











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkEcología en Bolivia
versión impresa ISSN 1605-2528versión On-line ISSN 2075-5023
Ecología en Bolivia vol.54 no.2 La Paz set. 2019
Artículo
Abundancia de hongos gasteroides (Agaricomycetes, Basidiomycota) en el norte de la Amazonia boliviana
Abundance of gasteroid Fungi in the northern Bolivian Amazon
Diana Rocabado1* & Oswaldo Maillard1,2
1Herbario del Oriente Boliviano, Museo de Historia Natural Noel Kempff Mercado, Universidad Autónoma Gabriel René Moreno, Casilla Postal 2489, Avenida Irala 565, Santa Cruz, Bolivia. *Autora de correspondencia: diana.rocabado@gmail.com
2Dirección actual: Fundación para la Conservación del Bosque Chiquitano (FCBC). Av. Ibérica calle 6 Oeste 95, esq. Puerto Busch, Barrio Las Palmas. Santa Cruz, Bolivia.
Recibido: 04.04.19, Aceptado: 18.06.19.
Resumen
El presente estudio fue realizado en cuatro sitios de los bosques amazónicos en el norte del país, en los Departamentos de Pando (Las Ruinas, Candelaria) y Beni (Las Palmeras, 7 de Julio), con el objetivo principal de evaluar la abundancia de los hongos gasteroides y su relación con la precipitación, temperatura máxima, entrada de luz del dosel y la cobertura del sotobosque. El método de muestreo empleado consistió en colectas efectuadas en cuatro parcelas de 50 x 50 m, divididas a su vez en 50 subparcelas circulares de 5 m de diámetro. Los muestreos se desarrollaron en la época húmeda (noviembre-enero) y seca (julio-septiembre). Un total de 83 muestras de hongos gasteriodes fueron colectadas pertenecientes a los géneros Geastrum y Lycoperdon. Las abundancias de hongos por localidad mostraron una variación significativa (F = 8.78; P = 0.0002), donde la localidad de Las Ruinas presentó el mayor número de basidiomas colectados. Por otro lado, la abundancia de hongos fue significativamente diferente entre épocas de muestreo (F = 33.14, P = 0.0001), presentando mayor abundancia en la estación lluviosa. En cuanto a la entrada de luz del dosel, cobertura del sotobosque, precipitación y temperatura máxima, no se encontró que exista una relación con la abundancia de los hongos gasteroides.
Palabras clave: Ecología, Geastrum, Hongos neotropicales, Lycoperdon, Taxonomía de hongos.
Abstract
This study was carried out in four sites of the Amazonian forests in the north of the country, in the Departments of Pando (Las Ruinas, Candelaria) and Beni (Las Palmeras, 7 de Julio), with the main objective of evaluating the abundance of the gasteroid fungi and their relation with precipitation, maximum temperature, canopy light inlet and understory cover. The method of sampling used consisted of collections carried out in four plots of 50 x 50 m, divided in turn in 50 circular subplots of 5 m in diameter. Samplings were developed in the humid (November-January) and dry season (July-September). A total of 83 samples of gasteriod fungi were collected from the genera Geastrum and Lycoperdon. The abundances of fungi per locality showed a significant variation (F = 8.78; P = 0.0002), where the locality of Las Ruinas presented the largest number of individuals. On the other hand, the abundance of fungi was significantly different between sampling times (F = 33.14, P = 0.0001), presenting greater abundance in the rainy season. As for the entrance of canopy light, vegetation cover, precipitation and maximum temperature, it was not found that there is a relationship with the abundance of gasteroid fungi.
Keywords: Ecology, Fungal taxonomy, Geastrum, Lycoperdon, Neotropical fungi.
Introducción
Los hongos son un grupo hiperdiverso de organismos heterotróficos, y muy importantes por la función que cumplen en los ecosistemas tropicales y otros (Hawksworth & Colwell 1992), especialmente en el reciclado de nutrientes, siendo claves en su rol como descomponedores, además de las relaciones mutualistas y patogénicas que establecen. Se estima que la riqueza de hongos a nivel mundial es de 2,2 a 3,8 millones de especies, de las cuales unas 120 mil han sido descritas, lo que equivale en el mejor de los casos a un 8% del conocimiento de los hongos en el planeta (Hawksworth & Lücking 2017). En Bolivia, se estima que la riqueza de hongos podría superar las mil especies (Piepenbring 2003).
En el caso de los hongos gasteroides, un grupo ubicado taxonómicamente en la clase Agaricomycetes (Basidiomycota), se caracterizan por presentar la más amplia gama de formas y estructuras en sus basidiomas. Presentan como carácter común el desarrollo angiocárpico, i.e. permanecen cerrados hasta que las esporas alcanzan la madurez, y estas son del tipo estatismosporas, o sea que no son expulsadas activamente del basidio y dependen de mecanismos externos para su dispersión (Calonge 1998).
Los hongos gasteroides son pobremente conocido en el país, con una riqueza que no supera las 40 especies (Rocabado et al. 2007). Históricamente se conocen colectas de estos hongos efectuadas por H.H. Rusby en la Amazonia boliviana (Stevenson & Cárdenas 1949) y colecciones de basidiomas de hongos gasteorides obtenidas por Rolf Singer en localidades de los departamentos de Pando y Beni en 1956 (Wright 1960, Suárez & Wright 1996).
En el presente estudio, se analiza la abundancia de los hongos gasteroides y se examina la relación con variables medioambientales de la región (precipitación, temperatura, entrada de luz del dosel y la cobertura del sotobosque), a partir de cuatro localidades del norte de la Amazonia boliviana, con la finalidad de aportar con información al conocimiento de la ecología de este grupo de hongos en el país.
Área de estudio
La llanura amazónica boliviana presenta un clima tropical, con una temperatura promedio anual que, dependiendo de la zona, oscila entre los 24 y 27°C, en tanto que la precipitación promedio va desde los 1.200-2.000 mm/año, con una estacionalidad marcada de las lluvias que ocurren entre los meses de diciembre y marzo, aunque estas condiciones son influenciadas, por ejemplo por el fenómeno de El Niño (Pouilly et al. 2004) entre otros diversos factores. El incremento de las lluvias origina que áreas extensas de bosque y pastizales, principalmente las de suelos mal drenados y las cercanas a los ríos, se inunden estacionalmente (Pouilly et al. 2004). Navarro (2011) indica que el área de estudio se ubica en la región amazónica, provincia amazónica suroccidental, sector del Acre y Madre de Dios. Este distrito presenta un relieve de llanura ondulada con distintos grados de disección fluvial.
Sitios de muestreo
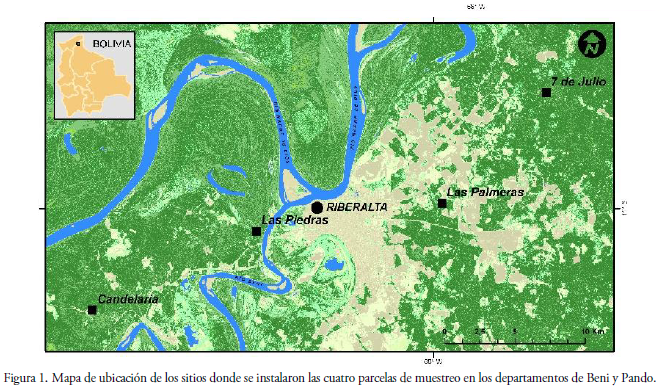
El presente estudio se realizó en cuatro localidades ubicadas en el norte de la Amazonia boliviana: Las Palmeras, 7 de Julio, Las Ruinas y Candelaria (Fig. 1). La comunidad de Las Palmeras se encuentra a 7 km al este de la población de Riberalta (10°59'48.4"S, 65°59'39.7"W, 120 m), Provincia Vaca Diez, Departamento de Beni. La comunidad 7 de Julio se localiza a aproximadamente 17 km al NE de Riberalta, de la misma provincia y departamento (10°55'34.1" S, 65°55'39.5" W, 107 m). Las Ruinas se sitúa a 1.5 km de la comunidad Las Piedras (11°00'52,6"S; 66°06'47,3"W, 152 m), Provincia Madre de Dios, Departamento de Pando, mientras que el bosque de Candelaria se encuentra a 10 km de Puerto Gonzalo Moreno de la misma provincia y departamento (11°03'54.8"S y 66°13' 05.5" W, 155 m).
Comunidad Las Palmeras. Se caracteriza por presentar un bosque secundario, denso y bajo con abundantes especies epífitas y lianas, presenta un dosel de 8-10 m de altura donde los géneros más representativos son Protium (isigo), Sapium (leche-leche) y Talisia. Los emergentes de 15 m de altura están representados por Apeiba tibourbou (cabeza de mono) y Didymopanax morototoni (guitarrero). El sotobosque se encuentra conformado por especies como Bactris major (marayaú) y otras de los géneros Warszewiczia sp. y Psychotria sp., entre otras (Fig. 2a).
Comunidad 7 de Julio. El bosque de tierra firme en este sitio es alto y denso con aproximadamente 1015 m. de altura y cuenta con árboles emergentes de Bertholletia excelsa (castaña) de hasta 3035 m., y especies como Phenakospermum guianensis (patajú gigante), Protium sp. (isigo) y Clarisia ilicifolia. El sotobosque se encuentra conformado en gran parte por numerosos helechos de los géneros Adiantum, Asplenium, Theypterisy Trichomanes (Fig. 2b).
Las Ruinas. Estructuralmente, la vegetación de este sitio se caracteriza por presentar un bosque secundario algo ralo y bajo de aproximadamente unos 810 m de altura, con presencia de árboles emergentes de Brosimun sp. de hasta 15 m. Entre las especies arbóreas más comunes se registró a Attalea phalerata (motacú), Ouratea sp. (almendrillo) y Triplaris sp. (palo diablo). Se observó la presencia de numerosas lianas y muchos claros de bosque dentro de la parcela establecida (Fig. 2c).
Candelaria. Este sitio presenta un bosque de tierra firme denso y alto de 1520 m, con árboles emergentes de Aspidosperma rigidum (gabetillo) y Ocotea sp. (negrillo) que miden 2535 m de altura. Entre las especies más representativas de la zona tenemos al Phenakospermum guianensis, Pseudolmedia laevis, Clarisia ilicifolia y Duguetia sp. La cobertura del suelo se encuentra dominada por numerosas especies de helechos pertenecientes a los géneros Adiantum, Alsophila Asplenium, Tectaria y Trichomanes (Fig. 2d).

Métodos
Instalación de parcelas. Para el estudio de los hongos gasteroides se empleó una variante del protocolo de muestreo para parcelas de tamaño fijo para hongos propuesta por Mueller et al. (2004). Se realizó una preselección de los tipos de bosque tomando en cuenta el estado de conservación y altura de la vegetación. Posteriormente se instalaron cuatro parcelas de 50x50 m, una por cada sitio de estudio, donde cada parcela estaba subdividida en cinco transectos de 50 m de largo separadas unas de otras por 10 m, formando un total de cinco carriles. A cada carril se le asignó una letra única (A-E), al medio de cada carril se fue marcando con una estaca cada 5 m y luego a cada estaca se le asignó un número secuencial (A1-A10; B1-B10;...E1-E10) con el propósito de poder diferenciar una subparcela de otra e identificar mejor las muestras colectadas. Se instalaron las subparcelas circulares de 5 m alrededor de cada estaca. Por cada transecto se instalaron 10 subparcelas lo que significó un total de 50 subparcelas por sitio de estudio. Adicionalmente, se realizaron colectas generales en los alrededores y/o lugares cercanos de las parcelas y a lo largo de caminos vecinales de la región.
Colecta de especímenes. Los muestreos se realizaron entre los años 2006 y 2007, en los cuatro sitios de estudio. En el primer año se efectuaron colectas durante los meses de noviembre-diciembre (estación lluviosa), mientras que en el segundo año se muestreó en los meses de agosto y septiembre (época seca). La colecta de hongos gasteroides fueron realizadas tres veces por estación a intervalos de muestreo de dos semanas y en cada sitio. Se colectaron muestras de basidiomas maduros y en buen estado en las subparcelas establecidas. Luego se registraron los siguientes datos: sustrato, color, tamaño, datos del lugar, coordenadas, fecha, número de subparcela. De los basidiomas en mal estado e incompletos, solo se registraron datos de color, sustrato, subparcela y fecha, pero no fueron colectados. Posteriormente las colectas fueron depositadas en contenedores plásticos con varios compartimentos o en bolsas de papel con sus respectivos datos. Para la identificación de los especímenes se realizaron análisis macroscópicos donde se observaron diferencias marcadas en cuanto a los caracteres morfológicos, clasificándose de esta manera a nivel de morfoespecies para lo cual también se utilizó literatura especializada (Martín 1988, Wright 1960, Soto & Wright 2000). Las muestras fueron secadas y depositadas en la colección científica de micología en el Herbario USZ del Museo de Historia Natural Noel Kempff Mercado.
Medición de la cobertura de vegetación del sotobosque. Para evaluar la cobertura del sotobosque se tomaron datos en las épocas húmeda y seca en cada una de las subparcelas de estudio empleándose un densiómetro que consistió de una hoja de acetato con 25 cuadrados de 3x3 cm dibujados con marcador indeleble, en base al método propuesto por Mostacedo & Fredericksen (2000). Se pudo tomar los datos desde afuera de cada subparcela con el acetato hacia abajo y según la cantidad de cuadrados que esta abarco estimándose la vegetación del sotobosque en porcentajes.
Medición de la entrada de luz. La entrada de luz del dosel fue evaluada tanto en la época húmeda como en la seca y se utilizó el mismo acetato empleado para medir la cobertura, pero esta vez los datos fueron tomados desde el centro de cada subparcela con el acetato hacia arriba para poder determinar la cantidad de luz observada a través de los cuadritos, estimándose de esta manera en porcentajes.
Abundancia. En el análisis se utilizó como unidad de muestreó a cada subparcela y se efectuó una sumatoria de todos los basidiomas colectados en ambas épocas y de esta manera se obtuvo la abundancia para cada parcela. Para determinar las diferencias de abundancia entre la época seca y húmeda para cada uno de los sitios de estudio, se utilizó el análisis de varianza (ANOVAs, P< 0.05) de dos factores (época y localidad). La prueba de Tukey fue usada para definir las diferencias entre las medias y verificar cuan similares o diferentes fueron los resultados obtenidos.
Cobertura de sotobosque y entrada de luz. Para evaluar la cobertura de vegetación del sotobosque y la entrada de luz del dosel se obtuvo el promedio de todas las subparcelas para cada uno de los bosques de estudio en ambas épocas y para el análisis se utilizó ANOVAs. La relación de la abundancia de hongos con la cobertura de vegetación del sotobosque y la entrada de luz del dosel, se calculó a través del análisis de correlación lineal de Pearson. Estos análisis se hicieron utilizando el programa estadístico STATISTICA.
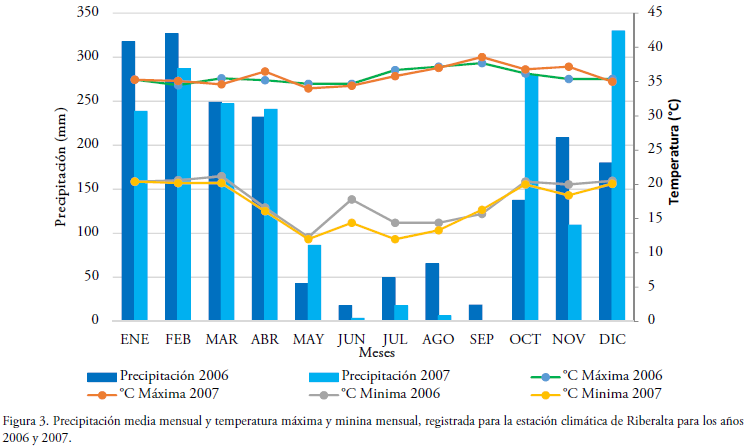
Precipitación y temperatura. Los datos diarios precipitación y temperatura máxima utilizados provienen de la estación climática de la ciudad de Riberalta procedentes del SISMET del Servicio Nacional de Meteorología e Hidrología (SENHAMI, www.senamhi.gob.bo) para los años 2006 y 2007 (Fig. 3). Para estimar como afectarían la precipitación/temperatura sobre la abundancia, se calculó la acumulación de lluvia (mm) y los promedios de la temperatura máxima (°C) en 7 y 14 días antes de la aparición de los basidiomas. La relación de la abundancia de los basidiomas con la precipitación y la temperatura máxima, se calculó a través del análisis de correlación lineal de Pearson.
Resultados
Géneros de hongos gasteroides
Durante el muestreo se lograron colectar 63 basidiomas de hongos gasteroides de los géneros Geastrum y 17 para Lycoperdon para la época húmeda, en tanto que para la época seca solo se encontró 3 basidiomas del género Geastrum (Fig. 4). Adicionalmente, en las áreas externas a las parcelas se identificaron basidiomas de los géneros Phallus en el departamento de Pando y Cyathus en el departamento de Beni.

Variación en la abundancia de hongos por sitios
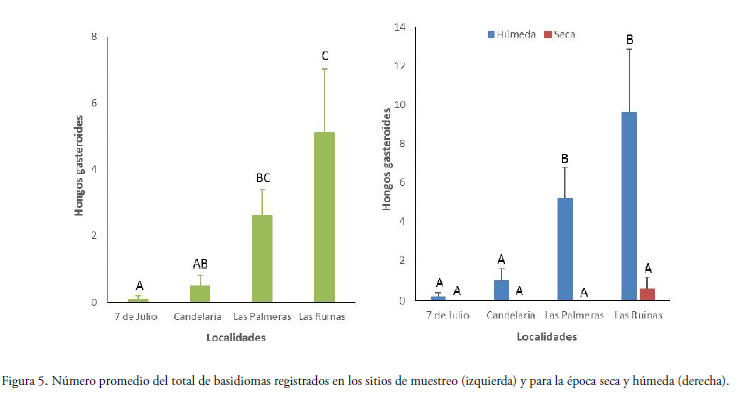
La abundancia de hongos gasteroides entre los sitios de estudio mostró una variación significativa (ANOVAs, F = 8.78; P = 0.0002), donde el bosque de Las Ruinas obtuvo el mayor número de hongos gasteroides, mientras que la parcela de la localidad 7 de Julio presentó la menor cantidad de basidiomas colectados. Las localidades de Candelaria y Las Palmeras mostraron valores intermedios a los otros sitios (Fig. 5).
Variación en abundancia de hongos por sitio y época
La abundancia de hongos fue significativamente diferente entre épocas de muestreo (ANOVAs, F = 33.14, P = 0.0001), siendo mayor en la estación húmeda, mientras que en la época seca solo se encontraron dos basidiomas del género Geastrum en Las Ruinas (Fig. 5). Se observó como la estación influyó en la abundancia de hongos para cada uno de los sitios de estudio, donde los bosques de Las Ruinas y Las Palmeras presentaron los mayores números de basidiomas colectados durante la época húmeda, siendo diferente en el resto de las localidades (Fig. 5).
Cobertura del sotobosque según las épocas por sitio
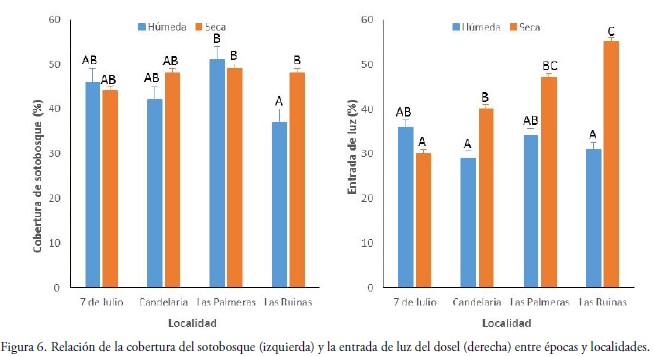
La cobertura de vegetación del sotobosque osciló entre el 5 y 80% durante la época seca, en tanto que en la época húmeda fue del 25 al 70%. En la época seca la cantidad de cobertura para todos los sitios se mantiene constante mientras que en la época húmeda es variable (Fig. 6), por lo que la cobertura de vegetación entre épocas no fue significativa (F = 2.83, P = 0.1025). Sin embargo, se observó una interacción significativa entre localidad y estación (F = 3.71, P = 0.0213), presentándose en la estación húmeda mayor cobertura en la localidad de las Palmeras y menor cobertura en la localidad de las Ruinas.
Entrada de luz del dosel según la localidad y épocas
La entrada de luz del dosel osciló entre el 5 y 100% durante la época seca, en tanto que en la estación lluviosa fue entre el 5-95%. Los niveles altos de porcentaje en algunas parcelas se deben a claros presentes en el bosque. Los análisis realizados muestran una interacción significativa entre localidad y época (F= 8.27 P= 0.0015), lo cual sugiere que la variación de la entrada de luz depende de la localidad. Se observa que las localidades Candelaria, Las Palmeras y Las Ruinas mostraron diferencias significativas entre épocas (Fig. 6). Solo en la localidad 7 de Julio la entrada de luz fue algo similar entre ambas épocas.
Relación entre abundancia con cobertura del sotobosque y entrada de luz
Estadísticamente el grado de relación entre la cobertura de vegetación del sotobosque y la abundancia de hongos para ambas épocas (Fig. 7), demostró que existe una tendencia negativa (R2 = 0.09, P = 0.066). En cuanto a la relación entre la entrada de luz del dosel y la abundancia de hongos para ambas épocas (Fig. 7), se observó que el número promedio de basidiomas tiene una tendencia negativa (R2= 0.04, P = 0.2447). Es decir, que a mayor porcentaje de cobertura y de entrada de luz es menor la cantidad de basidiomas.
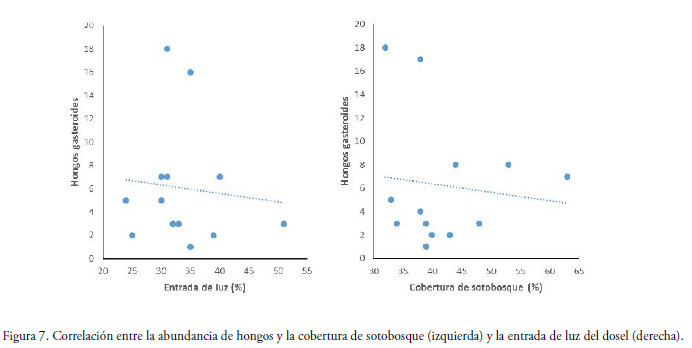
Relación entre abundancia con la precipitación
La relación entre la precipitación acumulada en un tiempo de una semana antes del muestreo y la abundancia de hongos para ambas épocas, se observó que el número de basidiomas tiene una tendencia positiva leve pero no significativa (R2= 0.311, P = 0.129) (Fig. 8). En tanto que, para la precipitación acumulada en un tiempo dos semanas, presenta también una tendencia positiva similar (R2= 0.328, P= 0.108) (Fig. 8).
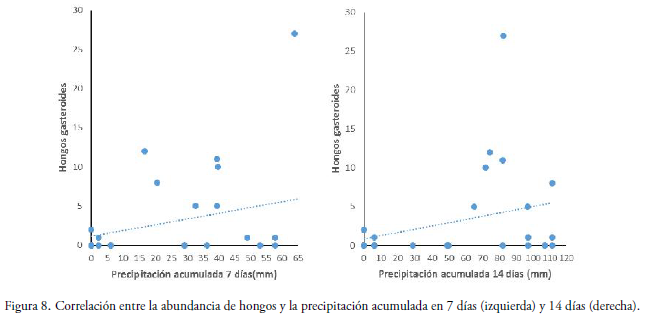
Relación entre abundancia con la temperatura máxima
La relación entre el promedio de la temperatura máxima en un tiempo de una semana antes del muestreo y la abundancia de hongos (número total de basidiomas) para ambas épocas, se observó que el número de basidiomas tiene una tendencia negativa no significativa (R2= 0.178, P = 0.384) (Fig. 9). Mientras que, para el promedio de la temperatura máxima en un tiempo dos semanas, presenta también una tendencia negativa similar (R2= 0,233, P = 0.250) (Fig. 9).
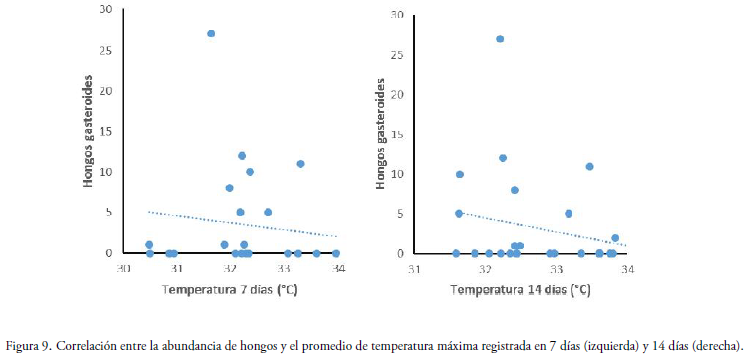
Discusión
La micobiota en la región amazónica es muy diversa (Lodge et al. 1995). Sin embargo, se encuentra insuficientemente estudiada, principalmente para el grupo de los hongos gasteroides. Tan solo en Brasil, la diversidad de gasteroides en la Amazonia en 2009 representaba el 9% del total de 238 especies (Trierveiler-Pereira & Baseia 2009, Trierveiler-Pereira et. al. 2009). Contrariamente en Bolivia, Colombia, Ecuador, Guyanas, Perú, Surinam y Venezuela el estado de conocimiento de los hongos es aún menor.
En el caso de Bolivia, hasta ahora las únicas referencias conocidas de hongos gasteroides para la Amazonia (Suárez & Wright 1996, Wright 1960) estaban representadas por cinco especies: Lycoperdon costaricense, L. fuligineum, Mutinus bambusinus, Phallus indusiatus, Staheliomyces cinctus, es decir el 12% del total reportadas para el territorio nacional (40 especies) en relación a la lista presentada por Rocabado et al. (2007). Estas colectas fueron efectuadas en dos localidades del Departamento de Pando (Conquista, Las Piedras) y tres en el Beni (Riberalta, Guayaramerin, Arroyo Florida).
Si bien no fueron encontradas durante el presente trabajo especies de los géneros Mutinus y Staheliomyces, se colectaron especies de Cyathus, Phallus, Geastrum y Lycoperdon (=Morganella). Los géneros Geastrum y Lycoperdon que fueron colectados en las parcelas de estudio son los primeros registros para los Departamentos de Beni y Pando en relación a Rocabado et al. (2007). De igual manera, las colectas de basidiomas de Cyathus en la localidad de Hamburgo (Beni) y el hallazgo de Phallus en Las Ruinas (Pando) durante las búsquedas no estandarizadas y colectas generales en áreas externas a las parcelas, se constituyen en los primeros reportes para estos Departamentos (Rocabado et al. 2007), ampliando notoriamente su área de distribución de este género en el país.
Considerando la carencia de información espacial y temporal de estos hongos, así como también los listados para otras regiones de la Amazonia en general (p.e., Gómez & Gazis 2006, Gazis 2007, Trierveiler-Pereira et. al. 2009, Trierveiler-Pereira et al. 2009), se podría esperar que otros géneros tales como Arachnion, Bovista, Calvatia, Clathrus, Crucibulum, Mutinus, Myriostoma, Pseudocolus, Sclerangium, entre otros, pudieran estar también presentes en el área de estudio, ampliando aún más la diversidad fúngica para esta región del país.
En este estudio, el género Geastrum fue el más dominante, lo cual también ha sido reportado para otros estudios de hongos gasteroides en regiones áridas, semiáridas y húmedas de México, Brasil y Argentina (Esqueda-Valle et al. 2000, Trierveiler-Pereira et al. 2013, Hernández Caffot et al. 2019). Esto podría deberse a la estrategia de dispersión de las esporas (Trierveiler-Pereira et al. 2013).
Algunos autores (Miller 1995, Mueller et al. 2004) mencionan que la composición de los hongos puede variar de un año a otro y es por este motivo que se requieren de estudios de por lo menos cinco años para tener una buena estimación de la diversidad. Es bajo esta premisa, que no se descarta la posibilidad que la baja diversidad de géneros gasteroides registrada en los cuatro sitios de estudio este influenciada por la duración del muestreo de campo, aunque puede estar relacionada al tipo de metodología empleada (p.e. número de parcelas por bosque).
También, podrían ser otros los factores bióticos y abióticos que se encuentren interviniendo directamente en la diversidad de macrohongos en el lugar. Al respecto, Mueller et al. (2004) indican que estos factores van desde la naturaleza del hábitat (p.e. pastizal o bosque, natural o artificial, etapas de sucesión o maduro), diversidad de especies de plantas (particularmente plantas hospederas), diversidad de sustrato, lugar geográfico (latitud, elevación), tipo de suelo y el clima.
En el presente estudio, durante la época húmeda se registraron ambos géneros (Geastrum y Lycoperdon), con la diferencia de que en la estación seca solo se colectaron únicamente especímenes deshidratados del género Geastrum pero en buen estado. Este fenómeno en la época seca podría deberse a la baja precipitación pluvial y humedad necesaria que estos hongos requieren para poder desarrollar los basidiomas. La precipitación es un factor climático importante, igual o más que la temperatura, en la determinación de la composición de la comunidad de hongos (Gazis 2007). En un estudio realizado por Esqueda-Valle et al. (2000) en México, encontraron la mayor diversidad de hongos gasteroides en verano, periodo en que prácticamente se presentaron las lluvias. Asimismo, el estudio de los hongos gasteroides en el noreste de Brasil que compararon las estaciones seca y húmeda, no hubo diferencias significativas entre períodos (Trierveiler-Pereira et al. 2013). Hasta ahora son muy pocos los estudios que se han realizado en hongos gasteroides y su relación con factores bióticos y abióticos, y más aún en la Amazonia. Algunos autores (p.e., Wright 1987) indican que el pH, fertilidad y el contenido del humus, pueden ser factores muy relevantes para la presencia de hongos gasteroides en un sitio.
La presencia de estos hongos también podría estar influenciada por la topografía del sitio, la concentración de arcilla, o la disponibilidad de materia orgánica en descomposición en el suelo, lo cual debería favorecer al establecimiento y su desarrollo. Si bien Alexopoulos (1962) y Suárez-Duque (2004) indican que la presencia de hojarasca y otros materiales (palitos, troncos) son considerados como un recurso indispensable para el desarrollo de los hongos, los resultados obtenidos demuestran lo contrario, ya que las localidades de 7 de Julio y Candelaria presentaron una cantidad de hojarasca en el suelo abundante y muy notoria, inclusive mucho más que en las Ruinas, lo que debería proporcionar un sustrato más diversificado y denso para el desarrollo de la comunidad de gasteroides (y otros hongos).
Braga-Neto et al. (2008) en una investigación de hongos de hojarasca efectuado en la Amazonia de Brasil (Manaus), descubrieron que la presencia de estos hongos estuvo influenciada por el contenido de arcilla en el suelo, pero fueron más determinantes los factores climáticos (precipitaciones).
Los factores que han influenciado en la abundancia de hongos gasteroides en cada sitio de estudio aún son inciertos, pero parece obedecer más a una interacción entre factores que a un solo factor aislado. Se descarta la posibilidad de que se deba a la metodología utilizada, ya que la frecuencia de parcelas de estudio refleja el área mínima ocupada por las especies en un área y tan solo provee una estimación aproximada de la abundancia (O'Dell et al. 2004).
El tipo de vegetación es uno de los factores que podría influir en la abundancia de hongos, ya que se esperaría que en los bosques con mayor estatura y con una complejidad estructural puedan dar lugar al establecimiento de más microhábitats y microclimas para los hongos, beneficiando al desarrollo de basidiomas. Gazis (2007) encontró que en los bosques amazónicos primarios de tierra firme existe una menor abundancia en relación a los bosques secundarios. Los resultados obtenidos en el presente trabajo muestran algo similar, ya que la mayor abundancia de hongos se registró en los bosques secundarios de Las Palmeras y Las Ruinas, áreas que presentan un dosel bajo (810m), en tanto que en los bosques 7 de Julio y Candelaria se caracterizaron por tener una mayor altura (1020 m) y una abundancia menor. Sin embargo, se necesitan de mayores estudios que permitan obtener evidencias claras para poder confirmar esta postura.
Por otro lado, en cuanto a las épocas del año (húmeda y seca), se observó una diferencia marcada, ya que en la época húmeda la cantidad de basidiomas en cada área de estudio fue mucho mayor que en la época seca en la que casi nada logró obtenerse. Sin embargo, esto no es un argumento para aseverar que las especies encontradas estuvieron ausentes en la época seca, sino que aparentemente el desarrollo de basidiomas no se llevó a cabo durante esta estación del año. Es importante señalar que el micelio de los hongos está debajo de la tierra o del sustrato que lo contenga y los basidiomas solo se hacen visibles en periodos reducidos (Rossman et al. 1998). Resultados similares en la Amazonia donde la abundancia de especies disminuyó en los meses de baja precipitación fueron obtenidos por Braga-Neto et al. (2008). A su vez, Suarez-Duque (2004) en un estudio basado en la diversidad y análisis estructural de hongos en el bosque de Mindo Lindo (Ecuador), en el que se incluyen los gasteroides, encontró que la abundancia de hongos fluctuó mensualmente por las características climáticas y la fenología de las especies.
La presente investigación demuestra que la abundancia de hongos esta directa e inversamente relacionada con la cobertura del sotobosque y la entrada de luz respectivamente. Esto significa que a mayor cantidad de entrada de luz, mayor cobertura de sotobosque y menor cantidad de hongos. Coincidentemente, Suárez-Duque (2004) mencionó que los hongos no son iguales a las plantas que compiten por la entrada de luz cuando aparecen los claros en el bosque, pero que la abundancia se incrementa debido a la caída y descomposición de los árboles, ya que hay una mayor degradación de materia orgánica. Una mayor cobertura del dosel ayuda a un mejor desarrollo de los hongos y a la cobertura del sotobosque, ya que la entrada de luz entra por los lados y no directamente disminuyendo el impacto de los rayos solares (Suárez-Duque 2004).
Conclusiones
El presente estudio es el primer trabajo micológico de esta naturaleza realizado en el norte del país, en el cual se determinó la abundancia de hongos gasteroides, además de su relación con ciertos factores variables medioambientales (precipitación, temperatura, entrada de luz del dosel y la cobertura del sotobosque), ya que el conocimiento adquirido de este grupo hasta ahora estaba basado en colectas ocasionales. El bosque de Las Ruinas y Las Palmeras registraron la mayor abundancia de especímenes colectados, y fue evidente que en la época húmeda se encontró la mayor cantidad de basidiomas en los cuatro sitios de muestreo. En cuanto a la entrada de luz del dosel, cobertura del sotobosque, precipitación y temperatura máxima, no se encontró que exista una relación con la abundancia de los hongos gasteroides.
Agradecimientos
Al Museo de Historia Natural Noel Kempff Mercado, especialmente al Herbario del Oriente Boliviano (USZ) por brindarnos su apoyo logístico para concluir el presente trabajo y también por el apoyo con la colección científica de hongos. A la Christensen Found, que a través del Missouri Botanical Garden proporcionaron el financiamiento para poder realizar esta investigación. A Steve Churchill por toda su ayuda. A Eliana Calzadilla por su asistencia en la fase de campo. A Luzmila Arroyo por los aportes al manuscrito. A Betty Flores por su apoyo con los análisis de datos y correcciones. Además, dos revisores anónimos realizaron importantes correcciones al manuscrito.
Referencias
Alexopoulos, J. C. 1962. Introductory mycology. Second Edition. John Wiley & Sons, Nueva York. 613 p.
Braga-Neto, R., Luizão, R.C.C. Magnusson, W.E. Zuquim, G. & de Castilho, C.V. 2008. Leaf litter fungi in a Central Amazonian forest: the influence of rainfall, soil and topography on the distribution of fruiting bodies. Biodiversity Conservation 17(11): 2701-2712. [ Links ]
Calonge, F.D. 1998. Gasteromycetes I. Lycoperdales, Nidulariales, Phallales, Sclerodermatales, Tulostomatales. Flora Mycologica Iberica. Vol. 3. Real Jardín Botánico Madrid.
Esqueda-Valle, M., E. Pérez-Silva, T. Herrera, M. Coronado-Andrade & A. Estrada-Torres. 2000. Composición de gasteromicetos en un gradiente de vegetación de Sonora, México. Anales del Instituto de Biología, Universidad Nacional Autónoma de México, Serie Botánica 71:39-62.
Gazis, R. 2007. Evaluation of the macrofungal community at Los Amigos Biological Station, Madre de Dios, Perú. Tesis de maestría, Texas Christian University, Fort Worth. 102 p. [ Links ]
Gómez, L.D. & R. Gazis. 2006. Dos Gasteromycetes (Basidiomycotina, Fungi) de Perú. Brenesia 65: 71.
Hawksworth, D.L. & R.R. Colwell. 1992. Microbial Diversity: biodiversity amongst micro-organisms and its relevance. Biodiversity Conservervation 1: 221–226.
Hawksworth, D.L. & R. Lücking. 2017. Fungal diversity revisited: 2.2 to 3.8 million species. Microbiology Spectrum 5(4): FUNK-0052-2016.
Hernández Caffot, M.L., L.C. Pereyra, G.L. Robledo, L.S. Domínguez. 2018. Ecology of gasteroid fungi (Agaricomycetidae and Phallomycetidae) affected by different successional stages of Polylepis australis Bitt. woodlands of Córdoba, central Argentina. Nova Hedwigia 106 (3-4): 455-472.
Lodge, J.C., G. Samuels, F.A. Uecker, D. Desjardin, E. Horak, O.K. Miller, Jr., G.L. Hennebert, C.A. Decock, J. Ammirati, H.H. Burdsall. Jr., P.M. Kirk, D.W. Minter, R. Hailing, T. Laessøe, G. Mueller, S. Huhndorf, F. Oberwinkler, D.N. Pegler, B. Spooner, R.H. Petersen, J.D. Rogers, L. Ryvarden, R. Watling, E. Turnbull & A. J. S. Whalley. 1995. A survey of patterns of diversity in non-lichenized Fungi. Mitt. Eidgenöss. Forsch.anst. Wald Schnee Landsch. 70, 1: 157-173.
Martín, M.P.E. 1988. Aportación al conocimiento de las Higroforaceas y los Gasteromycetes de Cataluña. Sociedad Catalana de Micología. Barcelona. 508 p. [ Links ]
Miller, S.L. 1995. Functional diversity in fungi. Canadian Journal of Botany (Suppl. 1): 550-557.
Mostacedo, B. & T.S. Fredericksen. 2000. Manual de métodos básicos de muestreo y análisis en ecología vegetal. BOLFOR, Santa Cruz. 87 p.
Mueller, G.M., J.P. Schmit, M. Sabine, H. L. Ryvarden, T. E. O’Dell, D. J. Lodge, P. R. Leacock, M. Mata, L. Umañia, Q. Wu &. D.L. Czederpiltz. 2004 Recommended protocols for sampling macrofungi. Pp. 106-172. En Mueller, G. M., Bills, G. F. & Foster, M. S. (eds.) Biodiversity of fungi, inventory and monitoring methods. Editorial Elsevier Academic Press. 777 p.
Navarro, G. 2011. Clasificación de la vegetación de Bolivia. Centro de Ecología y Difusión Simón I. Patino, Santa Cruz, Bolivia. 713 p. [ Links ]
O'Dell, T. Lodge E., D.J. Mueller, & G.M. Mueller. 2004. Approaches to sampling macrofungi. Biodiversity of fungi: inventory and monitoring methods. Pp. 163-168 En Mueller, G. M., Bills, G. F. & Foster, M. S. (eds.) Biodiversity of fungi, inventory and monitoring methods. Editorial Elsevier Academic Press. 777 p.
Piepenbring, M. 2003. Hongos. Pp. 90-96. En Ibisch P.I. & G. Mérida (eds.). Biodiversidad: La riqueza de Bolivia. Estado de conocimiento y conservación. Editorial FAN, Santa Cruz de la Sierra, Bolivia.
Pouilly, M., S.G. Beck, M. Moraes R. & C. Ibañez (eds.) 2004. Diversidad biológica en la llanura de inundación del Río Mamoré. Importancia ecológica de la dinámica fluvial. Centro de Ecología Simón I. Patiño, Santa Cruz.
Rocabado, D.R., J.E. Wright, O.Z. Maillard & N.F. Muchenik. 2007. Catálogo de los hongos Gasteromycetes (Fungi: Basidiomycotina) de Bolivia. Kempffiana 3(1): 3-13.
Rossman, A., R. Tulloss, T. O'Dell & G. Thorn. 1998. Protocols for an all taxa biodiversity Inventory of fungi in a Costa Rican conservation area. Parkway Publishers, Boone. 195 p.
Soto, M.K. & J.E. Wright. 2000. Taxonomía del género Geastrum (Basidiomycetes, Lycoperdales) en la Provincia de Buenos Aires, Argentina. Boletin de la Sociedad Argentina de Botánica 34 (3-4): 185-201.
Stevenson, J.A. & M. Cardenas. 1949. Lista preliminar de los hongos de Bolivia. Lilloa 21: 77–134.
Suárez, V.L. & J.E. Wright. 1996. South American Gasteromycetes V: The genus Morganella. Mycologia, 88(4): 655–661.
Suárez-Duque, D. 2004. Diversity and Structural Analisys of Aphyllophorales of the Protected Forest "Mindo Lindo" Pichincha province, Ecuador. Lyionia 17(1): 84-89. [ Links ]
Trierveiler-Pereira, L. & I. G. Baseia. 2009. A checklist of the Brazilian gasteroid fungi (Basidiomycota). Mycotaxon 108: 441–444.
Trierveiler-Pereira, L., A.C. Gomes-Silva & I.G. Baseia. 2009. Notes on gasteroid fungi of the Brazilian Amazon rainforest. Mycotaxon 110: 73-80.
Trierveiler-Pereira, L., P.J.P. Santos & I.G. Baseia. 2013. Ecological aspects of epigeous gasteromycetes (Agaricomycetes, Basidiomycota) in four remnants of the Brazilian Atlantic Forest. Fungal Ecology (6): 471-478.
Wright, J.E. 1960. Notas sobre Faloides sud y centroamericanas. Lilloa 30: 339–363.
Wright, J.E. 1987. The genus Tulostoma (Gasteromycetes). A world monograph. Bibliotheca Mycologica 113, Cramer, Stuttgart. 338 p. [ Links ]

