










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkEcología en Bolivia
Print version ISSN 1605-2528On-line version ISSN 2075-5023
Ecología en Bolivia vol.55 no.1 La Paz Apr. 2020
Artículo
Floral ecology and floral visitors of Puya atra (Bromeliaceae), a Bolivian endemic plant
Ecología floral y visitantes florales de Puya atra (Bromeliaceae), una planta endémica boliviana
Paola Velásquez-Noriega1*, César Mayta2, Elmer Cuba3, Emilia García E.1, Flavia Montaño-Centellas4 & Thorsten Krömer5
1Herbario Nacional de Bolivia, Instituto de Ecología, Universidad Mayor de San Andrés, c/27 Cota Cota, Campus Universitario, Casilla 1007 - Correo Central, La Paz, Bolivia.
2Departamento de Biología, Universidad de La Serena, Benavente 980, Casilla 554, La Serena, Chile.
3Simbiosis srl., Av. Arce 2105, edif Venus, 3A, La Paz, Bolivia.
4Department of Wildlife Ecology and Conservation, University of Florida, Gainesville, Florida 32611, USA.
4Centro de Investigaciones Tropicales (CITRO), Universidad Veracruzana, José María Morelos 44-46, CP 9100, Xalapa, Veracruz, Mexico.
*Autora de correspondencia: paola.vn19@gmail.com
Recibido: 07.11.19, Aceptado: 12.01.20.
Abstract
Hummingbirds and bromeliad species of the genus Puya occur exclusively in the Neotropics. Their interaction is important for the reproduction and genetic variability of these plants. To understand this relationship, it is important to study the phenology of Puya species, their floral characteristics, and their floral visitors. In this work, we studied the floral ecology of the Bolivian endemic Puya atra, which grows in the montane cloud forests of the Yungas. We made monthly visits to the study area for two years to record phenology data, to collect nectar samples, and to make direct observations of floral visitors (539 observation hours) during its three-months of flowering. In this period, P. atra produced 8.09 ± 1.59 μL of nectar per flower, which contains fructose and glucose. We confirmed that P. atra is visited by two species of longed-bill hummingbirds: Coeligena violifer and Pterophanes cyanopterus, being males of both species the main visitors. The morphology of both visitors allows contact with the reproductive parts of its flowers and the transfer of pollen, while the plant rewards hummingbirds with nectar. It seems that P. cyanopterus showed a tight synchronisation with the plant's phenology and visited more frequently with a greater abundance of flowers, showing a positive interaction with P. atra. However, it is necessary to continue with different studies on the breeding system and sugar production throughout the day of this Puya species as well as the behavioural response of these hummingbirds in its presence.
Key words: Bolivia, Montane cloud forest, Ornithophily, Phenology, Trochilidae.
Resumen
Los colibríes y especies de bromeliáceas del género Puya se distribuyen exclusivamente en el Neotrópico. Su interacción es importante para la reproducción y la variabilidad genética de dichas plantas. Para comprender esta relación, es importante estudiar la fenología de las especies de Puya, sus características y visitantes florales. En este estudio analizamos la ecología floral de la especie endémica boliviana Puya atra, que crece en el bosque nublado montano de los Yungas. Visitamos el área de estudio mensualmente durante dos años para registrar datos de fenología, colectar muestras de néctar y hacer observaciones directas de visitantes florales (539 horas de observación) durante tres meses de su floración. En este periodo, P. atra produjo 8.09± 1.59 μL de néctar por flor, el cual contenía fructuosa y glucosa. Confirmamos que P. atra es visitada por dos especies de colibríes de pico largo: Coeligena violifer y Pterophanes cyanopterus, siendo los machos los principales visitantes para ambas especies. La morfología de ambos visitantes permite el contacto con las partes reproductivas de las flores y la transferencia de polen, mientras que la planta ofrece a los colibríes néctar. Al parecer P. cyanopterus mostró una estrecha sincronización con la fenología de la planta y la visitó con mayor frecuencia cuando hay mayor abundancia de flores, mostrando una interacción positiva con P. atra. Sin embargo, es necesario continuar con diferentes estudios sobre su sistema de reproducción y la producción de azúcar a lo largo del día de esta especie de Puya, así como la respuesta comportamental de estos colibríes en su presencia.
Palabras clave : Bolivia, Bosque nublado montano, Fenología, Ornitofilia, Trochilidae.
Introduction
Angiosperms have developed several mechanisms to ensure the arrival of pollen to the stigma of the flower and the consequent fertilisation, which in many cases occurs with the participation of pollinating organisms (Dafni 1992). There are several examples of how plants attract pollinators and ways in which interspecific relationships are established (Faegri & Van der Pijl 1979). Thus, co-adaptation between pairs or groups of species is partially explained by mutualistic interactions (Jordano 1987, Thompson 2005, Bascompte & Jordano 2007). This kind of relationships occur between birds and several angiosperms, since many bird species are effective pollinators, and most plants offer nectar as the main reward for this service (Snow 1981, Proctor et al. 1996).
Bromeliaceae is a widely distributed Neotropical family with about 3600 species in 75 genera (Gouda et al. 2019). These plants are characterised by forming rosettes and having inflorescences with showy bracts and flowers in many of the species (Varadarajan & Brown 1988, Benzing 2000). Of the 321 species of Bromeliaceae in Bolivia distributed from the Andean paramo to the tropical lowlands, there are 146 endemics (Krömer et al. 2014). One example is Puya atra L.B. Sm., which grows in rocky areas and humid montane forests of the Yungas in the departments of La Paz, Cochabamba, and Santa Cruz between 2,500-3,500 m (Krömer et al. 1999, 2014). This terrestrial or saxicolous plant can form thick patches or grow as isolated individuals. According to the morphological characteristics of its flowers, and considering other similar Puya species, ornithophily by hummingbirds (Trochilidae) has been deduced as the main pollination mode for this species (Krömer et al. 2006).
Ornithophily has been reported to be the most common pollination syndrome for the family (Benzing 2000, Kessler & Krömer 2000, Krömer et al. 2008), and hummingbirds are their main visitors and pollen vectors (Canela & Sazima 2005, Krömer et al. 2006, Kessler et al. 2020). Their interaction originated in the early Tertiary period, and apparently, a (diffuse) co-adaptation between both families has occurred since then (Buzato et al. 2000, Givnish et al. 2014). This system of mutualistic interaction between bromeliads and hummingbirds is quite particular, given the endemic nature of both families in the Neo tropics.
Nectar is the most common reward that animal-pollinated plants offer to their mutualists (Proctor 1996). Glucose, fructose, and sucrose are the main components of nectar sugars, which also represent the largest amount of energy for consumers (Stiles & Freeman 1993, Baker et al. 1998, Petanidou et al. 2006). Hummingbirds are the most important group of nectarivore birds in the Neotropics (Schuchmann 1999). Also, the primary visitors of bromeliad species whose nectar is usually sucrose rich, while bromeliads pollinated by bats or passerine birds tent to secrete hexose rich nectars composed mainly of sucrose (Scogin & Freeman 1984, Baker et al. 1998, Galetto & Bernardello 2003, Krömer et al. 2008, Gottlinger et al. 2019).
Characterising the reproductive phenology of plants is very important in mutualistic interactions, because it is fundamental to understand the dynamics of the community (Williams et al. 1999). The time, duration and degree of synchrony of the different phenological phases have important implications for the quantity and quality of resources available to consumers (Williams et al. 1999, Bascompte & Jordano 2007, Elmendorf et al. 2016).
Due to the sessile nature of plants, pollination and seed dispersal are the most important processes to ensure gene flow among plant communities (Barrett & Harder 1996). Identification of key biotic dispersal agents is essential to conserve communities and ecosystems, especially for those families with many endemic species such as Bromeliaceae. In this sense, the present study aims to examine the floral ecology of the endemic bromeliad P. atra, in a cloud forest of the Cotapata National Park, La Paz, Bolivia. Specifically, we want to describe the phenology of this species, the nectar production and composition, and determine their floral visitors.
Methods
Study area
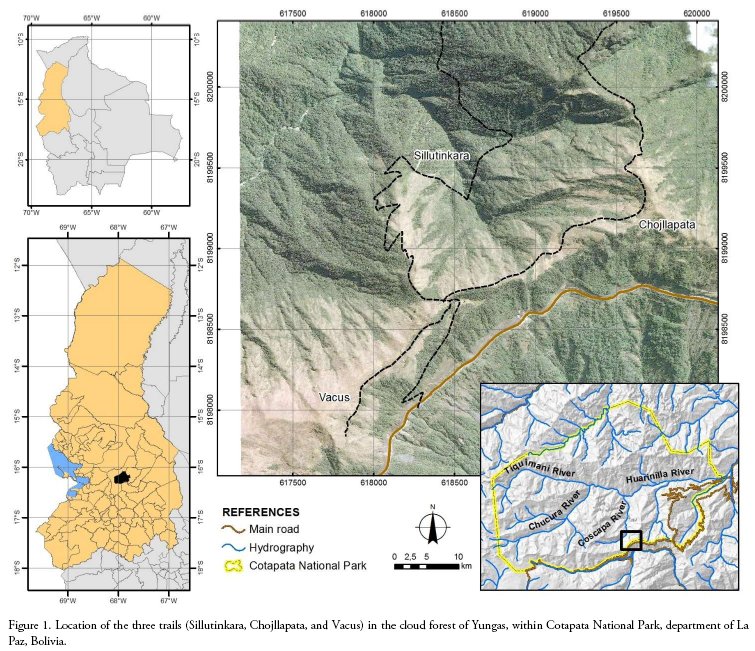
The study was conducted in Cotapata National Park and Natural Area of Integrated Management located in La Paz, Bolivia, where P. atra populations were recorded along three transects, each one along the main trails of the protected area (Sillutinkara, Chojllapata and Vacus; 67°53'28,848"W, 16°17 23,573"S, 3,000-3,500 m (Fig. 1). The park is located about 30 km NE from La Paz city. It occupies the Yungas ecoregion (Ibisch 1998), characterised by the orographic influence of winds laden with high atmospheric moisture, which causes almost permanent fog, low temperatures (between 1012°C approximately) and high levels of precipitation (more than 2500 mm). The vegetation is arboreal and shrubby, with twisted trunks and coriaceous leaves, and occurs in crests, tops and slopes of very steep mountains (Ribera 1995). This montane cloud forest has a vertical structure of three or four strata, with a continuous canopy that reaches up to 10 meters high (Meneses et al. 2004).
Species of study
In the study area, P. atra occurs on rocky slopes between 2,500 and 3,500 m (Krömer 2000). This endemic species consists of a shrubby rosette of 1 m in diameter and its inflorescence with scape up to 2 m high (Fig. 2a-c). Leaves are abundant, triangular and spiny. The inflorescence is large, cylindrical, woolly pubescent, and densely branched, with green to blue-green tubular flowers about 16-25 mm long (Smith & Downs 1974).

Plant reproductive phenology
The three transects were visited monthly from January 2016 to December 2017 to record data on the reproductive phenology of the species. The phenophase of an average of 182 individuals unevenly distributed on rocks and steep slopes across the whole three transects was recorded in the same order during each visit. Though these individuals were not marked due to difficult accessibility (Fig. 2a), they were easily identifiable and distinguishable by observation with the naked eye or use of binoculars from the transects.
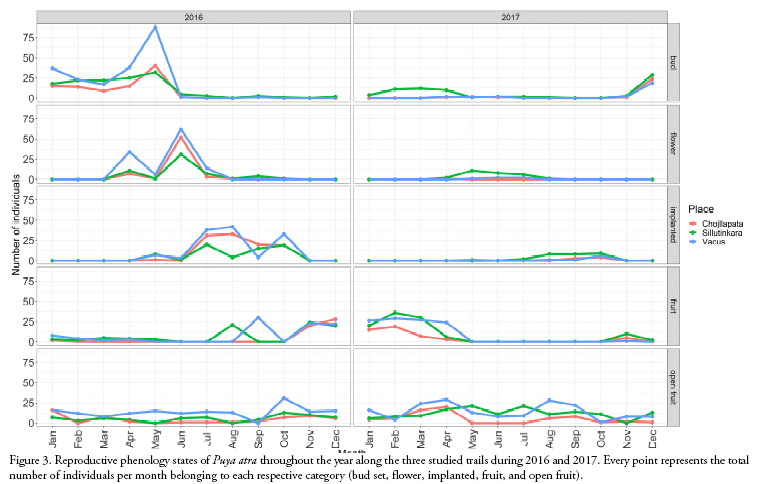
We described the phenophase as the following (Fig. 2d-g): (1) bud set, from the first appearance of the young inflorescence within the rosette, with about 13 cm until it reaches a height of 100 cm or more with scape, bracts and buds separated; (2) flowers, from the moment the first corolla leaves appear in the inflorescence, with light green or bluish-green colours; (3) flowers implanted, when all corollas have formed spirals, the stem and the petals acquire a yellow colour because they begin to wilt; (4) fruits, when the immature fruits develop; and (5) open fruits, when the dehiscent capsules have opened and they release the seeds in different periods without a particular sequence. At this time, the whole plant turns brown. With all these data, a diagram of plant phenology was elaborated (Fig. 3).
Nectar composition
Due to their easier accessibility fifteen selected inflorescences were covered with tulle bags to prevent the visitors' entry. The total volume of nectar present in each flower was harvested as soon as the flower opened between 6:30-7:00 h by inserting micro capillaries of 80 μl (Scogin & Freeman 1984). The flower was extracted from an inflorescence in order to obtain each sample of nectar, and thus we were only able to measure each flower once. To estimate the volume of nectar produced by plants, 75 nectar samples from individual flowers of these fifteen inflorescences were measured with micro capillaries. Then, the volume values of those were averaged (García & Hoc 1998). The sugar composition was measured in nectar samples of 18 flowers from these inflorescences by Nuclear Magnetic Resonance (NMR) spectroscopy in the Laboratorio de Biorganica of the Instituto de Investigaciones Químicas (at UMSA).
Floral visitors
Previous direct observations during five consecutive days and nights allowed us to determine that the only visitors of P. atra at our study area were hummingbirds, while no other diurnal or nocturnal animals such as insects or bats have been noticed. Thus, our observation was conducted without pause from 8:00-19:00 h; the duration was adjusted according to light availability and bird activity (Canela & Sazima 2005) for a total of 539 hours (49 days in total, for both years).
Before collecting the data of interactions, we identified observation areas (three at Vacus, four at Chojllapata and five at Sillutinkara). We considered an observation area of a half circle with a 20 m radius, from a point on the trail and looking to one side of the trail (i.e., left or right due to visibility). This method allowed us for an accurate identification of visitors (Ortiz-Pulido et al. 2012). Each observation area included more than two individual plants with available flowers grouped in clusters. The edges of the observation areas were at least 100 m apart from each other (Gonzalez & Loiselle 2016).
To analyse the activity of hummingbirds throughout the day, we divided the observation time in 11 one-hour intervals, in which we observed the frequency of visits for a whole hour each in the same Puya individuals (Woods & Ramsay 2001). This interval assignment allowed us to compare the daily activity patterns between hummingbird species and P. atra. Fifty-seven inflorescences were observed during 2016 (Sillutinkara: 35, Chojllapata: 10 and Vacus: 12), while only four inflorescences were observed along the Sillutinkara trail in 2017. We calculated the frequency of visits for each species, considering a visit as each event in which a hummingbird individual sipped nectar from a plant by inserting its bill into a flower within a one-hour interval (Vázquez et al. 2005).
Results
Two species of hummingbirds (Coeligena violifer Gould 1846, only males and Pterophanes cyanopterus Fraser 1839, both sexes) were recorded visiting 61 individuals of P. atra (Fig. 4a-b). In addition, there was a single event in which Colibri coruscans Gould 1846 sipped nectar from a single individual plant (Fig. 4c), and the passerine birds Cacicus chrysonotus d'Orbigny and Lafresnaye 1838 and Buthraupis montana d'Orbigny and Lafresnaye 1837 chewing the corolla of one plant each (Fig. 4d).

Reproductive phenology
We observed around 182 individuals along the three trails (Sillutinkara: 75±26, Chojllapata: 50±16, and Vacus: 57±20). The inflorescences of P. atra had an average of 19 ± 9.72 open flowers (CV: 87.64%; range: 1-35 flowers, n = 211) over its flowering period of three months, producing 7 flowers every 4 ± 1 days. The flowers of P. atra open early in the morning (≈ 6:00 am) and remain open for three days and nights. The different phenological states and durations were the following: bud set developed for about five months, from January to May; flowers were available from March to June, the implanted state lasted for six months (May to October), while the fruits lasted around nine months (August to April); and open fruits were recorded throughout the year (Fig. 3).
Nectar composition
Mean total nectar volume during anthesis was 61.78 ± 13.87 μL per inflorescence per day (n = 75 flowers from 15 individuals). Individuals produced 8.09 ± 1.59 μL of nectar per flower. According to the NMR results, it was determined that the major compounds in the nectar and their respective concentrations were the following: 3% β-fructofuranose, 18% β -fructopyranose, 11% β-glucopyranose and 6% α-glucopyranose, as well as traces of sucrose and 62% of water.
Floral visitors
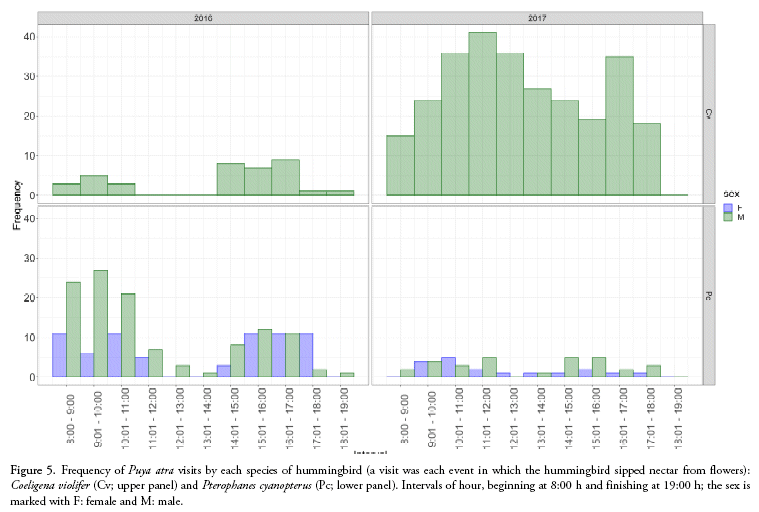
The frequency of activity for the two identified floral visitors of P. atra during the two years of sampling showed that C. violifer had a higher activity during the second year: the highest peak of activity changed from one year to another (Fig. 5). For P. cyanopterus, the levels of activity were different between males and females, and there was a higher activity in general during the first year. Males were more active in the morning hours at 8:00-11:00 h, especially during the first year. Females showed roughly the same frequency of visits throughout the day (Fig. 5) with slightly more activity at 8:00-11:00 h and 15:01-18:00 h.
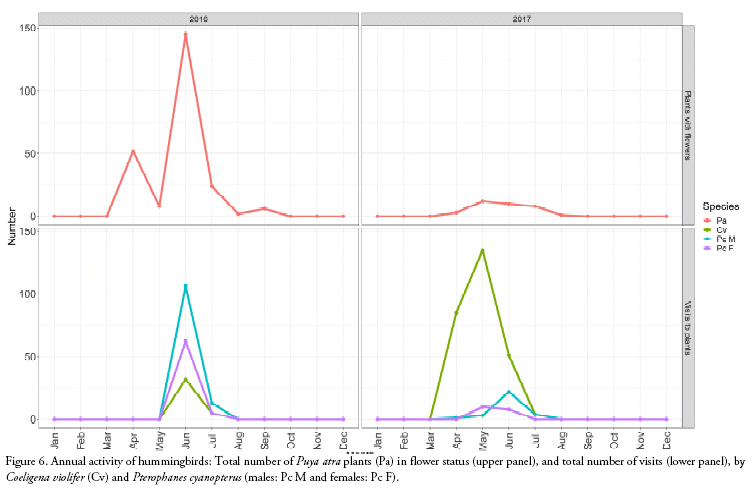
During the two years, the main flowering period was around June, but the numbers of inflorescences were highly variable in these two years (Fig. 3). However, during both years the plants received the same visitors, although in the first year the principal visitors were males of P. cyanopterus and in the second year males of C. violifer (Fig. 6).
Discussion
Reproductive phenology
Our results suggest that the reproductive cycle of P. atra is annual; the flowering period occurs once a year, similar to most other species of Bromeliaceae (Benzing 2000, Machado & Semir 2006, Wendt et al. 2008). During the two years of study P. atra had a well-delimited period of flowering from March to June, which is consistent with the dry season in the Yungas area (April to September). During this period, other plant species with flowers are scarce in the same area, which makes P. atra an important food resource for hummingbirds. This close relationship has also been reported in other studies on Puya species at high elevations in Peru (Salinas et al. 2007, Hornung-Leoni et al. 2013).
Nectar composition
Within the nectar composition, we detected sugars of fructose and glucose in their different cyclic forms. These sugars, along with sucrose, have been reported for other species of Puya, although their respective proportions changed according to the group of visitors (i.e. high sucrose content in the nectars of hummingbird flowers vs. low sucrose content in perching bird flowers) (Scogin & Freeman 1984, Baker et al. 1998). A comprehensive study of nectar composition in different genera of Bromeliaceae showed that the nectar always contained glucose, fructose and sucrose (Krömer et al. 2008), and according to the floral syndrome, nectars of bromeliad species pollinated by hummingbirds are generally sucrose-rich (Galetto & Bernardello 2003, Krömer et al. 2008, Gottlinger et al. 2019). However, the NMR, which detects cyclical structures of sugars more accurately, only detected trace amounts of sucrose and only found glucose and fructose in cyclical forms in the nectar of P. atra, which in our study area is only visited by hummingbirds. Thus, this species of Bromeliaceae is not sucrose-rich, but shows a predominance of hexoses as other Puya species pollinated by passerine birds (Scogin & Freeman 1984, Baker et al. 1998). Nevertheless, hummingbirds are able to digest fructose and glucose equally and use them as a source of energy (Chen & Welch 2014).
Floral visitors
The results obtained in this study showed that P. atra is a species visited almost exclusively by two species of long-billed hummingbirds, consistent with its flower morphology (tubular corolla), an thesis (opens early in the morning), phenology and nectar characteristics (diluted with 62% of water and 38% of sugars). This is similar to other studies on mainly hummingbird-visited Puya species in the Colombian and Peruvian Andes, such as P. rauhii L.B. Sm., P. alpestris Poepp. Gay, P. coerulea Miers, P. raimondii Harms, P. venusta Phil., P. nitida Mez. and P. trianae Baker (Hornung-Leoni & Sosa 2005, Salinas et al. 2007, Hornung-Leoni et al. 2013, Restrepo-Chica & Bonilla-Gómez 2017), supporting a co-adaptation between species of both families, Bromeliaceae and Trochilidae. However, passerine birds from different families (Emberizidae, Thraupidae, Mimidae, Tyrannidae and Icteridae) have also been observed visiting some of these Puya species, whereas bat-pollination has been recorded for two other Puya species (Aguilar-Rodríguez et al. 2019). The passerine birds C. chrysonotus and B. montana that visited P. atra chewed the plant's corolla, which has also been reported for Zonotrichia capensis Statius Müller 1776, Geospizopsis plebejus Tschudi 1844, G. unicolor d'Orbigny and Lafresnaye 1837, and Phrygilus punensis Ridway 1887 in P. raimondii (Salinas et al. 2007).
The flowers of P. atra remain open throughout three days and nights, when the reproductive organs of the plant remain accesible to visitors (anthesis). However, we did not observe other diurnal or nocturnal visitors such as insects or bats, only hummingbirds. This may be explained by the high elevation of 3,500 m in which the species grows, where the activity of bats and insects is very low, especially at night and early in the morning due to the low temperatures (Kessler & Krömer 2000, Krömer et al. 2006). A probable reason for the predominance of hummingbirds as pollinators in cold and humid montane habitats is that they are the best-adapted major biotic pollinator group under these conditions, whereas insects and bats are physiologically limited (Givnish et al. 2014, Kessler et al. 2020). As endothermic organisms, hummingbirds can function even at low temperatures, and their ability to fall into torpor at night is highly energy-efficient (Proctor et al. 1996, Abrahamczyk & Kessler 2015).
Pterophanes cyanopterus seems to be the main visitor of P. atra, whose plants are synchronising their flowering state with the visits of this hummingbird species. When the number of plants with flowers was high in the first year, the number of visits by P. cyanopterus was also high, and in the second year when the abundance of flowering plants decreased the number of visits was low as well. Although P. atra showed a high inter-annual variation, it seems that there is a specialisation with this hummingbird even though long-bill hummingbirds have been considered unspecialised (Cotton 1998) and it has been suggested that hummingbirds at high elevations can be flexible consumers and less dependent on particular sources of nectar compared to low elevation hummingbirds (Woods & Ramsay 2001). This contrasts to what was found in P. raimondii where the inter-annual variation does not permit a specialisation with its visitors (Salinas et al. 2007).
Pterophanes cyanopterus showed greater activity in the morning during the first year, whereas during the second year, it had a more constant activity throughout the day. This may be a result of the possible synchronisation between the plant and hummingbird mutualism. It is possible that P. cyanopterus may have learned about the plant reward availability; and the plant, in turn, might replenish its nectar after visitor removal as has been shown for other bromeliad species (Nepi et al. 2001, Ordano & Ornelas 2004). Whereas C. violifer showed greater activity during the afternoon in the first year, and in the second year it showed activity throughout the day. This is in contrast to the results obtained for P. hamata L.B. Smith where the highest activity for feeding of hummingbirds was between 14:00-15:00 h (Woods & Ramsay 2001).
Conclusions
This study showed that P. cyanopterus and C. violifer are the primary visitors of P. atra, which is an important resource of nectar since there are not many other plants in bloom during the dry season in the montane cloud forest. Puya atra had a well-delimited period of flowering from March to June, which is consistent with the dry season in the Yungas area. Additionally, it seems that P. cyanopterus has a synchronisation with the plant and that it is more sensitive to temporal and spatial resource variation at high-altitudes where climatic conditions become difficult (Woods & Ramsay 2012). Pterophanes cyanopterus is not only the main floral visitor of P. atra but also probably its main pollinator, as its morphology benefits the interaction between them. By having a larger body and bill size, this hummingbird usually made contact with the stigma and stamens when visiting the plant, which allows for transporting the pollen on the forehead. However, future studies about the breeding system and nectar production throughout the day would provide important information to better understand the dependence of cloud forest bromeliads to pollinators.
Acknowledgments
We would like to thank the Fondos de investigacion Erika Geyger for financing the research work. To Optics for the Tropics and Idea Wild for their support with equipment. To ANMI Cotapata for allowing us to enter the study area and DGBAP for the permissions. To Christian Ramos for help with graphics and suggestions. To Yonny Flores for processing the nectar samples. To Mariana Villegas, Isabel Gomez, Daniel Larrea, Sissi Lozada, Luis Pacheco and Leila Porter for commenting on the manuscript.
References
Abrahamczyk, S. & M. Kessler. 2015. Morphological and behavioural adaptations to feed on nectar: how feeding ecology determines the diversity and composition of hummingbird assemblages. Journal of Ornithology 156(2): 333-347.
Aguilar-Rodríguez, P.A., T. Krömer, M. Tschapka, J.G. García-Franco & M.C. MacSwiney. 2019. Bat pollination in Bromeliaceae. Plant Ecology & Diversity 12(1): 1–19.
Baker, H.G., I. Baker & S.A. Hodges. 1998. Sugar composition of nectars and fruits consumed by birds and bats in the tropics and subtropics. Biotropica 30(4): 559-586.
Barrett, S.C. & L.D. Harder. 1996. Ecology and evolution of plant mating. Trends in Ecology & Evolution 11(2): 73-79.
Bascompte, J. & P. Jordano. 2007. Plant-animal mutualistic networks: the architecture of biodiversity. Annual Review of Ecology, Evolution and Systematic 38: 567-593.
Benzing, D. H. 2000. Bromeliaceae: profile of an adaptive radiation. Cambridge University Press. [ Links ]
Buzato, S., M. Sazima & I. Sazima. 2000. Hummingbirdpollinated floras at three Atlantic forest sites. Biotropica 32(4b): 824-841.
Canela, M.B.F. & M. Sazima. 2005. The pollination of Bromelia antiacantha (Bromeliaceae) in southeastern Brazil: ornithophilous versus melittophilous features. Plant Biology 7(04): 411-416.
Chen, C.C.W. & K.C. Welch Jr. 2014. Hummingbirds can fuel expensive hovering flight completely with either exogenous glucose or fructose. Functional Ecology 28(3): 589-600.
Cotton, P.A. 1998. Coevolution in an Amazonian hummingbird‐plant community. Ibis 140(4): 639-646. [ Links ]
Dafni, A. 1992. Pollination ecology: a practical approach. Oxford University Press, Oxford. [ Links ]
Faegri, K. & L. Van der Pijl. 1979. Principles of pollination ecology. Pergamon Press, Toronto.
Elmendorf, S.C., K.D. Jones, B.I. Cook, J.M. Diez, C.A. Enquist, R.A. Hufft & M.D. Schwartz. 2016. The plant phenology monitoring design for the National Ecological Observatory Network. Ecosphere 7(4): e01303.
Galetto, L. & G. Bernardello. 2003. Nectar sugar composition in angiosperms from Chaco and Patagonia (Argentina): an animal visitor's matter? Plant Systematics and Evolution 238(1-4): 69-86.
García, M.T.A. & P.S. Hoc. 1998. Biología floral de Passiflora foetida (Passifloraceae). Revista de Biología Tropical 46(2): 191-202.
Givnish, T.J., M.H. Barfuss, B. Van Ee, R. Riina, K. Schulte, R. Horres, P.A. Gonsiska, R.S. Jabaily, M.C. Darren, J.A.C. Smith, K. Winter, G.K. Brown, M.E. Timothy, B.K. Holst, B.K., H.E. Luther, W. Till, G. Zizka, P.E. Berry & K.J. Sytsma. 2014. Adaptive radiation, correlated and contingent evolution, and net species diversification in Bromeliaceae. Molecular Phylogenetics and Evolution 71: 55-78.
Gonzalez, O. & B.A. Loiselle. 2016. Species interactions in an Andean bird–flowering plant network: phenology is more important than abundance or morphology. PeerJ 4, e2789.
Göttlinger, T., M. Schwerdtfeger, K. Tiedge & G. Lohaus. 2019. What do nectarivorous bats like? Nectar composition in Bromeliaceae with special emphasis on bat-pollinated species. Frontiers in Plant Science 10: 205.
Gouda, E.J., D. Butcher & C.S. Gouda. 2019. Encyclopaedia of Bromeliads, Version 4. http://bromeliad.nl/encyclopedia/ University Botanic Gardens, Utrecht (accessed: 22.04.2019).
Hornung-Leoni, C. & V. Sosa. .2005. Morphological variation in Puya (Bromeliaceae): an allometric study. Plant Systematics and Evolution 256(1-4): 35-53.
Hornung-Leoni, C.T., P.L. González-Gómez & A.J. Troncoso. 2013. Morphology, nectar characteristics and avian pollinators in five Andean Puya species (Bromeliaceae). Acta Oecologica 51: 54-61.
Ibisch, P.L. 1998. Bolivia is a megadiversity country and a developing country. pp. 213-241. In: Barthlott, W. & M Winiger (eds.) Biodiversity – A Challenge for Development Research and Policy. Springer, Heidelberg.
Jordano, P. 1987. Patterns of mutualistic interactions in pollination and seed dispersal: connectance, dependence asymmetries, and coevolution. The American Naturalist 129(5): 657-677. [ Links ]
Kessler, M. & T. Krömer. 2000. Patterns and ecological correlates of pollination modes among bromeliad communities of Andean forests in Bolivia. Plant Biology 2(6): 659-669.
Kessler, M., S. Abrahamczyk & T. Krömer. 2020. The role of hummingbirds in the evolution and diversification of bromeliads: unsupported claims and untested hypotheses. Botanical Journal of the Linnean Society 192(4): 592-608.
Krömer, T., M. Kessler, B.K. Holst., H.E. Luther, E.J. Gouda, P.L. Ibisch, W. Till & R. Vásquez. 1999. Checklist of Bolivian Bromeliaceae with notes on species distribution and levels of endemism. Selbyana 201-223.
Krömer, T. 2000. Distribution of terrestrial bromeliads along the La Paz to Caranavi road in Bolivia. Journal of the Bromeliad Society 50: 158-164. [ Links ]
Krömer, T., M. Kessler & S.K. Herzog. 2006. Distribution and flowering ecology of bromeliads along two climatically contrasting elevational transects in the Bolivian Andes. Biotropica 38(2): 183-195.
Krömer, T., M. Kessler, G. Lohaus & A.N. Schmidt‐Lebuhn. 2008. Nectar sugar composition and concentration in relation to pollination syndromes in Bromeliaceae. Plant Biology 10(4): 502-511.
Krömer, T., P.L. Ibisch, R. Vásquez, M. Kessler, B. Holst & H.E. Luther. 2014. Bromeliaceae. Pp. 418-439. In: Jørgensen, P. M., Nee, M. H., S. G. Beck, Arrázola, S. & M. Saldias (eds.) Catálogo de las Plantas Vasculares de Bolivia. Missouri Botanical Garden Press, St. Louis.
Machado, C.G. & J. Semir. 2006. Fenologia da floração e biologia floral de bromeliáceas ornitófilas de uma área da Mata Atlântica do Sudeste brasileiro. Brazilian Journal of Botany 29(1): 1163-174.
Meneses, R.I., S.G. Beck, E. García & H. Aranibar. 2004. Análisis del estado actual del conocimiento sobre la flora y vegetación del PN-ANMI Cotapata. Herbario Nacional de Bolivia-Programa BIAP-SERNAP, La Paz.
Nepi, M., M. Guarnieri & E. Pacini. 2001. Nectar secretion, reabsorption, and sugar composition in male and female flowers of Cucurbita pepo. International Journal of Plant Sciences 162(2): 353-358.
Ordano, M. & J.F. Ornelas. 2004. Generous-like flowers: nectar production in two epiphytic bromeliads and a meta-analysis of removal effects. Oecologia 140(3): 495-505.
Ortiz-Pulido, R., S.A. Díaz. O.I. Valle-Díaz & A.D. Fisher. 2012. Hummingbirds and the plants they visit in the Tehuacán-Cuicatlán Biosphere Reserve, Mexico. Revista Mexicana de Biodiversidad 83(1): 152-163.
Petanidou, T., A.N. Van Laere, W. Ellis & E. Smets. 2006. What shapes amino acid and sugar composition in Mediterranean floral nectars? Oikos 115(1): 155-169.
Proctor, M., P. Yeo & A. Lack. 1996. The natural history of pollination. Timber Press: Portland, Oregon.
Restrepo-Chica, M. & M.A. Bonilla-Gómez. 2017. Dinámica de la fenología y visitantes florales de dos bromelias terrestres de un páramo de Colombia. Revista Mexicana de Biodiversidad 88(3): 636-645.
Ribera, M.O. 1995. Aspectos ecológicos, del uso de la tierra y conservación en el Parque Nacional y Área Natural de Manejo Integrado Cotapata. pp. 1-84. In: Morales, C. (ed.) Caminos de Cotapata, Artes Gráficas Latina, La Paz.
Salinas, L., C. Arana & M. Suni. 2007. El néctar de especies de Puya como recurso para picaflores Altoandinos de Ancash, Perú. Revista Peruana de Biología 14(1): 129-134.
Schuchmann, K.L. 1999. Family Trochilidae (hummingbirds). Handbook of the birds of the world 5: 468-680. [ Links ]
Scogin, R. & C.E. Freeman. 1984. Floral pigments and nectar constituents in the genus Puya (Bromeliaceae). Aliso: A Journal of Systematic and Evolutionary Botany 10(4): 617-619.
Smith, L.B. & R.J. Downs. 1974. Pitcairnioideae (Bromeliaceae). Flora Neotropica. Monograph No. 14, Part 1. Hafner Press, New York.
Snow, D.W. 1981. Coevolution of birds and plants. Pp. 169-178. In: Forey, P.L. (ed.) The Evolving Biosphere. Part II. Coexistence and Coevolution, Cambridge University Press, Cambridge.
Stiles, F.G. & C.E. Freeman. 1993. Patterns in floral nectar characteristics of some bird-visited plant species from Costa Rica. Biotropica 25:191-205.
Thompson, J.N. 2005. The geographic mosaic of coevolution. The University of Chicago Press, Chicago. [ Links ]
Varadarajan, G.S. & G.K. Brown. 1988. Morphological variation of some floral features of the subfamily Pitcairnioideae (Bromeliaceae) and their significance in pollination biology. Botanical Gazette 149(1): 82-91.
Vázquez, D.P., W.F. Morris & P. Jordano. 2005. Interaction frequency as a surrogate for the total effect of animal mutualists on plants. Ecology Letters (10): 1088-1094.
Wendt, T., T.S. Coser, G. Matallana & F.A.G. Guilherme. 2008. An apparent lack of prezygotic reproductive isolation among 42 sympatric species of Bromeliaceae in southeastern Brazil. Plant Systematics and Evolution 275(1-2): 31-41.
Williams, R.J., B.A. Myers, D. Eamus & G.A. Duff. 1999. Reproductive phenology of woody species in a North Australian tropical savanna. Biotropica 31(4): 626-636.
Woods, S. & P.M. Ramsay. 2001. Variability in nectar supply: implications for high-altitude hummingbirds. Pp. 209-217. In: Ramsey, P.M. (ed.) The Ecology of Volcán Chiles: High-Altitude Ecosystems on the Ecuador-Colombia Border, Pebble and Shell, Plymouth.

