 no.1 La Paz Apr. 2021
Nota
Ecomorphological variations of Orestias sp. (gr agassizii, Cyprinodontiformes, Cyprinodontidae) from Eastern slope of the Andes
Variaciones ecomorfológicas de Orestias sp. (gr agassizii, Cyprinodontiformes, Cyprinodontidae) de la Vertiente Oriental de los Andes
Erick Loayza1*, Débora Alvestegui1, Kelvin Herbas1, Carla Ibañez1, Claudia Zepita1 & Guido Miranda-Chumacero1,2
1Unidad de Ecología Acuática, Instituto de Ecología, Facultad de Ciencias Puras y Naturales, Universidad Mayor de San Andrés, c/27, Cota Cota s/n Campus Universitario, La Paz, Bolivia
2Wildlife Conservation Society Bolivia (WCS), Calle Gabino Villanueva#340, entre las calles 24 y 25 de Calacoto, La Paz, Bolivia
*Autor de correspondencia: erickz.loayzatorrico@gmail.com
Recibido: .16.06.20, Aceptado: 12.12.20.
Andes mountains in Bolivia are a water divider that gives rise to the Amazon basin in the Eastern and the Altiplano basin in the Western slope. Orestias (Cuvier & Valenciennes 1846) is one of three native genera of endemic teleost fish of the Altiplano (Parenti 1984, Villwock 1986, Vila et al. 2007) and is distributed from southern Perú, western Bolivia and northeast Chile, with Lake Titicaca as its main distribution area (Parenti 1984, Guerlesquin 1992, Lauzanne 1992). More than 40 species were described in the Altiplano (Parenti 1984, Vila et al. 2007, Sarmiento et al. 2014), although there is some divergence on species validity based on genetic traits (Lüssen et al. 2003, Vila et al. 2007, Garrigos et al. 2013, Ibañez et al. 2014, Takahashi & Moreno 2015). The distribution of this native genus has been a consequence of geological events that occurred during the Holocene and late Pleistocene, which have developed small disconnected basins along the Altiplano in the Western slope, where Orestias maintains small populations that have remained isolated in an arid and extreme climate (Parker & Kornfield 1995, Vila et al. 2010, 2013). The wide variety of ecological conditions in the distribution area has led to morphological differences with local adaptations and a particular phenotypic expression (Lauzanne 1982, Maldonado et al. 2009). For instance, Orestias agassizii, the species with the largest distribution of the genus, inhabits different aquatic ecosystems from the north to the south of the Altiplano, and shows high intra-specific variation (Arratia 1982, De la Barra et al. 2009, Flores 2013, Takahashi & Moreno 2015).
Although Orestias species has been described only for Western slope, there are small little-known and well stablished populations of these fish in the headwaters from Eastern slope (Sarmiento & Barrera 1997, Identidad Madidi & SERNAP 2017), characterized by an abrupt relieve with pronounced slopes, deep and enclosed alluvial valleys, that provide new geomorphologic and ecological conditions to which this native fish could adapt (e.g. increased turbidity and water flow). Orestias populations of agassizii group reported for the Puina Valley, at the headwaters of the Tuichi River on the Eastern slope were introduced about 30 to 40 years ago. They are nowadays well established and correspond to the same group, although their identity is still unknown (Identidad Madidi & SERNAP 2017). The aim of this study was to analyse and compare the ecomorphological variation among the Orestias sp. (gr agassizii) populations from the Puina Valley on the Eastern slope and from Western slopes of the Andes mountains.
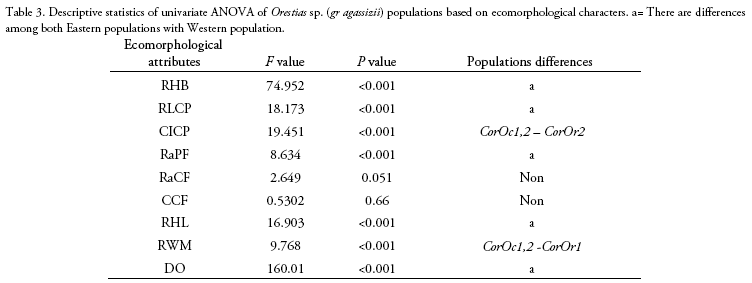
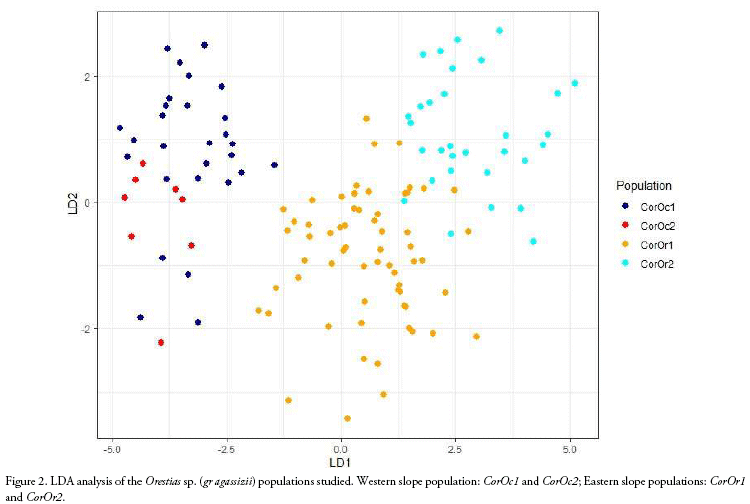
We compared ecomorphological measurements of 130 specimens corresponding to four Orestias sp. (gr agassizii) populations. Western slope populations were collected from Hichu Kotha (CorOc1: 28 specimens, -68.380019°; -16.174192°; 4,350 m), and Kara Kotha (CorOc2: 8 specimens, -68. 376306°; -16.150344°; 4,450 m) lagoons from Hichu Khota Valley, northwest of La Paz city, in the "glacial cordillera" hydro-ecoregion (Wasson et al. 2002). Eastern slope populations were collected from Celeste (CorOr1: 63 specimens, -69.163199°; -14.650412°; 4,200 m), and "Siembra" (CorOr2: 31 specimens, -69.089377°; -14.617934°; 4,300 m, Figure 1) lagoons in Puina Valley. Nine ecomorphological attributes were evaluated (Table 1). Measurements were made to the nearest 0.1 mm using digital calliper (absolute digimatic digital callipers, Mitutoyo, Japan). A one-way MAN OVA analysis (Willks lambda test) was performed to determine the significant differences on ecomorphological attributes among populations (p < 0.05). When MANO VA was significant, corresponding one-way ANOVAs followed of Dunnett's test were subsequently performed for each ecomorphological character to evalúate its potential contribution to differences among the populations. A Linear Discriminant Analysis (LDA) was carried out using R (package MASS; Ripley et al. 2020) to determine how accurately the morphology of specimens corresponded to their respective populations.
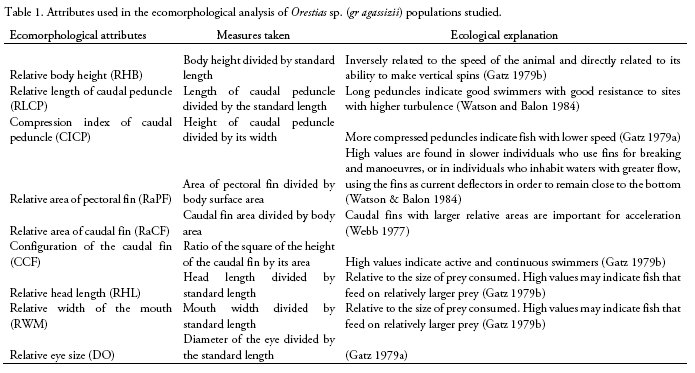
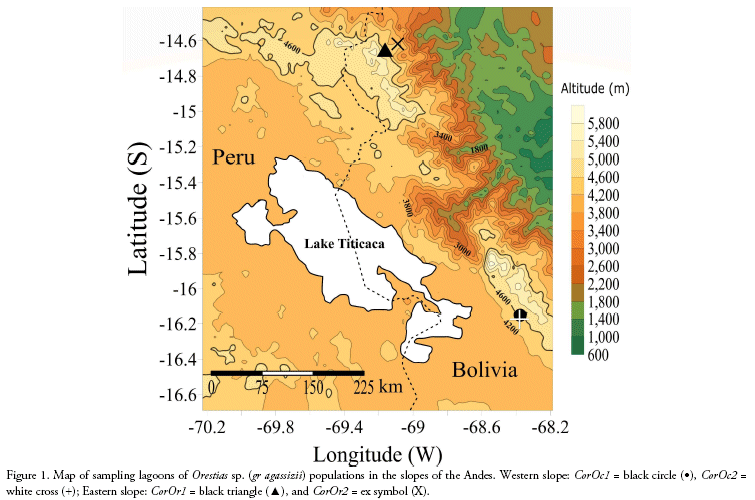
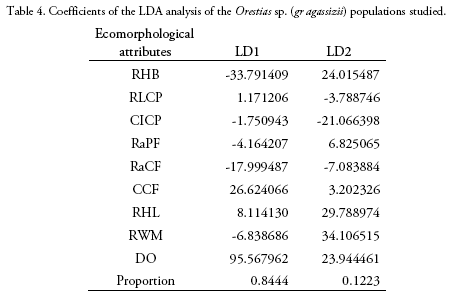
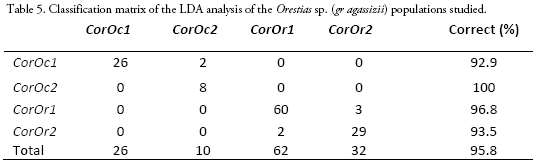
Length and weight data including ranges and means are shown in Table 2. The MANOVA revealed that studied populations showed globally significant ecomorphological differences (F=31.4, Wilk's λ = 0.08, p < 0.001). Most ecomorphological attributes differentiated the populations from each mountain slope, with the exception of the caudal fin configuration (CCF, Table 3). Indeed, LDA analysis showed that the three populations were correctly discriminated according to their ecomorphological attributes (>95% of correct classification, Fig. 2, Table 4 and 5). The linear discriminant coefficients revealed that the first function (explaining 84.44% of the variation) differentiated populations principally according to eye size (DO) and relative body height (RHB). The second function (12.23% of the variation) differentiated populations according to relative width of the mouth (RWM) and the compression index of caudal peduncle (CICP).
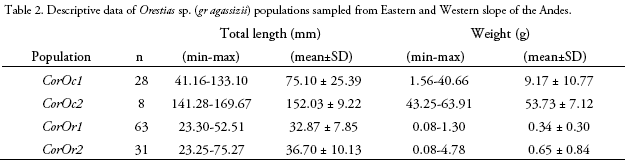
Living conditions in high altitude lagoons (> 3,500 m) are extreme, with high exposure to solar radiation, which can even be up to 20% higher than that recorded at sea level at the same latitude (Aceituno 1997). Moreover, the valleys located on both sides of the Andes slopes are exposed to intense climate warming caused by the combined effects of tropical location (16°S) and altitude, projecting twice the planet average warming towards 2100 (Bradley 2006, Valdivia et al. 2013, Rangecroft et al 2016).
The formation of diese lagoons on the western slope corresponds to the evaporation of large paleo-lakes (Keller & Soto 1998, Placzek et al. 2006), unlike the lagoons on the Eastern slope, which have a glacial origin. The retreat of the waters of paleo-lakes caused a fragmentation, leaving small lagoons scattered in the Altiplano, many of them without any connection between them.
Although, the Hichu Khota and Puina Valleys are located in the "glacial cordillera" hydro-ecoregion on the humid mountains region (Wasson et al. 2002), they have some differences, caused by the geography of each slope. The waters that flow into the Hichu Khota Valley come from melting glaciers that run through a series of low-slope moraine hills (7-10%), that arrives to a fiuvio-glacial foothill, to finally cross the Altiplano and end up in the Lake Titicaca (Marín 1989). In contrast, the waters that run through the Puina Valley, on the Eastern slope, flow on steep slopes (> 20%), descending rapidly on sedimentary rock and presenting high turbidity values caused by the natural instability of the substrate (Identidad Madidi & SERNAP 2017), finally flowing into the Tuichi River in the ANMI-Madidi in the Amazon basin.
Indeed, morphological variations can be influenced by the habitat, the search for food resource or environmental stress conditions (Webb 1984, Parsons & Robinson 2007). Environmental differences between both slopes of the Andes could influence the body shape of the Orestias populations studied, as for example the length of the peduncle (RLCP), adapting to a greater water flow in the Eastern slope. Likewise, this change in water flow could also influence the macroinvertebrates composition, the main food resource of Orestias, which could explain the differences found related to the head shape (head size, mouth width and eye diameter).
It is evident that the presence of Orestias in the Eastern slope was not entirely natural (Identidad Madidi & SERNAP 2017), despite the presence of aquatic environments with similar characteristics to those inhabited by Orestias (De la Barra et al. 2009). Conversely, the wide distribution of Orestias in the Altiplano is attributed to a very extensive hypothetical ancestral distribution (Lüssen et al. 2003), which accompanied the evaporation processes of the paleo-lakes. This fragmentation caused changes to accumulate separately, and evolve into different species that diversified through allopathic speciation (Northcote 2000). This process was observed in Orestias from north of Chile (Vila et al. 2013), even going so far as to describe a new genre (Arratia et al. 2017).
The presence of Orestias in the Eastern slope of the Andes is not well documented (Sarmiento & Barrera 1997). Furthermore, many aspects of the hydrology (e.g. water quality, flow characteristics) of the headwaters from Amazon basin remain unknown. In conclusion, significant differences between Orestias populations from each Andean slope were found, based on peduncle length, head size, width mouth and eye diameter. These results highlighted the need for further studies in order to identify these "Eastern Orestias", applying genetic analysis or DNA barcodes, which would allow us to confirm or exclude the hypothesis that these Eastern populations would be geographical variations of a single species.
Acknowledgments
The authors would like to acknowledge to the Research Institute for Development (IRD), for allowing us to use the samples from FISHLOSS project. Also would like to acknowledge to the Wildlife Conservation Society (WCS-Bolivia) and the Puina field guides for their cooperation and logistical support in the framework of the Identidad Madidi expedition.
References
Aceituno, P. 1997. Aspectos generales del clima en el Altiplano Sudamericano. pp 63–69. En: Charrier, R (ed.) El Altiplano, Ciencia y Conciencia de los Andes. Santiago de Chile.
Arratia, G. 1982. Peces del Altiplano de Chile. El hombre y los ecosistemas 1:93–133.
Arratia, G., I. Vila, N. Lam, C.J. Guerrero & C. Quezada-Romegialli. 2017. Morphological and taxonomic descriptions of a new genus and species of killifishes (Teleostei: Cyprinodontiformes) from the high Andes of northern Chile. PLOS ONE 12: e0181989.
Bradley, R.S. 2006. Climate Change: Threats to Water Supplies in the Tropical Andes. Science 312:1755–1756.
Cuvier, G.L. & A. Valenciennes. 1846. Des Orestias. pp 221–244. En: Cuvier, G. L., & A. Valenciennes (eds.). Histoire naturelle des poissons. Paris.
De la Barra, E., E. Maldonado, F. Carvajal & J. Coronel. 2009. The killifish Orestias in the Bolivian Puna. Rev. Bol. Ecol. Y Cons. Amb. 26:1-7.
Flores, A. 2013. Ecomorfología y ecología alimentaria del género Orestias (Pisces Cyprinodontiformes) en la puna xerofítica de la provincia de Sud Lípez, Potosí Bolivia. Tesis de licenciatura en biología, Universidad Mayor de San Andrés, La Paz. 80 p. [ Links ]
Garrigos, Y.E., B. Hugueny, K. Koerner, C. Ibañez, C. Bonillo, P. Pruvost, R. Causse, C. Cruaud & P. Gaubert. 2013. Non-invasive ancient DNA protocol for fluid-preserved specimens and phylogenetic systematics of the genus Orestias (Teleostei: Cyprinodontidae). Zootaxa 3640:373–394.
Gatz, A. 1979a. Ecological morphology of freshwater stream, fishes. Tulane studies in zoology and botany 21:91–119.
Gatz, A.J. 1979b. Community Organization in Fishes as Indicated by Morphological Features. Ecology 60:711–718.
Guerlesquin, M. 1992. Charophythes. pp 232–240. En: Dejoux, C. & A. Iltis (eds.) Lake Titicaca: A synthesis of limnological knowledge. Springer Netherlands, Dordrecht.
Ibañez, C., B. Hugueny, Y. Esquer-Garrigos, C. Zepita & R. Gutierrez. 2014. Biodiversidad íctica en el Lago Titicaca. pp 134–153. En: Pouilly, M., X., Lazzaro, D. Point & M. Aguirre (eds.) Línea base de conocimientos sobre los recursos hidrológicos en el Sistema TDPS con enfoque en la cuenca del Lago Titicaca. IRD-UICN, Quito.
Identidad Madidi & SERNAP (Servicio Nacional de Áreas Protegidas). 2017. Informe Científico 2015. Relevamientos de biodiversidad en el Parque Nacional y Área Natural de Manejo Integrado Madidi. La Paz. 184 p. [ Links ]
Keller, B. & D. Soto. 1998. Hydrogeologic influences on the preservation of Orestias ascotanensis (Teleostei: Cyprinodontidae), in Salar de Ascotán, northern Chile. Revista Chilena de Historia Natural 71:147–156.
Lauzanne, L. 1982. Les Orestias (Pisces, Cyprinodontidae) du Petit Lac Titicaca. Revue d’Hydrobiologie Tropicale. 15:39–70.
Lauzanne, L. 1992. Native species the Orestias. pp 405–419. En: Dejoux, C. & A. Iltis (eds) Lake Titicaca: A synthesis of limnological knowledge. Springer Netherlands, Dordrecht.
López, R.P. 2000. La prepuna boliviana. Ecología en Bolivia 34:45–65.
Lüssen, A., T. M. Falk & W. Villwock. 2003. Phylogenetic patterns in populations of Chilean species of the genus Orestias (Teleostei: Cyprinodontidae): results of mitochondrial DNA analysis. Molecular Phylogenetics and Evolution 29:151–160.
Maldonado, E., N. Hubert, P. Sagnes & B. De Merona. 2009. Morphology-diet relationships in four killifishes (Teleostei, Cyprinodontidae, Orestias) from Lake Titicaca. Journal of Fish Biology 74:502–520.
Marín, R. 1989. Elementos para la tipología de los ríos de altura de la región de La Paz: Características biológicas y potencialidades piscícolas. Tesis de licenciatura en biología, Universidad Mayor de San Andrés, La Paz. [ Links ]
Northcote, T.G. 2000. Ecological interactions among an Orestiid (Pisces: Cyprinodontidae) species flock in the littoral zone of Lake Titicaca. Advances in Ecological Research 31:399–420.
Parenti, L.R. 1984. A taxonomic revision of the Andean killifish genus Orestias (Cyprinodontiformes, Cyprinodontidae). American museum of natural history. Bulletin of the American Museum of Natural History 178(2): 1-214.
Parker, A. & I. Kornfield. 1995. Molecular Perspective on Evolution and Zoogeography of Cyprinodontid Killifishes (Teleostei; Atherinomorpha). Copeia 1995(1): 8-21.
Parsons, K.J. & B.W. Robinson. 2007. Foraging performance of diet‐induced morphotypes in pumpkinseed sunfish (Lepomis gibbosus) favours resource polymorphism. Journal of evolutionary biology 20:673–684.
Placzek, C., J. Quade & P.J. Patchett. 2006. Geochronology and stratigraphy of late Pleistocene lake cycles on the southern Bolivian Altiplano: Implications for causes of tropical climate change. Geological Society of America Bulletin 118: 515–532.
Rangecroft, S., A. J. Suggitt, K. Anderson & S. Harrison. 2016. Future climate warming and changes to mountain permafrost in the Bolivian Andes. Climatic Change 137: 231–243.
Ripley, B., B. Venables, D.M. Bates, K. Hornik, P. Albrecht & D. Firth. 2020. MASS: Support functions and datasets for Venables and Ripley’s MASS. (4th ed). R-Packages.
Sarmiento, J. & S. Barrera. 1997. Observaciones preliminares sobre la ictiofauna de la vertiente oriental andina de Bolivia. Revista Boliviana de Ecología y Medio Ambiente 2: 77–99.
Sarmiento, J., R. Bigorne, Carvajal-Vallejos, M. Maldonado, E. Leciak & T. Oberdorff. 2014. Peces de Bolivia. Plural, La Paz.
Takahashi, T. & E. Moreno. 2015. A RAD-based phylogenetics for Orestias fishes from Lake Titicaca. Molecular Phylogenetics and Evolution 93: 307–317.
Valdivia, C., J. Thibeault, J.L. Gilles, M. García & A. Seth. 2013. Climate trends and projections for the Andean Altiplano and strategies for adaptation. Advances in Geosciences 33: 69–77.
Vila, I., P. Morales, S. Scott, E. Poulin, D. Véliz, C. Harrod & M.A. Méndez. 2013. Phylogenetic and phylogeographic analysis of the genus Orestias (Teleostei: Cyprinodontidae) in the southern Chilean Altiplano: the relevance of ancient and recent divergence processes in speciation: phylogeny of Chilean Orestias. Journal of Fish Biology 82: 927–943.
Vila, I., R. Pardo & S. Scott. 2007. Freshwater fishes of the Altiplano. Aquatic Ecosystem Health & Management 10: 201–211.
Vila, I., S. Scott, N. Lam, P. Iturra & M.A. Mendez. 2010. Karyological and morphological analysis of divergence among species of the killifish genus Orestias (Teleostei: Cyprinodontidae) from the southern Altiplano. pp 471–480. En: Nelson, J.S., S. Hans & M.V.H. Wilson (eds.) Origin and Phylogenetic Interrelationships of Teleosts, Verlag Dr. Friedrich Pfeil, Munich.
Villwock, W. 1986. Speciation and adaptive radiation in Andean Orestias fishes. pp 387–403. En: Vuilleumier, F. & M. Monasterio (eds.) High Altitude Tropical Biogeography, Oxford University Press: American Museum of Natural History, Nueva York.
Wasson, J.-G., S. Barrera, B. Barrère, D. Binet, D. Collomb, I. Gonzales, F. Gourdin, J.-L. Guyot & G. Rocabado. 2002. Hydro-ecoregions of the Bolivian Amazon: A geographical framework for the functioning of river ecosystems. pp 69–91. En: McClain, M.E. (ed.) The Ecohydrology of South American Rivers and Wetlands. International Association of Hydrological Sciences, Wallingford.
Watson, D.J. & E.K. Balon. 1984. Ecomorphological analysis of fish taxocenes in rainforest streams of northern Borneo. Journal of Fish Biology 25: 371–384.
Webb, P.W. 1984. Body form, locomotion and foraging in aquatic vertebrates. American Zoologist 24: 107–120.

