Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
 Citado por SciELO
Citado por SciELO Links relacionados
 Similares en
SciELO
Similares en
SciELO  uBio
uBio Compartir

 Permalink
PermalinkEcología en Bolivia
versión impresa ISSN 1605-2528versión On-line ISSN 2075-5023
Ecología en Bolivia v.55 n.3 La Paz dic. 2020
Artículo
Modelo de distribución espacial de la almendra chiquitana (Dipteryx alata Vogel, Fabaceae) en Bolivia
Spatial distribution model of the almendra chiquitana (Dipteryx alata Vogel, Fabaceae) in Bolivia
Consuelo A. Pérez-Cruz 1 & Daniel Villarroel 2,3*
1Escuela Militar de Ingeniería, Unidad Académica Santa Cruz, 3er Anillo, Radial 13, Santa Cruz, Bolivia
2Fundación Amigos de la Naturaleza, km 7.5 carretera a la Guardia, Santa Cruz, Bolivia
3Museo de Historia Natural Noel Kempff Mercado, Universidad Autónoma Gabriel René Moreno, Av. Irak 565, Santa Cruz, Bolivia.
*Autor de correspondencia: danielvillarroel81@hotmail.com
Recibido: 07.05.20, Aceptado: 17.07.20.
Resumen
La almendra chiquitana (Dipteryx alata Vogel), es un recurso forestal no maderable que actualmente es aprovechado por las comunidades indígenas y originarias en algunas zonas de la Chiquitania, cuya demanda en el mercado nacional e internacional se ha incrementado anualmente. En el presente estudio generamos un modelo de distribución espacial de D. alata para la región de las tierras bajas de Bolivia, el cual fue obtenido mediante el consenso de los modelos generados a partir de tres métodos (Maxent, Random Forest y Support Vector Machine), 96 registros de la especie y cuatro factores ambientales. La eficiencia de predicción de los modelos generados fue evaluada mediante el método de cross-validation y los valores de AUC. Como resultado, todos los modelos generados presentaron valores de AUC entorno de 0.92, no existiendo diferencias significativas entre los métodos utilizados, por lo cual, todos alcanzaron un alto desempeño de predicción. De acuerdo con el modelo de consenso binario (presencia/ausencia), la distribución potencial de D. alata en Bolivia se extiende sobre sobre 71.200 km2 de superficie, las cuales como producto de la deforestación ocurrida hasta el 2018 habría disminuido en 23.1%, siendo el departamento de Santa Cruz el más afectado. Por tanto, este modelo de distribución potencial se constituye en uno insumo importante para la gestión de su conservación, así como para la identificación de nuevas áreas de manejo y aprovechamiento.
Palabras clave : Chiquitania, Conservación, Modelos de distribución de especies, Recurso no maderable.
Abstract
The 'almendra chiquitana' (Dipteryx alata Vogel), it's a non-timber forest resource that is currently used by indigenous and native communities in some areas of the Chiquitania region, whose demand in the national and international market has increased annually. In the present study we generated a spatial distribution model of D. alata for the Bolivian lowlands region generated by the consensus of the models obtained by applying three methods (Maxent, Random Forest and Support Vector Machine), 96 records of the species and four environmental factors. The prediction efficiency of the generated models was evaluated using the cross-validation method and the AUC values. As a result, all the models generated presented AUC values around 0.92, and there were no significant differences between the methods used, therefore, all of them achieved a high prediction performance. According to the binary consensus model (presence / absence), the potential distribution of D. alata in Bolivia extends over 71.200 km2 of surface, having been reduced by 23.1% due to deforestation until 2018, the department of Santa Cruz the most affected. Therefore, this potential distribution model constitutes an important input for the management and conservation of the species, as well as for the identification of new areas of management and sustainable use.
Keywords: Chiquitania, Conservation, Non-timber resources, Species distribution models.
Introducción
Dipteryx alata Vogel (Fabaceae) es una especie arbórea ampliamente distribuida en la ecorregión del Cerrado (sistema de clasificación basado en Dinerstein et al. 2017), siendo bastante frecuente en Brasil (Ratter et al. 2000, 2003, Sano et al. 2004, Lima & de Lima 2015), relativamente común en Bolivia (Base de datos Tropicos, Mostacedo et al. 2015, Villarroel 2015) y aparentemente raro en Paraguay, ya que hasta ahora sólo se han registrados pocas colectas botánicas (Instituto de Botánica Darwinion, Natural History Museum). Dentro de la ecorregión del Cerrado, D. alata habita principalmente en las fisionomías vegetales del cerrado sensu stricto y cerradão (Ratter et al. 2000, 2003, Sano et al. 2004, Villarroel 2015, Villarroel et al. 2015, 2016), además de zonas donde estas fisionomías entran en contacto con los bosques de la ecorregión de la Amazonia, Caatinga, Mata Atlántica y el Chaco seco (Tropicos , Sano et al. 2004, Lima & de Lima 2015). Otras de las ecorregiones donde también se han registrados individuos de D. alata son, las Sabanas del Beni, con especímenes colectados en su extremo norte, en Bolivia y Perú (Tropicos), creciendo en fisionomías vegetales similares a las del Cerrado sensu stricto; y la ecorregión del Chaco Seco, en la fisionomía de sabanas denominadas en Bolivia como Abayoy (Mostacedo et al. 2015, Villarroel et al. 2015, Villarroel et al. 2016).
Ecológicamente, D. alata es una especie típica de zonas donde la estacionalidad está bien diferenciada (entre época seca y húmeda), por lo que generalmente llega a perder sus hojas por un corto periodo durante finales de la época seca (brevidecidua). Es categorizada como heliófita durable, típica de fisionomías vegetales con dosel arbóreo discontinuo (Cerrado sensu stricto) y/o frecuentemente con claros (cerradão). Al ser intolerante a la sombra, para un óptimo desarrollo, la especie requiere de una alta y constante radiación solar. Crece principalmente sobre suelos bien drenado, ácidos, y de mediana a baja fertilidad (Sano et al. 2004, Mostacedo et al. 2015, Villarroel 2015, Villarroel et al. 2016). En Bolivia, D. alata, popularmente conocida como almendra chiquitana, es una especie que ha cobrado una alta relevancia socioeconómica, especialmente para las comunidades indígenas y originarias de la región chiquitana (Vennetier et al. 2012, Coimbra 2016, Delgado et al. 2018), pues el aprovechamiento y comercialización de sus frutos y semillas se ha convertido en una fuente adicional de recursos económicos para dichas familias (Delgado et al. 2018, Vicente & Otondo 2018); pero además, también se ha posicionado como una especie estratégica para la conservación de la biodiversidad, ya que su cosecha no implica el cambio de uso del suelo, incrementando el valor de la vegetación natural. De acuerdo con Delgado et al. (2018), a nivel nacional anualmente la demanda interna de semillas de la almendra chiquitana oscila en torno de 2.3 toneladas; así como cada año la demanda potencial de exportación alcanza aproximadamente a 42 toneladas de semillas.
Considerando la demanda interna y externa de semillas de la almendra chiquitana, y su implicancia como un recurso forestal no maderable estratégico para la conservación de la biodiversidad y el mejoramiento de los medios de vida de las comunidades indígenas y originarias, es que, en el presente trabajo generamos un modelo de distribución espacial de D. alata para la región de las tierras bajas de Bolivia, con la finalidad de identificar zonas con actitudes ambientales adecuadas para la presencia y/o desarrollo de la especie, así como su conservación y manejo.
Área de estudio
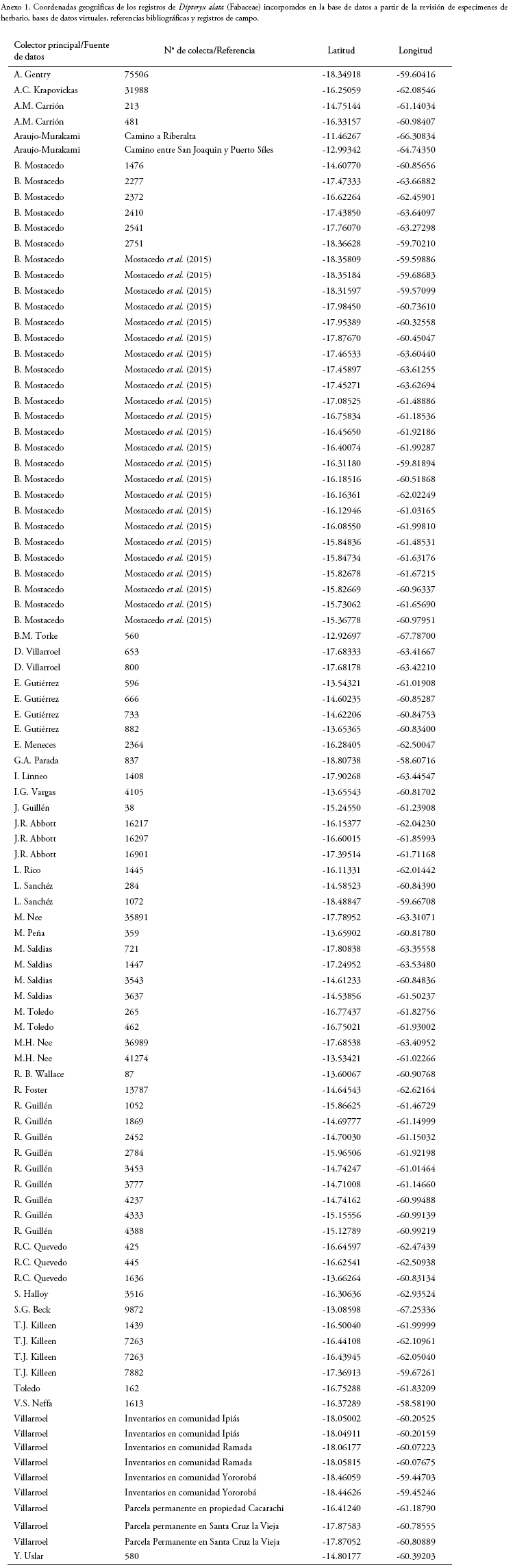
La extensión geográfica del área de estudio abarcó la región denominada como las zonas o tierras bajas de Bolivia (Ibisch et al. 2003, Beck 2015), la cual se extiende por debajo de los 1.000 m de altitud (Fig. 1). De acuerdo con Beck (2015), esta región ocupa aproximadamente el 66.4% de todo el territorio nacional, extendiéndose principalmente sobre las unidades fisiográficas siguientes: 1. las serranías subandinas, la cual se distribuye paralelamente a la cordillera de los Andes, por encima de los 500 m de elevación y cuyos sustratos son procedentes de distintos periodos geológicos; 2. las llanuras (también denominada como llanura Chaco-beniana), la cual climáticamente es divida en una porción húmeda (extremo norte, llanos de Moxos y el Pantanal) y otra seca (región del Chaco, al sur del país), en ambos casos por debajo de los 500 m y está conformado principalmente por sustratos de origen cuaternario; 3. el Escudo Brasileño, la cual representa el límite oeste de esta formación geológica de origen precámbrico, y cuya topografía corresponde a una peniplanicie o llanura ondulada; y 4. las serranías chiquitanas, las cuales están conformadas por un conjunto de cadenas montañosas y mesetas cuyas elevaciones contrastan en el paisaje, representando así las zonas más elevadas de toda la región oriental (Monte de Oca 1995, Rafiqpoor & Ibisch 2003, Villarroel et al. 2016).
Dentro de esta región, el clima varía gradualmente entre un extremo y otro (Ibisch et al. 2003, Beck 2015), lo cual se refleja en la actividad fotosintética de la vegetación debido a la estacionalidad pluvial (Fig. 1), así como en el cambio de ecorregiones (Dinerstein et al. 2017) (Fig. 1), siendo húmedo y cálido en el extremo norte y contorno este y oeste, donde dominan los bosques amazónicos (precipitación: 1-500-2.400 (>3.000) mm/año; 1-3 meses secos; temperatura promedio anual de 24-27°C, min-máx: 9-33°C), pasando a subhúmedo y cálido en la región de las sabanas del Beni, el bosque chiquitano y el Cerrado (precipitación: 600-2.300 mm/año; 3-8 meses secos; temperatura promedio anual de 21-28°C, min-máx: 3-40°C) y finalmente, seco y cálido en el extremo sur, región dominada por el bosque chaqueño (precipitación: 200-600 mm/año; 6-10 meses secos; temperatura media anual de 25-28°C, min-máx: 0-48°C).
Métodos
La generación del modelo de distribución espacial de D. alata fue realizada mediante la ejecución sucesiva de cinco etapas de análisis, específicamente como sigue:
Generación de la base de datos de presencia de la especie; construida a partir de: 1. la revisión de los especímenes botánicos de D. alata depositados en el Herbario del Oriente Boliviano (USZ); 2. la información contenida en bases de datos virtuales disponibles, específicamente, de Global Biodiversity Information Facility (GBIF; www.gbif.org) y del Jardín Botánico de Missouri (www.tropicos.org); y 3. los registros de D. alata resultante de los inventarios cuantitativos realizados por Mostacedo et al. (2015) y registros de campo proporcionados por Alejandro Araujo-Murakami y Daniel Villarroel (investigadores asociados del MHNNKM). La georreferenciación de cada uno de los registros incorporados en la base de datos fue revisada, eliminando los elementos duplicados (duplicatas) y/o realizando la corrección de la ubicación geográfica cuando fuese necesario en función de la descripción del lugar de colecta y habitat de la especie.
Obtención y selección de variables ambientales; cuatro de las 19 variables bioclimáticas (bio 1= temperatura promedio anual; bio 11= temperatura del trimestre más frío; bio 12= precipitación anual; bio 15= estacionalidad de la precipitación) modeladas por Fick & Hijmans (2017) fueron descargadas de la plataforma virtual de Worldclim (www.worldclim.org, versión 2, 19702000); estas variables fueron seleccionadas inicialmente debido a la relevancia ecológica que representan para la especies según Mostacedo et al. (2015) y Villarroel et al. (2015), así como también, por ser parte de las ocho variables bioclimáticas consideradas menos correlacionadas en Bolivia (Naoki et al. 2006). Así también, a estas cuatro variables bioclimáticas se integraron, un modelo de elevación digital (ASTER; Global Digital Elevation Model - GDEM), cuya cartografía fue descargada de la plataforma GeoBolivia (www.geo.gob.bo); y un mapa del índice de Vegetación de Diferencia Normalizada (NDVI) correspondiente a la estación seca (junio a septiembre) de los años 2015-2018 (generadas y descargadas mediante la plataforma de Google Earth Engine, https://earthengine.google.com, mediante el uso de imágenes Lansat 8). Estas seis variables ambientales fueron reajustadas a pixeles de 100 x 100 m y posteriormente evaluadas mediante el cálculo del Factor de Inflación de Varianza (VIF) con la finalidad de valorar la relación de las variables predictoras y evitar la multicolinealidad en el análisis (VIF < 10; r < 0.8). La evaluación de las variables fue realizada mediante el algoritmo VIF del paquete "usdm - Uncertainty Analysis for Species Distribution Models" (Naimi 2017).
Generación y ajustes de modelos; iniciando con la construcción de la matriz de datos ambientales a partir de los puntos de presencia y de 500 puntos de ausencia de la especie (background), proceso que fue realizado mediante la aplicación del método gRandom contenida en el algoritmo sdmData del paquete "sdm - Species Distribution Modelling" (Naimi & Araujo 2016). Así también, basados en el mismo algoritmo y una vez obtenida la matriz de datos, esta fue empleada para realizar el ajuste de los modelos de distribución, utilizando para este fin tres métodos de tipo discriminante, siendo estos, el método clásico de Máxima Entropía (Maxent) (Phillip et al. 2006), y los métodos de aprendizaje automatizado de Random Forest (RF; Breiman 2001) y Support Vector Machines (SVM; Vapnik 1995, 1998), estos últimos relativamente recién incorporados como alternativas para la generación de modelos de distribución, habiendo demostrado una alta eficiencia de predicción (Naimi & Araújo 2016, Zurell et al. 2020).
Validación y evaluación de los modelos; los modelos fueron validados mediante la denominada Receiver Operating Characteristic (ROC), la cual resulta de la relación entre, la tasa de identificación correcta de presencias verdaderas (sensitivity/sensitividad, eje "y") vs. la tasa de identificación de falsas presencias (1-specificity/especificidad, eje "x"). La eficiencia de las curvas ROC fue medida en base al valor del Area Under the Curve (AUC; mientras más cercano a 1, mayor será la capacidad de predicción) (Phillips et al. 2006). La evaluación de los modelos fue realizada con el método de cross-validation (Roberts et al. 2017), para lo cual, la matriz de datos fue particionada en cinco grupos (cuatro de entrenamiento y uno de prueba), con los cuales, simultánea y sucesivamente se generaron cinco nuevos modelos por cada uno de los métodos empleados, así como también sus respectivos límites de corte (threshold). Finalmente, los valores de AUC obtenidos con los algoritmos aplicados fueron contrastados mediante un análisis de varianza (ANOVA).
Predicción y ensamble de modelos; la predicción de la distribución potencial de D. alata fue espacializada en base a los 15 modelos generados según el método de cross-validation (cinco por cada método). Estos 15 modelos de predicción de distribución fueron ensamblados mediante el cálculo del promedio ponderado (weighted en inglés), generando de esta forma tres nuevos modelos, uno por cada método empleado (SVM, RF, Maxent). Después, cada uno de estos modelos fue diferenciado binariamente (presencia/ausencia) según sus respectivos límites de corte (threshold en inglés), para así posteriormente obtener un modelo de consenso de probabilidad de ocurrencia de D. alata. Con la generación del modelo de consenso, el cual representa la sinergia probabilística de las predicciones de tres métodos, se espera representar espacialmente las zonas con mayor probabilidad de presencia de la especie, pues a priori, éste resulta más confiable que los modelos basados en un solo método (Araújo & New 2007, Mateo et al. 2010, Naimi & Araújo 2016, Zurell et al. 2020). Finalmente, el mapa de consenso fue contrastado con el mapa de deforestación propuesto por Hansen et al. 2013, para estimar la disminución del área potencial de distribución de la especie por efecto de la pérdida reciente de bosque.
El análisis de validación (algoritmo sdmData), evaluación (algoritmo getEvaluation), predicción (algoritmo predict) y ensamble de modelos (algoritmo ensemble), fueron realizados con el paquete "sdm - Species Distribution Modelling" (Naimi & Araujo 2016) en el software R (R Core Team 2020, versión 3.6.3).
Resultados
Datos generales
La base de datos compilada para el análisis estuvo conformada por 96 registros de D. alata, de las cuales, 92 fueron obtenidos en el departamento de Santa Cruz, y los cuatro restantes correspondieron a registros del Norte del departamento La Paz y el Beni (Anexo 1). De las seis variables ambientales, dos fueron excluidas (bio 1; bio 11), ya que éstas presentaron una elevada redundancia (correlación) con el resto de las variables predictores. Por lo que, con la exclusión de estas dos variables se estaría evitando la posible generación modelos sesgados producto de la colinealidad de los datos ambientales (Anexo 2).
Modelos de distribución de la especie
Todos los modelos de distribución potencial de D. alta alcanzaron un alto desempeño de predicción con valores de AUC promedio entorno de 0.92 y variaciones entre 2.6 y 4.8%, no existiendo diferencias notoriamente discrepantes entre las eficiencias de predicción según el análisis de varianza (f= 0.015, p-value = 0.985, tabla 1, Fig. 2).
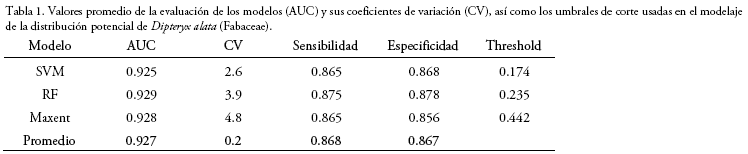
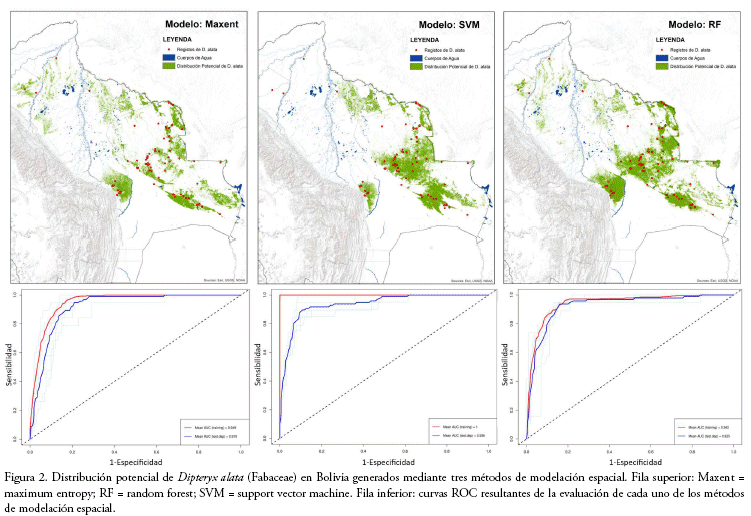
Los umbrales de corte de los modelos cambiaron en función del método utilizado (Tabla 1, Fig. 2), ya que, en la construcción de los modelos de distribución, cada uno de los métodos atribuye un peso o importancia diferente a las variables ambientales (Tabla 2).
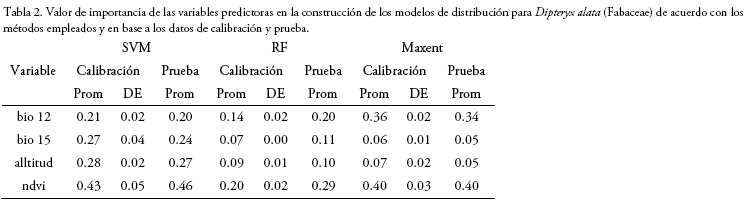
Los modelos generados mediante los tres métodos habrían demostrado tener una alta exactitud estadística en la distinción entre las áreas con presencia y/o ausencia de la especie. En todos los modelos, el NDVI fue la variable ambiental de mayor importancia para la predicción de la distribución de D. alata, así como también, bio 12 para los métodos de RF y Maxent; y, la elevación y bio 15 para SVM. Los valores de importancia de las variables ambientales correspondientes a los registros utilizados para el entrenamiento y evaluación de los modelos resultaron similares (Tabla 2). De acuerdo con el ensamble y consenso de los modelos de distribución potencial, en Bolivia, D. alata podría llegar a extenderse sobre aproximadamente 71.200 km2 de superficie (Fig. 3), abarcando una mayor extensión en los departamentos de departamento de Santa Cruz (82.6%) y el Beni (15.8%), así como también en los departamentos de La Paz (1.6%) y Pando (0.1%), aunque en una menor proporción.
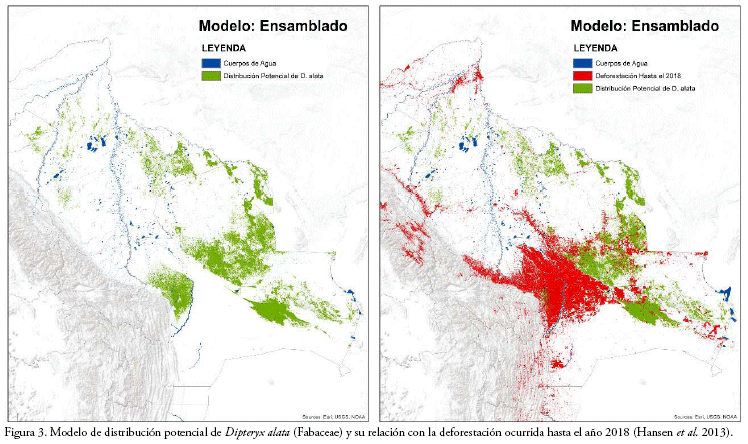
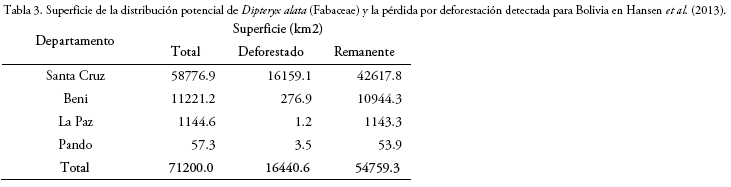
Como consecuencia de la deforestación ocurrida en Bolivia hasta finales de 2018 (estimación basada en el mapa de deforestación por de Hansen et al. 2013), el área de distribución potencial de D. alata habría disminuido en 23.1%, lo cual, indica que su extensión de distribución remanente se redujo a 54.759 km2 (Fig. 3). Del total de la superficie disminuida por la deforestación, el 98.3% ocurrió en el departamento de Santa Cruz, lo cual es equivalente al 27.5% de toda la extensión potencial estimada dentro de dicho departamento (Tabla 3; Fig. 3). Por otro lado, en los departamentos del Beni, La Paz y Pando, la pérdida de la extensión potencial de D. alata habría sido mínima (Tabla 3), por lo que, dichas regiones podrían estar conservando una elevada proporción de su distribución natural (Fig. 3).
Discusión
Los modelos de distribución de especies son generados con la finalidad de espacializar geográficamente zonas con alta y/o baja idoneidad ambiental, donde, probabilísticamente un taxón podría o no estar presente, siendo el resultado de la relación entre, la ubicación geográfica de los registros de presencia del taxón (variable respuesta; presencia verdadera) y las características ambientales del sitio ocupado ("variables predictoras") (Mateo et al. 2011). Ninguna técnica o método matemático de predicción ha resultado ser completamente perfecto, pero si han demostrado tener una mayor y/o menor eficiencia de predicción cuando comparados unos con otros (Naoki et al. 2006; Mateo et al. 2011). Por otro lado, la eficiencia de predicción de los modelos de distribución dependerá estrictamente de la calidad y del número de muestras y variables predictoras a ser utilizadas (Naoki et al. 2006, Peterson & Nakazawa 2008, Mateo et al. 2010), así como de la escala geográfica del área de estudio (Manzoor et al. 2018). Bajo este contexto, los modelos de distribución del presente estudio fueron generados y evaluados considerando un adecuado número de muestras de presencia, pues, al contar con una base de datos de 96 registros de D. alata se estaría superando el número mínimo de presencias indicado por Mateo et al. (2010) (18 a 20 presencias), tanto para el entrenamiento como para las pruebas.
La decisión de trabajar con pixeles de 100 m (una resolución menor a la que generalmente es utilizada, 1 km) fue basada en las consideraciones expresadas recientemente por Manzoor et al. (2018) en relación a la transferibilidad de los modelos según la biología de la especie (generalistas y no generalistas). Al ser D. alata una especie no generalista (Mostacedo et al. 2015, Villarroel 2015, Villarroel et al. 2016), su distribución podría responder también en mayor proporción a variables biofísicas y otras como heterogeneidad del paisaje y, tipo y extensión de cobertura vegetal, las mismas que en la región del área de estudio pasarían desapercibidas si se utilizaran pixeles de 1 km, pues, éstas variables pueden cambiar drásticamente a cortas distancias, tal como lo describe Villarroel et al. (2016).
Con la exclusión de las variables predictoras bio 1 y bio 11 (ambas correlacionadas entre sí, así como también con las otras cuatro variables predictoras), todos los modelos generados estuvieron basados estrictamente en función de variables predictoras independientes, libres de multicolinealidad, con lo cual se habría disminuido la probabilidad de obtener modelos poco fiables y con valores significativos de errores de comisión u omisión (Naoki et al. 2006, Peterson & Nakazawa 2008, Mateo et al. 2010, Manzoor et al. 2018).
La importancia de las variables predictoras finalmente utilizadas para la construcción de los modelos (Tabla 2) concuerdan con las características ecológicas descritas para D. alata y el tipo de hábitat donde se desarrolla (Sano et al. 2004, Mostacedo et al. 2015, Villarroel 2015, Villarroel et al. 2016). Los valores de NDVI entre 0,4 y 0,6 durante la estación seca generalmente indican coberturas vegetales deciduas, semideciduas o con dosel discontinuo, similares a las del cerrado sensu stricto y cerradão; así como también, concuerdan con las características fisiológicas de la especie, ya que ésta es brevidecidua (pierde sus hojas por un corto periodo de tiempo al finalizar la época seca), momento que además marca el inicio de su floración y la finalización de la estación seca. Por lo cual, para la generación de estas condiciones ambientales, es necesario un escenario con dos estaciones climáticas diferenciadas (bio 15; estación seca y húmeda), y una precipitación anual (bio 12) superior a la del Chaco (>400900 mm/año) e inferior al de la Amazonia (<1.200-3.000 mm/año) (Ibisch et al 2003, Beck 2015).
La pérdida de hábitat natural de D. alata como producto de la deforestación, es una amenaza creciente para el estado de conservación de esta especie en el país. Aunque actualmente está categorizada como vulnerable (VU) a nivel global por la IUCN, en Bolivia no ha sido incluida en ninguna de las listas de especies bajo riesgo o amenaza de conservación (Meneses & Beck 2005, Mamani et al 2010, Ministerio de Medio Ambiente y Agua 2020). Es necesario reevaluar su estado de conservación a nivel nacional dada la relevancia económica que su aprovechamiento representa para las comunidades indígenas y originarias, especialmente en la región chiquitana (Vennetier et al. 2012, Coimbra 2016, Delgado et al. 2018), donde la cosecha y comercialización de sus frutos y semillas han ayudado a mejorar y diversificar los ingresos económicos de sus familias (Delgado et al. 2018, Vicente & Otondo 2018); incrementando así, el valor de la vegetación natural.
Por otro lado, al ser una especie típica de suelos ácidos y de mediana a baja fertilidad (Sano et al. 2004, Mostacedo et al. 2015, Villarroel 2015, Villarroel et al. 2016), su aprovechamiento se torna una actividad estratégica para valorizar los suelos donde la producción agrícola es casi imposible sin una alta inversión de recursos económicos, el cual no es un atributo con el que cuentan los habitantes indígenas y originarios de la región. De acuerdo con Delgado et al. (2018), la demanda interna y externa de semillas de D. alata ha mostrado un crecimiento constantemente, alcanzando un potencial de exportación cercano a las 42 toneladas anuales. Actualmente, el aprovechamiento de D. alta es una actividad practicada por las comunidades próximas a la ciudad de Concepción y San Ignacio de Velasco en el departamento de Santa Cruz (Vennetier et al. 2012, Delgado et al. 2018, Vicente & Otondo 2018), no obstante, se estima que hasta ahora no se ha conseguido llegar a cubrir ni un tercio de la demanda nacional e internacional de este producto, pues, por ejemplo, el 2018, sólo se habrían llegado a cosechar cerca de 9 toneladas (Delgado et al. 2018). La identificación de otras zonas y/o regiones cartografiadas detectadas con el modelo de distribución desarrollado en este estudio, tales como, la zona Sur del municipio de San José de Chiquitos y Roboré, la zona sur y sureste del ANMI San Matías, así como la zona centro y norte del municipio de San Ignacio de Velasco, podrían constituirse en nuevas áreas con alto potencial de aprovechamiento de la especie. Los resultados del presente estudio representan la base técnico-científica para desarrollar una estrategia regional de manejo y aprovechamiento de la especie.
Conclusiones
El consenso de los productos generados mediante los tres métodos utilizados permitió obtener un modelo de distribución que sugiere una distribución potencial de la especie en Bolivia que cubre cerca 71.200 km2 (6.5% de la superficie de Bolivia).
Este resultado se constituye en un insumo técnico-científico clave para la planificación de acciones de manejo y aprovechamiento actual y futuro de la especie, permitiendo que, por ejemplo, planificar acciones de aprovechamiento en nuevas regiones además de las tradicionalmente conocidas. Por otra parte, la pérdida del hábitat natural de la especie producto de la deforestación representa una amenaza creciente para su conservación en Bolivia.
Agradecimientos
Este trabajo de investigación fue realizado en el marco del proyecto "Improving indigenous Bolivian Chiquitano people's livelihoods through sustainable forest management", el cual es financiado por la Iniciativa Darwin (proyecto # 26-024, Royal Botanic Gardens, Kew). Así también, el primer autor agradece a la Fundación Amigos de la Naturaleza y al Museo de Historia Natural Noel Kempff Mercado por brindarle el apoyo técnico y logístico para la generación de este manuscrito. Agradecemos a los revisores anónimos que contribuyeron a mejorar esta publicación.
Referencias
Araújo, M.B. & M. New. 2007. Ensemble forecasting of species distributions. Trends in Ecology & Evolution 22(1): 42-47.
Beck, S.G. 2015. Las regiones y zonas de vegetación. pp 3-20. en: Jørgensen, P.M., M.H. Nee & S.G. Beck (eds.) Catálogo de las Plantas Vasculares de Bolivia. Missouri Botanical Garden Press, St. Louis, Missouri.
Breiman, L. 2001. Random Forests. Machine Learning 45(1): 5-32. [ Links ]
Coimbra, D.J. 2016. Almendra Chiquitana (Dipteryx alata): Guía para su aprovechamiento, manejo y cultivo. Fundación para la Conservación del Bosque Chiquitano, Santa Cruz. [ Links ]
Delgado, R., P. Navarro & R. Trujillo. 2018. Estudio sobre el estado actual y avances en las cadenas productivas del cusi, copaibo y almendra chiquitana. Informe no publicado, Fundación Amigos de la Naturaleza, Proyecto Gestión Integral de Bosques para la Reducción de la Deforestación/GIZ, Santa Cruz.
Dinerstein, E., D. Olson, A. Joshi, C. Vynne, N.D. Burgess, E. Wikramanayake, N. Hahn, S. Palminteri, P. Hedao, R. Noss, M. Hansen, H. Locke, E.C. Ellis, B. Jones, C.V. Barber et al. 2017. An ecoregion-based approach to protecting half the terrestrial realm. BioScience 67(6): 534-545.
Fick, S.E. & R.J. Hijmans. 2017. WorldClim 2: new 1-km spatial resolution climate surfaces for global land areas. International Journal of Climatology 37(12): 4302-4315.
Hansen, M.C., P.V. Potapov, R. Moore, M. Hancher, S.A. Turubanova, A. Tyukavina, D. Thau, S.V. Stehman, S.J. Goetz, T.R. Loveland, A. Kommareddy, A. Egorov, L. Chini, C.O. Justice & J.R.G. Townshend. High-resolution global maps of 21st-century forest cover change. Revisado última vez: 30 de mayo de 2020. Sitio Web: http://earthenginepartners.appspot.com/science-2013-global-forest
Ibisch, P.L., S.G. Beck, B. Gerkmann & A. Carretero. 2003. Ecoregiones y ecosistemas. pp 47-88. en: Ibisch, P.L. & G. Mérida (eds.) Biodiversidad: La Riqueza de Bolivia, Estado de Conocimiento y Conservación, Fundación Amigos de la Naturaleza, Santa Cruz.
Instituto de Botánica Darwinion. Version 1.0. Revisado última vez: 28 de abril de 2020. Sitio Web: https://doi.org/10.15468/vtfbe3 [ Links ]
Lima, H.C. & I.B. de Lima. Dipteryx. Flora do Brasil 2020 em construção. Revisado última vez: 28 abr. 2020. http://floradobrasil.jbrj.gov.br/jabot/floradobrasil/FB29628
Mamani, F., P. Pozo, D. Soto, D. Villarroel & J.R.I. Wood. 2010. Libro rojo de las plantas de los cerrados del Oriente Boliviano. Museo de História Natural Noel Kempff Mercado - Darwin Initiative, Santa Cruz.
Manzoor, S.A., G. Griffiths & M. Lukac. 2018. Species distribution model transferability and model grain size – finer may not always be better. Scientific Reports 8(1): 7168.
Mateo, R.G., Á.M. Felicísimo & J. Muñoz. 2010. Effects of the number of presences on reliability and stability of MARS species distribution models: the importance of regional niche variation and ecological heterogeneity. Journal of Vegetation Science 21(5): 908-922.
Mateo, R.G., Á.M. Felicísimo & J. Muñoz. 2011. Modelos de distribución de especies: Una revisión sintética. Revista Chilena de Historia Natural 84: 217-240.
Montes-de-Oca, I. 1995. Geografía y clima de Bolivia. Bulletin de l'Institut Français d'Études Andines 24(3): 357-368. [ Links ]
Meneses, R.I. & S.G. Beck. 2005. Especies amenazadas de la flora boliviana. Herbario Nacional de Bolivia, La Paz. Informe no publicado.
Ministerio de Medio Ambiente y Agua. 2020. Libro rojo de plantas amenazadas de las tierras bajas de Bolivia. Fundación Amigos de la Naturaleza, Santa Cruz. [ Links ]
Mostacedo, B., B. Avendaño, D. Villarroel & D. Espinoza. 2015. Factores que afectan la distribución de la almendra chiquitana. pp 46-51. en: Mostacedo, B. & D. Villarroel (eds.) Identificación de Variedades, Ecología y Productividad de la Almendra Chiquitana (Dipteryx alata). Dirección Universitaria de Investigación, Universidad Autónoma Gabriel René Moreno, Santa Cruz.
Naimi, B. & M.B. Araujo. 2016. sdm: a reproducible and extensible R platform for species distribution modelling. Ecography 39(4): 368-375.
Naoki, K., M.I. Gómez, R.P. López, R.I. Meneses & J. Vargas. 2006. Comparación de modelos de distribución de especies para predecir la distribución potencial de vida silvestre en Bolivia. Ecología en Bolivia 41(1): 65-78.
Natural History Museum. Collection specimens. Revisado última vez: 28 de abril de 2020. Sitio Web: https://doi.org/10.15468/vtfbe3 [ Links ]
Peterson, A.T. & Y. Nakazawa. 2008. Environmental data sets matter in ecological niche modelling: an example with Solenopsis invicta and Solenopsis richteri. Global Ecology and Biogeography 17(1): 135-144.
Phillip, S.J., R.P. Anderson & R.E. Schapired. 2006. Maximum entropy modeling of species geographic distributions. Ecological Modelling 190: 231–259.
Rafiqpoor, D. & P.L. Ibisch. 2003. El medio físico. pp 4-17. En: Ibisch, P.L. & G. Mérida (eds.) Biodiversidad: La Riqueza de Bolivia, Estado de Conocimiento y Conservación. Fundación Amigos de la Naturaleza, Santa Cruz.
Ratter, J.A., S. Bridgewater, J.F. Ribeiro, T.A.B. Dias & R.M. Silva. 2000. Estudo preliminar da distribuição das espécies lenhosas da fitofisionomia Cerrado sentido restrito nos estados compreendidos pelo Bioma Cerrado. Boletim do Herbârio Ezequias Paulo Heringer 5: 5-43.
Ratter, J.A., S.B. Wridgewater & J.F. Ribeiro. 2003. Analysis of the floristic composition of the Brazilian Cerrado vegetation III: Comparison of the woody vegetation of 376 Areas. Edinburgh Journal of Botany 60(1): 57-109.
Roberts, D.R., V. Bahn, S. Ciuti, M.S. Boyce, J. Elith, G. Guillera-Arroita, S. Hauenstein, J.J. Lahoz-Monfort, B. Schröder, W. Thuiller, D.I. Warton, B.A. Wintle, F. Hartig & C.F. Dormann. 2017. Cross-validation strategies for data with temporal, spatial, hierarchical, or phylogenetic structure. Ecography 40(8): 913-929.
Sano, S.M., J.F. Ribeiro & M.A. Brito. 2004. Baru: biologia e uso. Embrapa Cerrados, Ministerio da Agricultura, Pecuaria e Abastecimento Documento 116, Brasilia DF.
Tropicos - Data base. Missouri Botanical Garden. Revisado última vez: 28 abril de 2020. Sitio Web: http://www.tropicos.org/NameSearch.aspx
Vapnik, V. 1995. The nature of statistical learning theory. Springer-Verlag, Nueva York. [ Links ]
Vapnik, V. 1998. Statistical learning theory. John Wiley and Sons, Inc., Nueva York. [ Links ]
Vennetier, C., R. Peltier & J. Coimbra. 2012. Valorizar la almendra chiquitana, Dipteryx alata Vogel: ¿Una estrategia para mitigar el impacto ambiental del desarrollo agropecuario en Bolivia? Bois et Forêts des Tropiques 311(1): 36-48.
Vicente, J.J. & A. Otondo. 2018. Dipteryx alata (Vogel): La almendra Chiquitana. Instituto Interamericano de Cooperación para la Agricultura – Programa Cooperativo para el Desarrollo Tecnológico Agroalimentario y AgroIndustrial del Cono Sur, La Paz.
Villarroel, D. 2015. Aspectos generales y usos de la almendra Chiquitana. pp 19-25. en: Mostacedo, B. & D. Villarroel (eds.) Identificación de Variedades, Ecología y Productividad de la Almendra Chiquitana (Dipteryx alata). Dirección Universitaria de Investigación, Universidad Autónoma Gabriel René Moreno, Santa Cruz.
Villarroel, D., B. Mostacedo, M. Toledo, M.V. Antezana & A. del Valle. 2015. Densidad, estructura poblacional y regeneración natural de la almendra chiquitana. pp 52-67. en: Mostacedo, B. & D.
Villarroel (eds.) Identificación de Variedades, Ecología y Productividad de la Almendra Chiquitana (Dipteryx alata). Dirección Universitaria de Investigación, Universidad Autónoma Gabriel René Moreno, Santa Cruz.
Villarroel, D., C.B.R. Munhoz & C.E.B. Proença. 2016. Campos y sabanas del Cerrado en Bolivia: delimitación, síntesis terminológica y sus características fisionómicas. Kempffiana 12(1): 47-80.
Zurell, D., N.E. Zimmermann, H. Gross, A. Baltensweiler, T. Sattler & R.O. Wüest. 2020. Testing species assemblage predictions from stacked and joint species distribution models. Journal of Biogeography 47(1): 101-113.