











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkEcología en Bolivia
versión impresa ISSN 1605-2528versión On-line ISSN 2075-5023
Ecología en Bolivia vol.55 no.1 La Paz abr. 2020
Artículo
Variación de la amplitud del nicho isotópico de tres especies de peces en un gradiente de disponibilidad de recursos
Isotopic niche width variation of three fish species in a resource availability gradient
Aldo Echeverría1, Marc Pouilly2 & Danny Rejas1*
1Unidad de Limnología y Recursos Acuáticos, Universidad Mayor de San Simón, Av. Ballivián 591, Cochabamba, Bolivia.
2Institut de Recherche pour le Développement UMR BOREA, Museum National d'Histoire Naturelle, 61 rue Buffon F75006 Paris, Francia.
*Autor de correspondencia: danny.rejas@gmail.com
Recibido: 21.08.19, Aceptado: 05.12.19.
Resumen
Con el objetivo de estudiar el efecto de la disponibilidad de recursos sobre la amplitud del nicho trófico en peces de la Amazonia boliviana, comparamos los nichos isotópicos (como sustituto del nicho trófico) de tres especies de peces (Acestrorhynchus altus, Hoplias malabaricus y Pygocentrus nattereri) en un gradiente de disponibilidad de presas. Tomamos muestras de peces en lagos localizados en la llanura de inundación de los ríos Blanco (baja disponibilidad de presas), Iténez (disponibilidad intermedia) y San Martín (disponibilidad alta), analizamos los ratios de isótopos estables (δ13C and δ15N) y estimamos las elipses estándar y amplitud de nicho isotópico (área) empleando el paquete SIBER en R. Acestrorhynchus altus se comportó como un forrajero óptimo, mostrando un nicho isotópico más amplio a menor disponibilidad de presas y un nicho más estrecho en los sitios con disponibilidad intermedia y alta. Pygocentrus nattereri mostró un nicho más amplio a densidades de presa intermedias, y no encontramos efecto de la disponibilidad de presas sobre la amplitud del nicho trófico de H. malabaricus. Esto indica que, para estas dos especies, factores tales como la selectividad de presas, transparencia del agua, tamaño de la boca y hábitos alimenticios, podrían ser más importantes que la disponibilidad de presas para determinar la amplitud del nicho trófico.
Palabras clave : Amazonia, Bolivia, Ecología trófica, Isótopos estables, SIBER.
Abstract
To study the effect of resource availability on the trophic niche width of fishes in the Bolivian Amazon basin, we compared isotopic niche widths (as a surrogate for trophic niche) of three fish species (Acestrorhynchus altus, Hoplias malabaricus and Pygocentrus nattereri) in a gradient of prey availability. We sampled fishes in lakes located in the floodplain of the rivers Blanco (low prey availability), Iténez (intermediate prey availability) and San Martín (high prey availability), analyzed them for ratios of stable isotopes (δ13C and δ15N), and estimated the standard ellipse and isotopic niche width using the SIBER package in R. Acestrorhynchus altus behaved as an optimal forager, showing wider isotopic niche where prey were less abundant, and less wide isotopic niche in rivers with intermedíate and high prey availability. P. nattereri showed wider trophic niche at intermedíate prey densities and H. malabaricus showed no effect of prey availability on isotopic niche width. This indicates that, for these species, factors such as prey selectivity, water transparency, mouth size or feeding habits, may be more important than prey availability in determining trophic niche width.
Key words: Amazonia, Bolivia, SIBER, Stable isotopes, Trophic ecology.
Introducción
El nicho ecológico es un concepto fundamental en ecología (Newsome et al. 2007), actualmente definido como un hipervolumen n-dimensional que puede ser representado por componentes biológicos y ambientales (Hutchinson 1957). El nicho trófico es uno de los principales componentes del nicho ecológico, pues ofrece información primaria sobre el uso de recursos (presas) y puede ser relacionado con variables ambientales (hábitat). A menudo, se evalúa la amplitud del nicho trófico estudiando la diversidad de la dieta en el contenido gástrico (Hyslop 1980). Sin embargo, este método presenta limitaciones tales como la dificultad pata medir con precisión la abundancia relativa de los distintos ítems, la falta de integración temporal de la información, y la imposibilidad de incorporar la variabilidad en las tasas de asimilación de las presas (Bearhop et al. 2004, Inger et al. 2006, Robb et al. 2012).
El uso de la varianza de las señales isotópicas de los tejidos de los consumidores fue propuesto por Bearhop et al. (2004) como alternativa para medir la amplitud del nicho trófico, superando las limitaciones anteriormente mencionadas. Los valores de δ13C proporcionan información sobre las fuentes de carbono utilizadas, mientras que los valores de δ15N indican el nivel trófico en el que se encuentra dicha especie (Fry 2006, Grey & Jackson 2012). Asimismo, ambas variables pueden ser utilizadas como los ejes principales dentro del nicho ecológico de Hutchinson (Jackson et al. 2011), representando los componentes biológicos y ambientales en un espacio bidimensional denominado "nicho isotópico" (Newsome et al 2007).
Varios estudios han tratado de determinar el efecto de la disponibilidad de recursos sobre la amplitud del nicho isotópico, mostrando que estas variables pueden estar directa (Brush et al. 2014) o inversamente relacionadas (Young & Ferguson 2014), pero también que la amplitud del nicho puede ser independiente de la disponibilidad de recursos (Jayasinghe et al. 2014). Una relación directa (positiva) entre disponibilidad de recursos y amplitud del nicho puede ser explicada por la teoría clásica de la competencia, que establece que la coexistencia entre especies es sobrellevada a través de la diferenciación de nichos (o segregación), que reduce la competencia (Pianka 1974, Pianka 1976). La teoría clásica de la competencia predice que la amplitud del nicho será menor cuando los recursos sean escasos y mayor cuando los recursos sean abundantes. Por otro lado, una relación inversa concuerda con la teoría del forrajeo óptimo (MacArthur & Pianka 1966, Townsend & Winfield 1985, Rooney et al. 2006), que sugiere que los animales son capaces de elegir sus recursos con el fin de maximizar la ganancia neta de energía en el menor tiempo posible. En este sentido, la teoría del forrajeo óptimo predice que los organismos poseerán nichos isotópicos más estrechos en condiciones favorables, debido a un proceso de especialización relacionado a la gran oferta alimenticia disponible. Cuando la disponibilidad de recursos disminuye, la amplitud de nicho incrementa debido a que el forrajeador se ve obligado a usar la mayor variedad de recursos para sobrevivir. Por último, la ausencia de relación entre la disponibilidad de los recursos y la amplitud del nicho responde a la teoría de la inflexibilidad (Connor & Simberloff 1983, Correa & Winemiller 2014), que sugiere que las variaciones de la amplitud del nicho no pueden ser explicadas sólo por la disponibilidad de recursos, pues existen otros factores externos que la determinan.
La parte alta de la cuenca amazónica presenta un amplio rango de variación de productividad primaria y secundaria. Al mismo tiempo, existen varias especies de peces con amplia distribución (Rejas & Maldonado 2000, Pouilly et al. 2004) que, como consecuencia, habitan en un amplio rango de variación de disponibilidad de recursos. Esto proporciona el marco ideal para determinar el efecto de la disponibilidad de recursos sobre la amplitud del nicho isotópico de tres especies de peces piscívoros de amplia distribución en las tierras bajas bolivianas: Acestrorhynchus altus, Hoplias malabaricus y Pygocentrus nattereri.
Partiendo de esa base conceptual, se plantearon tres hipótesis alternativas para explicar las variaciones de la amplitud del nicho isotópico de esas tres especies en tres sistemas acuáticos diferenciados por su disponibilidad de recursos: i) La amplitud del nicho será mayor en ambientes con mayor disponibilidad de presas (teoría de competencia clásica); ii) la amplitud de nicho se verá reducida en ambientes con mayor disponibilidad de presas (teoría del forrajeo óptimo) y iii) las variaciones en la amplitud del nicho serán independientes de la disponibilidad de recursos (teoría de la inflexibilidad).
Área de estudio
El estudio se llevó a cabo en los ríos Blanco (aguas blancas), Iténez (aguas claras) y San Martín (aguas mixtas), localizados en la cuenca del río Iténez (noreste de Bolivia) que abarca alrededor de 300.000 km2 en territorios de Brasil y Bolivia, con altitudes que varían entre 126-200 m y una temperatura media de 27°C. La precipitación anual es de 1.400 mm con períodos de lluvias (diciembre a mayo) y seco (junio a noviembre) claramente diferenciados (Pouilly et al. 2012). Los peces se capturaron en dos lagunas de la llanura de inundación de cada río: Zacarías (13°16'S-63°43'W) y La Granja (13°16'S-63°43'W) en el río Blanco; Curichá (12°37'S-63°26'W) y Negra (12°38'S-63°25'W) en el río Iténez; Cambarazal (13°18'S-63°37'W) y Redonda (13°18'S-63°33'W) en el río San Martín (Fig. 1). El río Blanco discurre a través de la llanura aluvial por lo que acumula y transporta gran carga de material sedimentario. Presenta aguas turbias con gran cantidad de material en suspensión, característico de ríos de orígenes andinos de aguas blancas (Maldonado & Goitia 2011). Los ríos Iténez y San Martín drenan principalmente zonas del Escudo Brasileño, compuestas por rocas metamórficas de lenta erosión; tienen poca carga sedimentaria, y concentraciones menores de nutrientes respecto al río Blanco. Sin embargo, la productividad primaria en ambos ríos es mayor respecto a la del río Blanco debido a su mayor transparencia (López Siangas et al. 2012). Pouilly & Camacho (2011) realizaron capturas con una batería de redes igual a la descrita más adelante, mostrando que existe un gradiente de disponibilidad de recursos para los peces piscívoros en estos tres ríos (presas: peces con longitud estándar ≤20 cm).
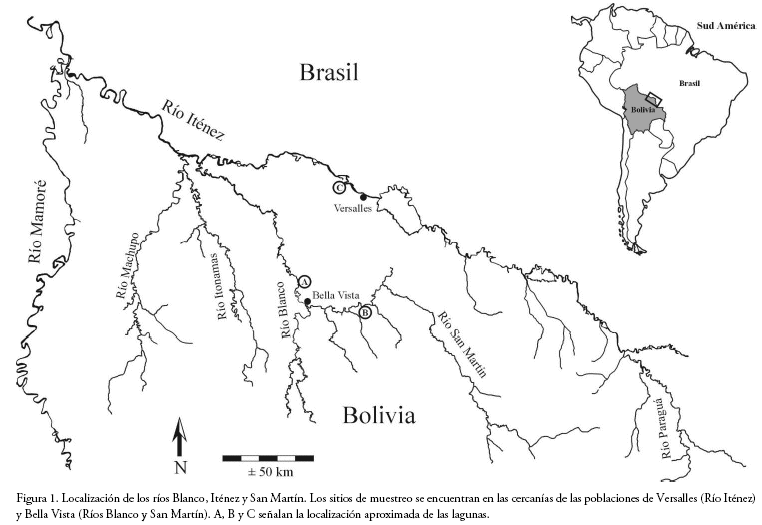
El río Blanco presenta la menor disponibilidad de presas (X= 528 CPUE), el río Iténez muestra disponibilidad intermedia (867 CPUE) y la mayor disponibilidad de presas se observa en el río San Martín (1.249 CPUE).
Métodos
El material fue colectado durante la temporada seca, en tres muestreos en el río Iténez (junio, septiembre y noviembre), y en seis muestreos en los ríos Blanco y San Martín (mensualmente, de junio a noviembre). Todas las lagunas se encuentran cerca al canal principal del río (entre 50 a 200 m), se conectan al río durante el período de inundación, pero se encuentran aisladas durante el período seco. El diseño de muestreo asegura que los peces capturados habitaron las lagunas al menos dos meses, tiempo superior al tiempo medio de vida de los isótopos en el tejido muscular (Tieszen et al. 1983).
Las capturas se realizaron en el marco de un estudio más amplio que incluyó varias otras especies de peces, por lo cual se utilizó una batería de redes agalleras (2.5 m de altura x 25 m de largo) con 20, 25, 30, 35, 40 y 50 mm entre nudos (Pouilly & Camacho 2011). Los peces fueron identificados y se midió la longitud estándar (LE); muestras de 4-5 g de tejido muscular fueron tomadas de la región dorsal, colocadas individualmente en tubos de plástico y congeladas. En laboratorio las muestras fueron liofilizadas, pulverizadas y envasadas en cápsulas de estaño. Los análisis de δ13C y δ15N fueron realizados en el laboratorio de la UC Davis (Stable Isotope Facility laboratory, Davis, California). Para este estudio empleamos muestras de especímenes adultos. El criterio utilizado para la elección del tamaño fue la talla de madurez sexual. Por tanto, analizamos muestras de individuos de A. altus LE ≥ 149 mm (Dieb-Magalhães et al. 2015), H. malabaricus LE ≥ 139 mm (Marques et al. 2007) y Pygocentrus nattereri LE ≥ 140 mm (Duponchelle et al. 2007).
Los ratios de los isótopos fueron expresados en partes por mil (%o) en relación con estándares internacionales: PD Belemnita (PDB) para el carbono y nitrógeno atmosférico para el nitrógeno. Los ratios isotópicos se definen como:
![]()
Donde, R = 15N/14N o 13C/12C (Fry 2006).
Puesto que la amplitud del nicho isotópico puede ser afectada por el tamaño de los peces, para cada especie comparamos las medias de longitud estándar de las poblaciones de los tres ríos mediante ANOVAs. Empleamos análisis de regresión lineal para explorar la relación entre el tamaño de los peces (LE) y las señales de 13Cy15N.
Para comparar la amplitud de nicho de cada par de poblaciones, se utilizó la función de estimado posterior del paquete SIBER (Stable Isotope Bayesian Ellipses in R). El programa calcula primero la elipse estándar que describe los datos en el espacio δ13C- δ15N para cada población y el área de esta elipse (Standard Ellipse Area, SEA). Posteriormente, realizamos una corrección aritmética de la elipse (SEAc) para número pequeño de muestras (n<30). Finalmente, el programa calcula un "estimado posterior" que se emplea para construir una Elipse Posterior y sus parámetros derivados, específicamente el área (SEAb). El proceso produce un rango de valores probables para el parámetro calculado, reflejando la incertidumbre en el estimador. Estos valores estimados permiten la comparación de la amplitud de nicho de pares de poblaciones, determinando la probabilidad de que dos áreas de nicho sean diferentes mediante inferencia Bayesiana (Young et al. 2017). Todos los análisis estadísticos fueron realizados en el programa de software libre R 3.0.3 (R Core Team 2014).
Resultados
Analizamos un total de 306 muestras de peces: 73 de A. altus, 120 de H. malabaricus y 113 de P. nattereri. Para cada población, analizamos al menos 20 individuos, excepto para la población de A. altus del río Blanco, de la que solamente obtuvimos muestras de 12 individuos. El número de individuos, longitud estándar media y los ratios isotópicos de 13C y 15N de las poblaciones estudiadas se muestran en la tabla 1.
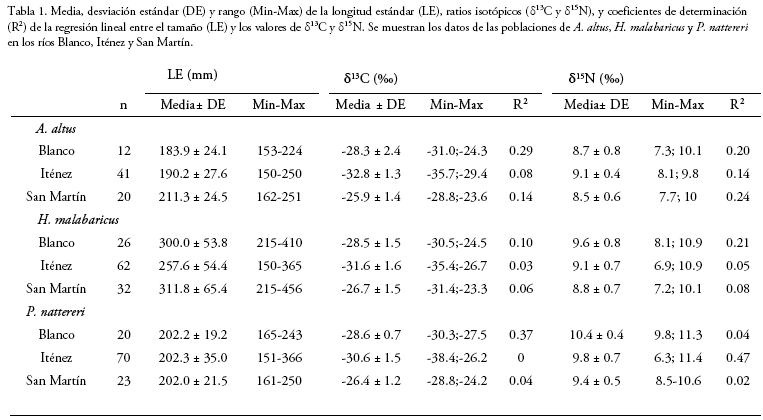
La media de LE de la población de A. altus del río San Martín fue significativamente mayor que las medias de LE de las poblaciones de esta especie en los ríos Blanco e Iténez (ANOVA, p < 0.01, Tukey, valores de p < 0.05; Fig. 2a). No existieron diferencias significativas entre estas dos últimas poblaciones (Tukey, p=0.74). La media de LE de la población de H. malabaricus del río Iténez fue significativamente menor respecto a las medias de LE de las poblaciones de esta especie en los ríos Blanco y San Martín (ANOVA, p < 0.001; Tukey, valores de p < 0.01), no existiendo diferencias significativas entre las poblaciones de estos últimos dos ríos (Tukey, p=0.71; Fig. 2b). Por último, no encontramos diferencias significativas entre las medias de LE de las poblaciones de P. nattereri de los tres ríos estudiados (ANOVA, p= 0.99; Fig. 2c).
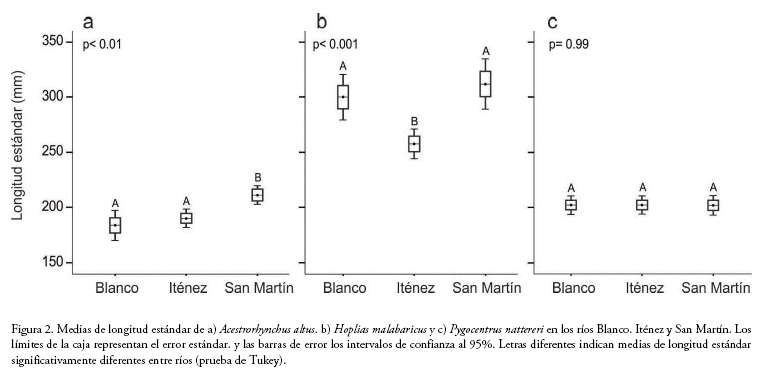
No encontramos relaciones significativas entre el tamaño de los peces (LE) y las señales isotópicas de 13C o 15N para ninguna de las poblaciones estudiadas en los tres ríos (R2 entre 0.04 y 0.47, p > 0.05, tabla 1).
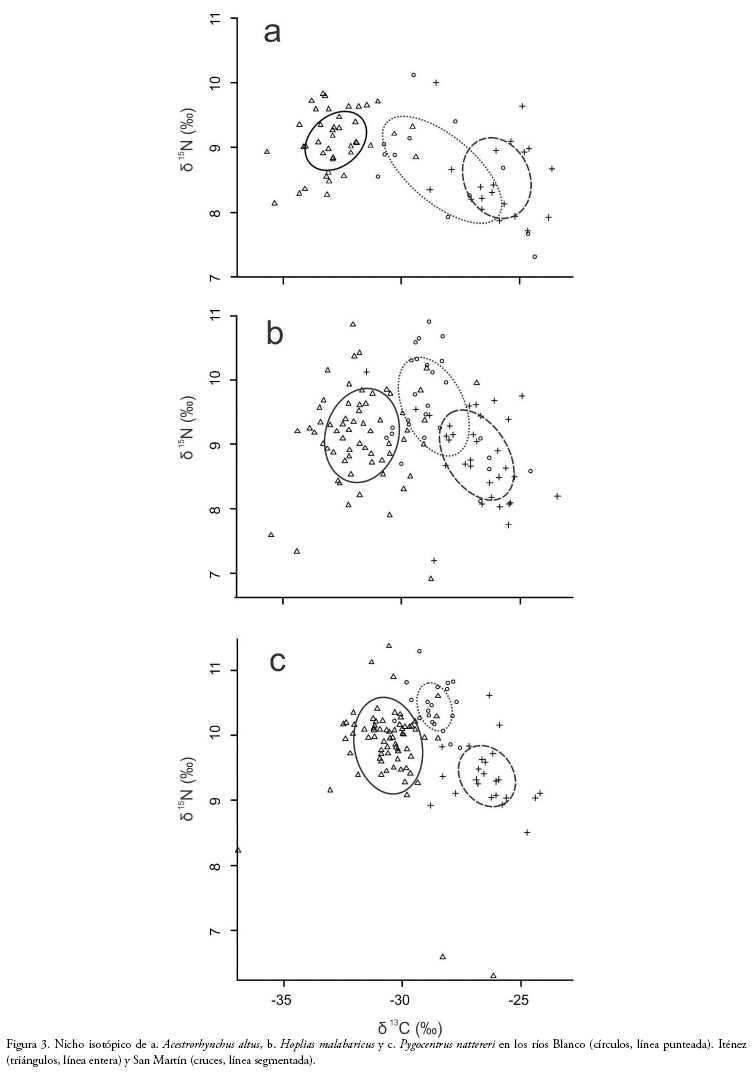
La amplitud del nicho isotópico (medida como SEAb) de la población de A. altus del río Blanco (4.99%o2) fue significativamente mayor que las de las poblaciones de esta especie en los ríos San Martín (2.77%o2; p=0.95) e Iténez (1.82 %o2; p=0.99) (Fig. 3). No existieron diferencias significativas entre las poblaciones de los ríos San Martín e Iténez (p=0.93). No encontramos diferencias significativas de amplitud de nicho isotópico (p< 0.95) entre las poblaciones de H. malabaricus de los ríos Blanco (3.36%o2), Iténez (3.61%o2) y San Martín (3.11%o2). La amplitud del nicho isotópico de P. nattereri fue significativamente mayor en la población del río Iténez (3.41%o2) que en las poblaciones de los ríos Blanco (0.87%o2; p = 1) y San Martín (1.69%o2; p = 1). No encontramos diferencias significativas entre las poblaciones de P. nattereri de los ríos Blanco y San Martín (p=0.82).
Discusión
Las señales isotópicas de 13C de las tres especies estudiadas mostraron valores menores en el río Iténez, intermedios en río Blanco y mayores en el San Martín. Las variaciones isotópicas de δ 13C se originan en productores primarios: en general las macrófitas acuáticas con vía fotosintética C4 son la fuente de carbono más enriquecida (alrededor de -13%o), el fitoplancton es la fuente de carbono más empobrecida (-33%o o menos) y las plantas terrestres y macrófitas con vía fotosintética C3 varían entre -25%o y -30%o (Manetta et al. 2003, Mortillaro et al. 2015, Rejas 2018). Los valores medios de δ13C encontrados en las poblaciones de los tres ríos estudiados, presentan una variación entre -32.8 y -25.9, lo que corresponde a una cadena trófica mayormente basada en productores primarios con vía fotosintética C3. La señal empobrecida de δ13C observada en el río Iténez podría deberse al uso de fuentes alternativas de carbono que tienen usualmente valores más empobrecidos de δ13C, tales como perifiton (Pouilly et al. 2013) o bacterias metanótrofas (Rejas 2018).
Por otro lado, los valores medios de δ15N de las tres especies (entre 8.5 y 10.4%o) son similares a los encontrados en otros estudios realizados en estas especies (Pouilly et al. 2013, Mortillaro et al. 2015, Carvalho et al. 2018) y se encuentran dentro del rango encontrado para especies piscívoras de la Amazonia (Rejas 2018). Los rangos de variación de δ15N fueron estrechos para las tres especies estudiadas, y están por debajo del rango de variación estimado para un nivel trófico (+2.80 ± 0.40%o para muestras de músculo sin tratamiento para extracción de lípidos; McCutchan et al. 2003), lo que confirma que las tres especies estudiadas son piscívoras especialistas.
Relación entre la longitud del pez y la amplitud del nicho isotópico
Debido a que las tasas de fraccionamiento del 13C a lo largo de la cadena trófica son relativamente pequeñas (+1.30 ± 0.30%o en cada nivel trófico, McCutchan et al. 2003), en general, a nivel poblacional no existe correlación entre el tamaño de los peces y los valores de δ13C a menos que exista un cambio drástico en la dieta a lo largo del ciclo de vida (Tieszen et al. 1983), pues individuos de distintos tamaños suelen alimentarse en base a presas que utilizan las mismas fuentes de carbono (Overman & Parrish 2001). Por otro lado, debido a su factor constante de fraccionamiento (Jepsen & Winemiller 2002), puede existir un efecto de la longitud de los peces sobre su señal de δ15N (Gu et al. 1996a, b,Beaudoin et al. 1999, Overman & Parrish 2001). En el presente trabajo, pese a que encontramos diferencias significativas de longitud estándar entre poblaciones de A. altus y H. malabaricus, estas diferencias no tuvieron efecto sobre las señales isotópicas de 13C y 15N y por tanto sobre la amplitud del nicho isotópico.
Disponibilidad de recursos y amplitud de nicho
La amplitud de nicho de A. altus fue mayor en el río con menor disponibilidad de presas (río Blanco) y disminuyó en los ríos con disponibilidad intermedia y alta. Este patrón nos lleva a la conclusión de que A. altus es un forrajeador óptimo, que elige las presas con mayor valor energético cuando ellas están disponibles. Como consecuencia tiende a especializarse y reducir su nicho trófico/isotópico cuando aumenta la cantidad de presas y tiende a ser más generalista/oportunista cuando la cantidad de presas disminuye. Estudios previos respaldan esta conclusión, pues mencionan que A. altus es un pez principalmente carnívoro, que en condiciones de escasez de recursos es capaz de consumir una gran cantidad de ítems alimenticios tales como escamas de peces, e invertebrados como camarones y moluscos (Rocha et al. 2011, Neuhaus et al. 2016). Por otro lado, Cantanhêde et al. (2008) encontraron que el nicho trófico de A. altus se redujo al aumentar la disponibilidad de presas. Luego de la formación del embalse el Manso (Brasil), las densidades del pequeño pez Moenkhausia dichroura incrementaron, convirtiéndose esta especie en la presa principal para A. altus.
Dentro del rango estudiado, la amplitud del nicho isotópico de H. malabaricus fue independiente de la disponibilidad de presas, es decir que se comporta de acuerdo a lo establecido en la teoría de la inflexibilidad. Ciertas características de esta especie, tales como sus hábitos sedentarios (De Almeida et al. 1997), selectividad por peces pelágicos (Characidae, Poecilidae) en su alimentación (Correa et al. 2012) y tamaño de presas limitado por el tamaño de su boca (Prejs 1987), la hacen una especie especializada. Estas características determinan que su amplitud de nicho isotópico sea limitada, y posiblemente las presas obtenidas por H. malabaricus no reflejen la disponibilidad total de presas presente en su medio ambiente. Sin embargo, no podemos descartar que en un gradiente más amplio de disponibilidad de presas, pueda existir un cambio en los hábitos alimenticios en esta especie. De hecho, nuestros resultados difieren de los registrados por Luz-Agostinho et al. (2008), quienes mencionan que tras la construcción de una represa, la disminución de hábitats, presas y refugios tuvo como efecto un cambio en la dieta, y un aumento en la amplitud del nicho trófico de H. malabaricus por la inclusión de varios ítems, como el camarón (Macrobrachium amazonicum) y algunas especies de Gymnotus. Los valores de SEAc encontrados en nuestro trabajo (3.11-3.61%o2) fueron menores que los registrados para otras especies del género Hoplias en la cuenca del rio Paraná (3.8-5.2%o2; (Philippsen et al. 2015). Lo que indica que H. malabaricus es especialista en comparación con otras especies del mismo género.
De la misma manera, aunque la amplitud de nicho isotópico de P. nattereri fue mayor en el río Iténez que en los ríos Blanco y San Martín, no encontramos un patrón respecto a la disponibilidad de recursos. Al igual que las dos anteriores, P. nattereri es una especie predominantemente piscívora, los peces representan más del 90% de su contenido gástrico (Novakowski et al. 2008, Ferreira et al. 2014) lo que sugiere un alto grado de especialización, que se verá reflejado en un nicho isotópico reducido (Sa-Oliveira et al. 2014). Varios estudios han encontrado que el nicho de P. nattereri es menor al de otros piscívoros (Novakowski et al. 2008, Ferreira et al. 2014), lo que coincide con nuestros resultados, particularmente en los ríos Blanco y San Martín.
El éxito de los peces piscívoros en la captura de sus presas estará influenciado por su dependencia del sentido de la vista (Rodríguez & Lewis Jr. 1997), transparencia del agua (Montenegro et al. 2013), selectividad de presas (Correa et al. 2012) y tamaño de la boca (Prejs 1987). Todos estos factores harán que la disponibilidad de presas para un individuo de una especie determinada, sea diferente de la disponibilidad estimada mediante determinado método de captura.
Conclusiones
En síntesis, hemos observado un efecto de la disponibilidad de presas sobre la amplitud del nicho isotópico en A. altus (forrajeo óptimo), mientras que no existió un efecto sobre los nichos isotópicos de H. malabaricus y P. nattereri (teoría de la inflexibilidad). Las tres especies presentan un nicho relativamente estrecho, confirmando resultados obtenidos por ecomorfología (Pouilly et al. 2003). Nuestros resultados sugieren que la amplitud del nicho isotópico de estas últimas dos especies depende de múltiples factores más complejos que simplemente la disponibilidad de presas.
Agradecimientos
Este trabajo fue financiado parcialmente por el IRD mediante el proyecto "Trophic Ecology of Amazonian Aquatic Systems" (JEAI-EMAA) y por ASDI mediante el proyecto C-26 "Impacto del pez invasor Paiche (Arapaima gigas) sobre peces nativos de la Amazonia boliviana".
Referencias
De Almeida, V.L.L., N.S. Hahn & A.E.A. de M. Vazzoler. 1997. Feeding patterns in five predatory fishes of the high Paraná River floodplain (PR, Brazil). Ecology of Freshwater Fish 6(3): 123-133.
Bearhop, S., C.E. Adams, S. Waldron, R.A. Fuller & H. Macleod. 2004. Determining trophic niche width: a novel approach using stable isotope analysis. Journal of Animal Ecology 73(5): 1007-1012.
Beaudoin, C.P., W.M. Tonn, E.E. Prepas & L.I. Wassenaar. 1999. Individual specialization and trophic adaptability of northern pike (Esox lucius): an isotope and dietary analysis. Oecologia 120(3):
386-396.
Brush, J.M., M. Power, K.D. Clarke & C.J. Pennell. 2014. The impact of low flow on riverine food webs in south-central Newfoundland. River Research and Applications 31(9): 1082-1092.
Cantanhêde, G., N.S. Hahn, R. Fugi & É.A. Gubiani. 2008. Alterations on piscivorous diet following change in abundance of prey after impoundment in a Neotropical river. Neotropical Ichthyology 6(4): 631-636.
Carvalho, F., M. Power, B.R. Forsberg, L. Castello, E.G. Martins & C.E.C. Freitas. 2018. Trophic Ecology of Arapaima sp. in a ria lake—river–floodplain transition zone of the Amazon. Ecology of Freshwater Fish 27(1): 237-246.
Connor, E.F. & D. Simberloff. 1983. Interspecific competition and species co-occurrence patterns on islands: null models and the evaluation of evidence. Oikos 41(3): 455-465.
Corrêa, F., M.C. Claudino, R.F. Bastos, S. Huckembeck & A.M. Garcia. 2012. Feeding ecology and prey preferences of a piscivorous fish in the Lagoa do Peixe National Park, a Biosphere Reserve in Southern Brazil. Environmental biology of fishes 93(1): 1-12.
Correa, S.B. & K.O. Winemiller. 2014. Niche partitioning among frugivorous fishes in response to fluctuating resources in the Amazonian floodplain forest. Ecology. 95(1): 210-224.
Dieb‐Magalhães, L., A.C. Florentino & M.G.M. Soares. 2015. Length–weight relationships and length at first maturity for nine fish species of floodplain lakes in Central Amazon (Amazon Basin, Brazil). Journal of
applied ichthyology 31(6): 1182-1184.
Duponchelle, F., F. Lino, N. Hubert, J. Panfili, J. Renno, E. Baras, J.-P. Torrico, R. Dugué & J. Nunez. 2007. Environment‐related life‐history trait variations of the red‐bellied piranha Pygocentrus nattereri in two river basins of the Bolivian Amazon. Journal of Fish Biology 71(4): 1113-1134.
Ferreira, F.S., W. Vicentin, F.E. Dos S. Costa & Y.R. Súarez. 2014. Trophic ecology of two piranha species, Pygocentrus nattereri and Serrasalmus marginatus (Characiformes, Characidae), in the floodplain of the Negro River, Pantanal. Acta Limnologica Brasiliensia 26(4): 381-391.
Fry, B. 2006. Stable isotope ecology. Springer, NuevaYork. [ Links ]
Grey, J. & M.C. Jackson. 2012. “Leaves and eats shoots”: direct terrestrial feeding can supplement invasive red swamp crayfish in times of need. PLoS One 7: e42575.
Gu, B., C.L. Schelske & M.V. Hoyer. 1996a. Intrapopulation Feeding Diversity in Blue Tilapia: Evidence from Stable-Isotope Analyses. Ecology 78(7): 2263-2266.
Gu, B., C.L. Schelske & M.V Hoyer. 1996b. Stable isotopes of carbon and nitrogen as indicators of diet and trophic structure of the fish community in a shallow hypereutrophic lake. Journal of fish biology 49(6): 1233-1243.
Hutchinson, G.E. 1957. Cold spring harbor symposium on quantitative biology. Concluding remarks 22: 415–427.
Hyslop, E.J. 1980. Stomach contents analysis—a review of methods and their application. Journal of fish biology 17(4): 411-429.
Inger, R., G.D. Ruxton, J. Newton, K. Colhoun, J.A. Robinson, A.L. Jackson & S. Bearhop. 2006. Temporal and intrapopulation variation in prey choice of wintering geese determined by stable isotope analysis. Journal of Animal Ecology 75(5): 1190-1200.
Jackson, A.L., R. Inger, A.C. Parnell & S. Bearhop. 2011. Comparing isotopic niche widths among and within communities: SIBER – Stable Isotope Bayesian Ellipses in R. Journal of Animal Ecology 80(3): 595-602.
Jayasinghe, U.A.D., G. Emili, Z. Li, W. Li, T. Zhang & J. Liu. 2015. Co-occurring bighead and silver carps show similar food preference but different isotopic niche overlap in different lakes. Environmental Biology of Fishes 98(4): 1185-1199.
Jepsen, D.B. & K.O. Winemiller. 2002. Structure of tropical river food webs revealed by stable isotope ratios. Oikos 96(1): 46-55.
López Siangas, E., M. Pouilly, A. Vallejos, T. Pérez & D. Rejas. 2012. Effect of water quality on growth of four fish species in the Iténez basin (Upper Madera, Amazon). Environmental Biology of Fishes 95(3): 371-381.
Luz-Agostinho, K.D.G., A.A. Agostinho, L.C. Gomes & H.F. Júlio. 2008. Influence of flood pulses on diet composition and trophic relationships among piscivorous fish in the upper Paraná River floodplain. Hydrobiologia 607(1): 187.
Macarthur, R.H. & E.R. Pianka. 1966. On optimal use of a patchy environment. The American Naturalist 100(916): 603-609.
Maldonado, M. & E. Goitia. 2011. La cuenca del río Iténez en Bolivia: descripción ecológica. Aguas del Iténez o Guaporé: recursos hidrobiológicos de un patrimonio binacional (Bolivia y Brasil). Edit. INIA, Cochabamba.
Manetta, G.I., E. Benedito-Cecilio & M. Martinelli. 2003. Carbon sources and trophic position of the main species of fishes of Baía River, Paraná River floodplain, Brazil. Brazilian Journal of Biology 63(2): 283-290.
Marques, C., A.S. Braun & N.F. Fontoura. 2007. Estimativa de tamanho de primeira maturação a partir de dados de IGS: Oligosarcus jenynsii, Oligosarcus robustus, Hoplias malabaricus, Cyphocharax voga, Astyanax fasciatus (Characiformes), Parapimelodus nigribarbis, Pimelodus maculatus, Trachelyopterus lucenai, Hoplosternum littorale, Loricariichthys anus (Siluriformes) e Pachyurus bonariensis (Perciformes) no Lago Guaíba e Laguna Dos Patos, RS. Biociências 15(2): 230-256.
McCutchan Jr, J.H., W.M. Lewis Jr., C. Kendall & C.C. Mc Grath. 2003. Variation in trophic shift for stable isotope ratios of carbon, nitrogen, and sulfur. Oikos 102(2): 378–390.
Montenegro, A.K.A., A.C.B. Vieira, M.M.L. Cardoso, J.E.R.T. De Souza & M.C. Crispim. 2013. Piscivory by Hoplias aff. malabaricus (Bloch, 1794): a question of prey availability?. Acta Limnologica Brasiliensia 25(1): 68-78.
Mortillaro, J.M., M. Pouilly, M. Wach, C.E.C. Freitas, G. Abril & T. Meziane. 2015. Trophic opportunism of central Amazon floodplain fish. Freshwater Biology 60(8): 1659-1670.
Neuhaus, E.B., D.A. Antonetti & U.H. Schulz. 2016. The invasive fish Acestrorhynchus pantaneiro Menezes, 1992 in Southern Brazil: Occurrence and food niche overlap with two native species. Journal of Applied Ichthyology 32(6): 1107-1112.
Newsome, S.D., C. Martinez Del Rio, S. Bearhop & D.L. Phillips. 2007. A niche for isotopic ecology. Frontiers in Ecology and the Environment 5(8): 429-436.
Novakowski, G.C., N.S. Hahn & R. Fugi. 2008. Diet seasonality and food overlap of the fish assemblage in a pantanal pond. Neotropical Ichthyology 6(4): 567-576.
Overman, N.C. & D.L. Parrish. 2001. Stable isotope composition of walleye: 15N accumulation with age and area-specific differences in δ13C. Canadian Journal of Fisheries and Aquatic Sciences 58(6): 1253-1260.
Philippsen, J.S., M. Hauser & E. Benedito. 2015. Isotopic niches of sympatric native and exotic fish species in a Neotropical floodplain. Anais da Academia Brasileira de Ciências 87(2): 825-833.
Pianka, E.R. 1974. Niche overlap and diffuse competition. Proceedings of the National Academy of Sciences 71(5): 2141-2145. [ Links ]
Pianka, E.R. 1976. Competition and niche theory. pp.114-141. En: May, R. (ed.) Theoretical ecology: principles and applications. WB Saunders, Philadelphia.
Pouilly, M. & J. Camacho. 2011. Composición de la comunidad de peces en la cuenca del río Iténez (Bolivia). pp 159–171. En: Van Damme P.A., M. Maldonado, M. Pouilly, and C. Doria (Eds.) Aguas del Iténez o Guaporé: recursos hidrobiológicos de un patrimonio binacional (Bolivia y Brasil). Editorial INIA, Cochabamba.
Pouilly, M., F. Lino, J.G. Bretenoux & C. Rosales. 2003. Dietary-morphological relationships in a fish assemblage of the Bolivian Amazonian floodplain. Journal of Fish Biology 62(5): 1137–1158.
Pouilly, M., T. Pérez, D. Rejas, F. Guzman, G. Crespo, J.- L. Duprey & J.R.D. Guimarães. 2012. Mercury bioaccumulation patterns in fish from the Iténez river basin, Bolivian Amazon. Ecotoxicology and Environmental Safety 83: 8-15.
Pouilly, M., D. Rejas, T. Pérez, J.L. Duprey, C.I. Molina, C. Hubas & J.R.D. Guimarães. 2013. Trophic structure and mercury biomagnification in tropical fish assemblages, Iténez River, Bolivia. PLoS ONE 8(5): e65054.
Pouilly, M., T. Yunoki, C. Rosales & L. Torres. 2004. Trophic structure of fish assemblages from Mamoré River floodplain lakes (Bolivia). Ecology of Freshwater Fish 13(4): 245–257.
Prejs, A. 1987. Risk of predation and feeding rate in tropical freshwater fishes: field evidence. Oecologia 72(2): 259-262. [ Links ]
R Core Team. 2014. R: A language and environment for statistical computing. http://www.R-project.Org [ Links ]
Rejas, D. 2018. Trophic structure of a floodplain fish assemblage in the upper Amazon basin, Bolivia. Revista de biologia tropical 66(3): 91-99 [ Links ]
Rejas, D. & M. Maldonado. 2000. Variaciones espaciotemporales de la composición de la comunidad de peces de una laguna de la várzea del río Ichilo (Cochabamba, Bolivia). Revista Boliviana de Ecología y Conservación 7: 37–46.
Robb, G.N., S. Woodborne & N.C. Bennett. 2012. Subterranean Sympatry: An Investigation into diet using stable isotope analysis. PLoS ONE 7(11): e48572.
Rocha Da, A.A.F., N.C.L. Dos Santos, G. De Araújo Pinto, T. Do Nascimento Medeiros & W. Severi. 2011. Diet composition and food overlap of Acestrorhynchus britskii and A. lacustris (Characiformes: Acestrorhynchidae) from Sobradinho reservoir, São Francisco river, Bahia State. Acta Scientiarum. Biological Sciences 33(4): 407–415.
Rodríguez, M.A. & W.M. Lewis Jr. 1997. Structure of fish assemblages along environmental gradients in floodplain lakes of the Orinoco River. Ecological monographs 67(1): 109–128.
Rooney, N., K. Mccann, G. Gellner & J.C. Moore. 2006. Structural asymmetry and the stability of diverse food webs. Nature 442 (7100): 265-269.
Sa-Oliveira, J.C., R. Angelini & V.J. Isaac-Nahum. 2014. Diet and niche breadth and overlap in fish communities within the area affected by an Amazonian reservoir (Amapá, Brazil). Anais da Academia Brasileira de Ciências 86(1): 383-406.
Tieszen, L.L., T.W. Boutton, K.G. Tesdahl & N.A. Slade. 1983. Fractionation and turnover of stable carbon isotopes in animal tissues: implications for δ13C analysis of diet. Oecologia 57(1-2): 32-37.
Townsend, C.R. & I.J. Winfield. 1985. The application of optimal foraging theory to feeding behaviour in fish. In: Fish energetics. Springer, Dordrecht.
Young, B.G. & S.H. Ferguson. 2014. Using stable isotopes to understand changes in ringed seal foraging ecology as a response to a warming environment. Marine Mammal Science 30(2): 706-725.
Young, H., K. Nigro, D.J. Mccauley, L.T. Ballance, E.M. Oleson & S. Baumann-Pickering. 2017. Limited trophic partitioning among sympatric delphinids off a tropical oceanic atoll. Plos One 12(8): e0181526.

