











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkEcología en Bolivia
versión impresa ISSN 1605-2528versión On-line ISSN 2075-5023
Ecología en Bolivia vol.53 no.2 La Paz set. 2018
Artículos
Baccharis boliviensis (Asteraceae), ¿especie clave de ambientes semiáridos andinos?
Baccharis boliviensis (Asteraceae), a key species in Andean semiarid environments?
Stephanie Torrico-Paz1*, Ramiro P. López 1,2, Oscar C. Ayala1, L. Adilen Fernández-Paz1, Nayra M. Antezana1 & Emmilce Morillas1
1Carrera de Biología, Facultad de Ciencias Puras y Naturales, Universidad Mayor de San Andrés, c/27 Cota Cota s/n Campus Universitario, La Paz, Bolivia. *Autora de correspondencia: s.torrico.paz92@gmail.com
2Herbario Nacional de Bolivia, Universidad Mayor de San Andrés, Casilla 10077- Correo Central, La Paz, Bolivia.
Resumen
La importancia del conocimiento de la estructura y dinámica de ambientes áridos y semiáridos de Bolivia es crucial, considerando que estos ambientes ocupan gran parte del territorio boliviano y hoy en día quedan pocos sectores de estos en buen estado. El estudio de patrones espaciales se considera adecuado para la comprensión de estos ecosistemas, lo que permitiría realizar planes de conservación y restauración. El presente estudio fue realizado en Mecapaca y Collana, en el valle de La Paz, con el objetivo de comprender los procesos potenciales que moldean la estructura de estas comunidades. Se muestrearon los individuos leñosos en una parcela por cada localidad, y se utilizó el índice de correlación de par g (r) para análisis univariados y bivariados alrededor de los primeros 500 centímetros de cada individuo utilizando tres diferentes modelos nulos. En total se muestrearon 840 individuos en Mecapaca (12 especies) y 721 en Collana (10). Se encontró un patrón agregado en la mayoría de las especies reportadas, siendo este más notorio en Collana que en Mecapaca. En ambas comunidades se encontraron asociaciones positivas entre la mayoría de las especies y B. boliviensis, lo que hace que esta sea posiblemente una especie clave que permite la sobrevivencia de otras plantas leñosas (facilitación biótica). Aún queda pendiente la realización de estudios sobre la relación de los arbustos y hierbas, y sobre patrones espaciales que involucren la relación de Prosopis alba (algarrobo) con otras especies para poder desentrañar la estructura y dinámica de estas comunidades.
Palabras clave: Agregación, Comunidad vegetal, Facilitación, Patrones espaciales, Valles secos interandinos.
Abstract
The importance of acquiring knowledge about the structure and dynamics of arid and semiarid environments in Bolivia is crucial, considering that these environments occupy a large part of Bolivian territory and are greatly degraded, with few patches in a good conservation state. The study of spatial patterns is considered appropriate to begin to understand ecosystems and their dynamics, allowing to make future plans for conservation and restoration afterwards. The present study was carried out in two sites within La Paz valley: Collana and Mecapaca, in order to understand the processes that structure these communities. Wooden individuals were recorded and measured in a single plot per locality, the pair correlation index g (r) was used for univariate and bivariate analyzes around the first 500 centimeters using three different null models. In total, 840 individuals in Mecapaca were measured, and 721 in Collana, recording five and six species, respectively. Most of the recorded species showed an aggregated pattern, this being stronger in Collana than in Mecapaca. In both sites, positive associations between most of the species and B. boliviensis were found making it likely that this species might be a keystone species that allows survival of other woody plants (biotic facilitation). Future studies about the relationship between woody and herbaceous species, and also the spatial relations between Prosopis alba (Mesquite tree) and B. boliviensis and other woody species may allow us to further gain insight into the structure and dynamics of these communities.
Key words: Aggregation, Facilitation, Interandean dry valleys, Plant community, Spatial patterns.
Introducción
Una de las concepciones clásicas de la ecología de comunidades sostiene que, en un gradiente ambiental, en su extremo más severo (mayor estrés) los organismos están limitados por su tolerancia al factor abiótico limitante; por el contrario, en el extremo más benigno (donde existe un menor estrés), la capacidad del organismo para competir contra otros es la clave del éxito (Connell 1961, Lomolino et al. 2006). Basados en esa premisa, se consideró que en los ecosistemas áridos, las interacciones bióticas desempeñaban un papel menor en comparación con la disponibilidad de agua (Noy-Meir 1973, Polis et al. 1997, Holmgren et al. 2006). Gran parte de esta imagen proviene de estudios realizados en desiertos mediterráneos, donde se registran las precipitaciones en invierno (Pugnaire et al. 1996, Armas & Pugnaire 2005). Estos ecosistemas dependen, en gran medida, de los eventos de El Niño. Otros desiertos, sin embargo, poseen lluvias más predecibles, como por ejemplo los desiertos con lluvias de verano, donde no se esperaría una dependencia tan fuerte del medio abiótico. De hecho, esta perspectiva fue cambiada en buena medida con el tiempo, pues varios estudios mostraron que en los ecosistemas áridos las interacciones podían desempeñar un papel de importancia en la organización de la comunidad vegetal (Flores & Jurado, 2003),
Debido a la visión competicionista predominante hasta los años 80 en el campo de la ecología, se verificó que en los desiertos la competencia por agua y nutrientes podía ser determinante (King & Woodel 1973, Tielbörger & Kadmon 2000, Maestre et al. 2006), constituyendo una interacción clave en este tipo de ecosistemas. Posteriormente, en la década de los 90 y comienzos de los 2000 surgieron diferentes trabajos que pusieron de relieve otro tipo de interacción, la facilitación (Bertness & Callaway 1994, Callaway 1995, Flores & Jurado 2003, Bruno 2003), la cual se define como el efecto positivo de una planta sobre sus vecinas, donde al menos una de las partes involucradas en la interacción recibe beneficios. Desde entonces se empezó a forjar el paradigma de que la organización de las comunidades desérticas se dapor un equilibrio entre competencia y facilitación (Callaway 2007). La hipótesis más conocida sobre interacciones en ambientes no solo desérticos, sino severos en general (caracterizados por sequía, frío, salinidad u otro factor estresante), es la hipótesis de gradiente de estrés (SGH, por sus siglas en inglés; Bertness & Callaway 1994), la cual postula que la competencia será predominante en el extremo húmedo de un gradiente de humedad, y la facilitación lo será en el otro.
En Bolivia, se conoce relativamente poco sobre cómo se organizan las comunidades en nuestros ambientes áridos y semiáridos, a pesar del enorme valor teórico y práctico que supondría tal conocimiento. Los estudios realizados en la Prepuna señalan la existencia de facilitación, aunque sugieren que ésta no parece ser muy fuerte para el caso de leñosas (López et al. 2007), y en cambio sí parece afectar la diversidad de hierbas (López & Ortuño 2008).
Una de las razones del poco conocimiento sobre ecología comunitaria se debe a la dificultad de realizar estudios a este nivel, en especial si de experimentos se trata. Por ello, las aproximaciones observacionales han adquirido gran relevancia para realizar estudios a esta escala. Uno de los enfoques más prometedores es el de los patrones espaciales, que consiste en mapear las coordenadas cartesianas de los individuos de una comunidad dentro de una parcela (Wiegand & Moloney 2004). Inicialmente, el estudio de patrones espaciales apuntaba simplemente a realizar descripciones de la estructura horizontal de las comunidades. Hoy en día, en gran parte gracias a la aparición de modelos nulos muy específicos y poderosos softwares (p.e., el software Programita; Wiegand & Moloney 2004, 2014), el estudio de patrones espaciales permite poner a prueba hipótesis sobre procesos subyacentes y es una herramienta muy valiosa para entender la organización de las comunidades biológicas (Wiegand & Moloney 2004).
Las zonas áridas de Bolivia ocupan una importante porción del país (López & Zambrana-Torrelio 2006). Gran parte de estas áreas ha sufrido procesos de erosión intensa, y hoy en día quedan pocos sectores de vegetación relativamente bien conservada. Por esta razón es fundamental entender aspectos básicos de estas comunidades si se pretende realizar posteriormente trabajos de restauración. El estudio de patrones espaciales es una buena manera de empezar a conseguirlo, pues permite obtener información fidedigna de manera rápida. La idea básica es utilizar los espacios de vegetación menos perturbados para conocer cómo se organizan las comunidades áridas de forma natural y es esto lo que se propone en este estudio.
En la cordillera Oriental de los Andes del valle de La Paz, todavía existen unas pocas zonas semiáridas de valles secos. Algunos estudios previos en estos valles evaluaron la influencia de Prosopis alba (erróneamente considerado P. flexuosa) sobre la diversidad y abundancia de especies leñosas y hierbas perennes (Larrea et al. 2005), la influencia de los arbustos sobre la distribución de cactáceas (López & Valdivia 2007) y la lluvia de semillas y la influencia de los granívoros sobre estas semillas (Terán 2008). Estos valles secos son los últimos de su tipo, y todavía resultan un buen laboratorio natural para comprender qué factores estructuran las comunidades áridas y semiáridas de los Andes.
Realizamos un estudio de patrones espaciales en dos zonas del valle de La Paz donde todavía existe vegetación nativa, con el objetivo de, a partir del conocimiento obtenido de las relaciones espaciales de las plantas, comprender cuáles son los procesos potenciales que parecen organizar estas comunidades. Este es un primer intento de abordar el tema de cómo se organizan las comunidades semiáridas del valle de La Paz y de ambientes parecidos.
Métodos
Área de estudio
Se muestrearon dos localidades con vegetación todavía relativamente bien conservada, una en Mecapaca (16°40'12"S; 68°00'00"W, 2.950 m) y la otra en Collana (16°41'45.42"S; 67°55'45.01"W, 3.500 m). El trabajo de campo se realizó en la estación lluviosa, en el mes de marzo de 2017 en Mecapaca y entre los meses de febrero y marzo del 2017 en Collana. Mecapaca posee la típica vegetación de los valles secos interandinos (Beck et al. 2015). La vegetación dominante del lugar consiste en matorrales de Baccharis boliviensis (Asteraceae) y Dodonea viscosa (Sapindaceae), con presencia de algunas especies de hábito arbóreo como Prosopis ferox y Acacia aroma (Fabaceae). El clima de la zona se caracteriza por tener una estación árida prolongada y una estación húmeda de cuatro a meses de duración (Beck et al. 2015).
Collana posee una vegetación de transición entre valles secos y Puna inferior. En la zona predomina la vegetación arbustiva compuesta por plantas espinosas y suculentas donde pueden encontrarse especies de los géneros Adesmia (Fabaceae), Proustia y Dasyphyllum (Asteraceae) y cactáceas, como Opuntia, Trichocereus,Oreocereus y Corryocactus. También se encuentran arbustos altos (de 2 a 2.5 m) de los géneros Caesalpinia y Acacia (Fabaceae), Dodonea, Baccharis, Ephedra (Ephedraceae), Agalinis y Calceolaria (Scrophulariaceae). El clima de esta zona es también semiárido, con temperaturas medias alrededor de 14°C y bajas precipitaciones, teniendo cuatro meses húmedos al año, todavía se registran heladas a esta altitud.
Muestreo
La parcela de Mecapaca tenía dimensiones de 50 x 20 m, mientras que la parcela en Collana de 30 x 10 m, una de las condiciones de nuestro diseño de muestreo fue el que las parcelas no presentaran ningún tipo de heterogeneidad visible, en este caso topográfica. Collana presenta una densidad de individuos alta, por lo que una parcela de 30 x 10 m fue considerada del tamaño apropiado. Mecapaca por su parte, tiene una densidad de individuos mucho más baja y se requirió de una parcela de mayor tamaño para conseguir más individuos.
En cada una de las parcelas mapeamos las coordenadas x,y de todos los individuos leñosos y suculentos de más de 20 cm de altura, exceptuando individuos de Echinopsis bridgesii (Cactaceae) de los cuales todos fueron considerados. De cada individuo muestrado se tomó además la altura y la cobertura (a partir del eje mayor de la copa y su perpendicular).
Análisis de datos
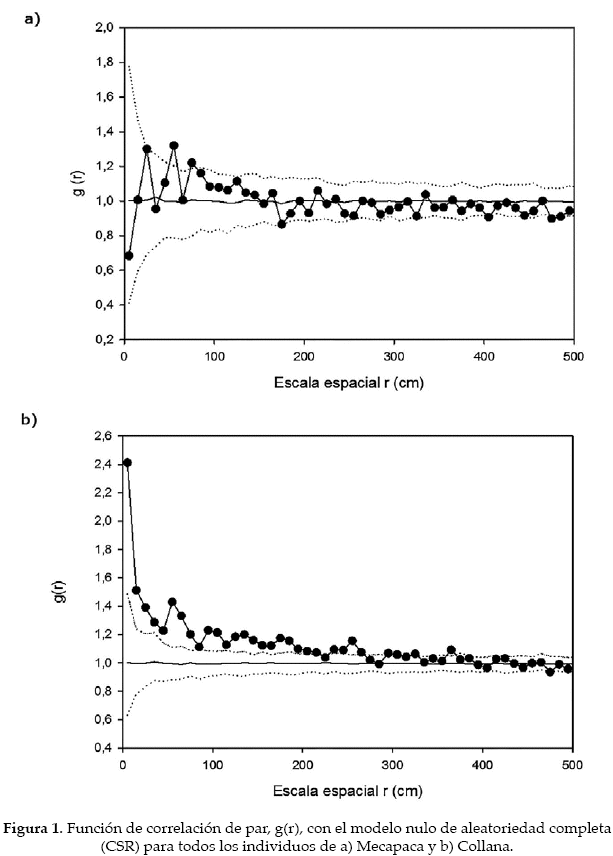
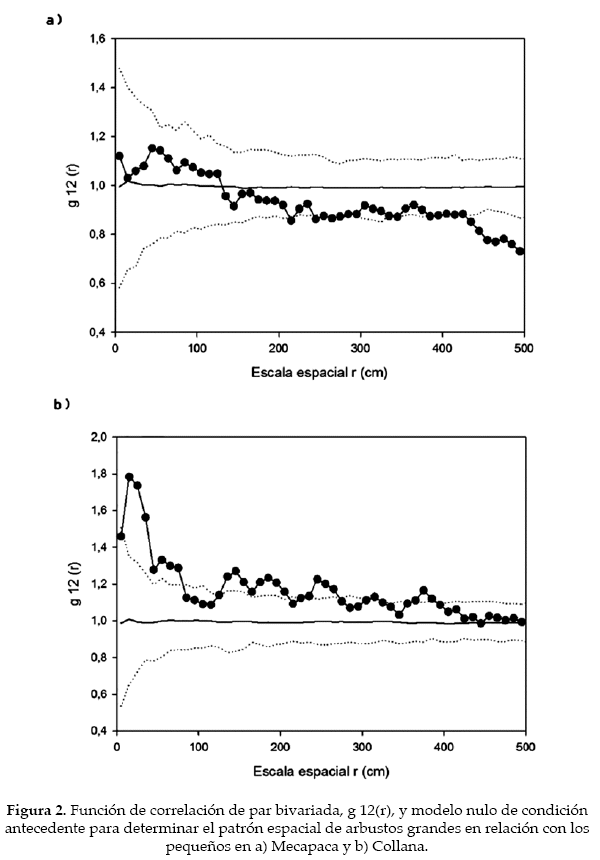
Para el análisis empleamos la función de correlación de par (pair-correlation function, g(r) (Stoyan & Stoyan 1994), que es considerada como uno de los mejores índices para la detección de patrones espaciales a diferentes distancias (Wiegand & Moloney 2004, Wiegand et al. 2013). La función g(r) se considera el equivalente no acumulativo de la función K de Ripley, y se entiende como la densidad esperada de puntos dentro de un anillo de radio r localizado en un punto (individuo) típico (Wiegand et al. 2013). La función g(r) para el caso de aleatoriedad espacial completa (CSR, de sus siglas en inglés) da un valor g(r) = 1. Los valores > 1 señalan que hay agregación para un patrón univariado (cuando todos los individuos muestreados se encuentran en una sola categoría; p.e., todas las leñosas de la comunidad o todos los individuos de una especie) y atracción para el caso de un patrón bivariado (cuando los individuos están divididos en dos categorías, las cuales son estudiadas en su relación recíproca; p.e., la distribución espacial de una especie 1 versus una especie 2; los individuos vivos versus los muertos). Los valores < 1 para un patrón univariado indican regularidad (o uniformidad), y repulsión si se trata de un patrón bivariado.
Para los análisis con la función g(r) se requieren, idealmente, >70 puntos (Wiegand & Moloney 2014), pero rara vez se alcanzan esos valores cuando se realizan múltiples análisis, de modo que en la literatura se ve el uso de valores en torno a 30 (a veces se usan tamaños incluso menores, que, cuando hay patrones muy marcados, pueden producir resultados significativos). Se emplearon simulaciones Montecarlo para poner a prueba la significación de los resultados en el caso de diferentes modelos nulos. Los límites de confianza (envueltas, o confidence envelopes en inglés; ver Wiegand & Moloney 2014) se obtuvieron a partir del quinto valor mayor y del quinto valor menor de las funciones tomadas de 199 simulaciones de los modelos nulos empleados (Wiegand & Moloney 2014). Intuitivamente, lo que hace el software es ver cuántos puntos (individuos) en promedio hay alrededor de cada uno de los puntos de la parcela dentro del anillo (cuyo ancho suele darlo el programa por defecto) para diferentes distancias, y eso se compara con los valores encontrados en las simulaciones para ver si hay más o menos de los esperados por el azar. Utilizamos tres modelos nulos: 1) el de aleatoriedad espacial completa (o total, CSR), 2) el de condición antecedente y 3) el modelo de independencia (toroidal shift). El primer modelo se empleó para todos los individuos de la parcela sin considerar la especie y para todos los individuos de cada una de las especies con altura mayor a 20 cm. El segundo se empleó para evaluar la relación espacial de los arbustos pequeños en las inmediaciones de los arbustos grandes y el tercer modelo se empleó para evaluar la asociación entre especies. Elegimos un umbral de 30 cm de altura para separar los individuos grandes (≥ 30 cm) de los pequeños (< 30 cm), debido a que encontramos un número relativamente grande de individuos entre el rango de 20 y 30 cm de altura (45 en Mecapaca, 77 en Collana) como para realizar los análisis. Además, la mayoría de los individuos de todas las especies de entre 20 y 30 cm se pueden considerar como individuos juveniles, de modo que la comparación puede considerarse como la de individuos adultos frente a individuos jóvenes.
Resultados
En las dos parcelas de los valles secos de La Paz inventariamos 840 individuos en Mecapaca y 721 en Collana. Todos los individuos censados fueron juveniles o adultos. Ambas comunidades presentaron un patrón agregado, en Mecapaca la agregación se observó solamente en un par de picos ubicados poco por encima de la envuelta, en Collana la agregación fue muy alta hasta los 200 cm (Fig. 1). La relación entre arbustos grandes y pequeños fue aleatoria en Mecapaca, tornándose uniforme a partir de los 400 cm; y marcadamente agregada en Collana hasta los 100 cm con pequeños picos (Fig. 2).
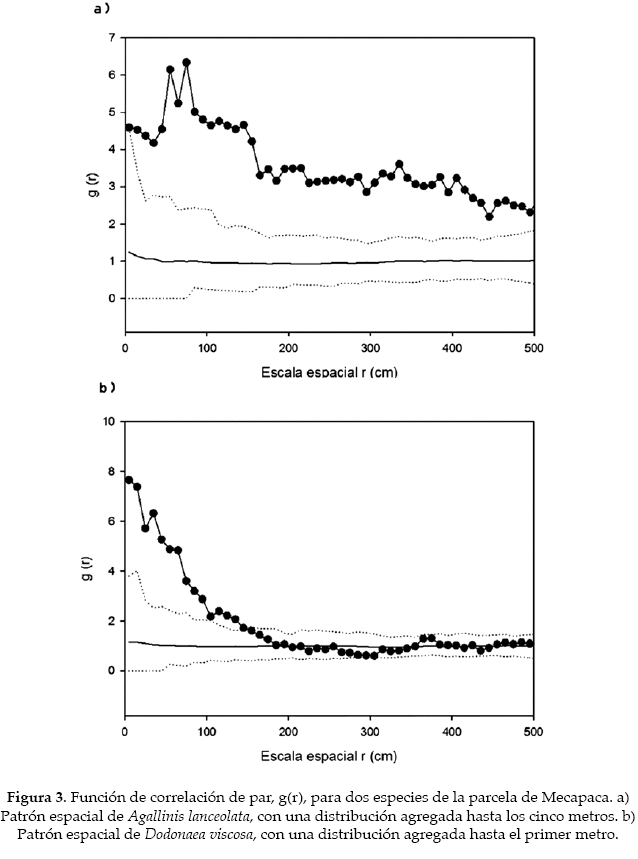
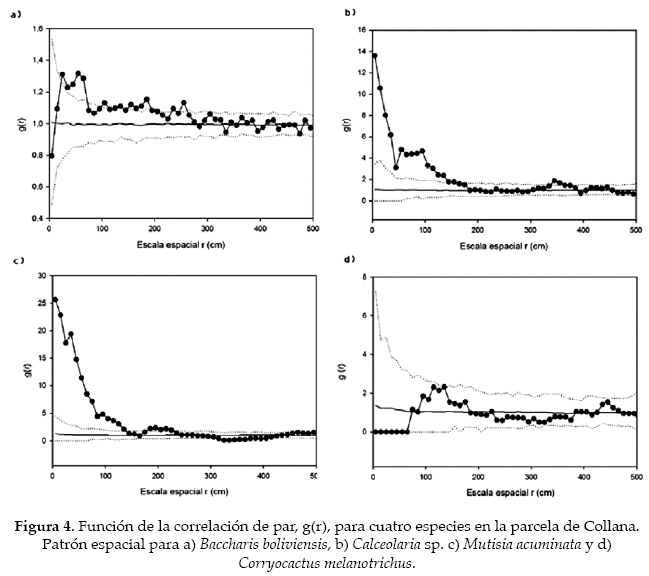
La distribución de las distintas especies de la comunidad de Mecapaca fue agregada en cinco de las seis especies, de las cuales Agallinis lanceolata, Echinopsis bridgesii y Verbesina sp. (Asteraceae) presentaron un patrón agregado hasta los 5 m, Dodonea viscosa hasta el primer metro (Fig. 3). Adicionalmente, Echinopsis bridgesii presentó un patrón agregado a lo largo de los 500 cm, al igual que Verbesina sp. Mientras que Baccharis boliviensis mostró un patrón levemente agregado entre los 50 y 150 cm. Solamente Corryocactus melanotrichus presentó una distribución aleatoria, aunque debido al bajo tamaño de muestra, no podemos estar seguros de que realmente sea así (Tabla 1). En Collana ocurrió algo parecido, ya que cinco de las seis especies presentaron patrones agregados: Baccharis boliviensis entre 20-80 cm, Calceolaria sp. hasta los 130 cm y Mutisia acuminata (Asteraceae) hasta los 120 cm. Para C. melanotrichus, se vio una ligera tendencia a la uniformidad hasta los 80 cm, siendo después su distribución aleatoria (Fig. 4). Por otra parte, Satureja boliviensis (Lamiaceae) presentó un patrón agregado hasta el primer metro y Minthostachys sp. (Lamiaceae) presentó el mismo patrón hasta los primeros 80 cm. La única especie que no mostró un patrón agregado fue nuevamente C. melanotrichus, lo que refleja que el patrón espacial de esta especie no es un efecto del bajo tamaño de muestra (Tabla 1).
Respecto alas asociaciones interespecíficas, en las dos localidades aproximadamente la mitad de éstas presentaron atracción, mientras que la otra mitad fue aleatoria. En ambos lugares, la mayoría de las especies se encontró asociada positivamente a Baccharis boliviensis. En Mecapaca Echinopsis bridgesii presentó una interacción positiva en los primeros 50 cm y Verbesina sp. presentó una interacción positiva en los primeros centímetros, con algunas tendencias irregulares de repulsión entre 170-260 cm (Fig. 5) (Tabla 2.). En Collana, Mutisia acuminatay Calceolaria sp. presentaron interacciones positivas con B. boliviensis, para el caso de M. acuminata, la interacción positiva se presentó entre 60-230 cm, mientras que para el caso de Calceolaria sp. el patrón agregado fue evidente solo en los primeros 30 cm e irregularmente entre los 250 y 350 cm (Fig. 6, tabla 2).
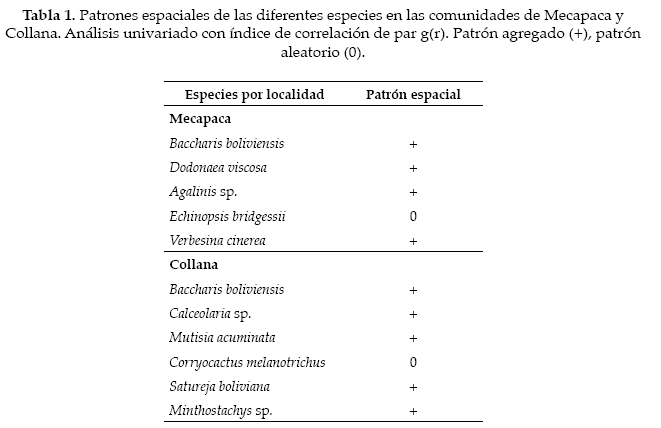
Discusión
Las dos comunidades muestreadas presentan varios patrones en común. En ambas hay un patrón comunitario agregado, aunque éste es mucho mayor en Collana. La mayoría de las especies presenta este patrón excepto el cactus C. melanotrichus; aproximadamente la mitad de las asociaciones entre especies es positiva (hay atracción). Pero lo que más destaca es que en ambas comunidades B. boliviensis, la especie dominante por cobertura o densidad, muestra un patrón de atracción con la mayoría de las otras especies de la comunidad.
Un patrón de asociación interespecífica de este tipo podría indicar facilitación. B. boliviensis ya ha sido mencionada como potencial nodriza en un estudio anterior (López et al. 2007). No obstante, el patrón encontrado en nuestro estudio también podría indicar preferencias de hábitat compartidas. Creemos que esta última opción queda descartada por varias razones. Primero, solo una parte de las especies asociadas a B. boliviensis muestran a su vez asociación positiva entre ellas (que es lo que se esperaría si compartieran características específicas de hábitat). Se podría aducir que en el caso de algunas de ellas, debido a su baja densidad, no necesariamente coincidirían en su ubicación en todos o en la mayoría de los presuntos parches de hábitat favorable (p.e., los individuos de una de las especies, por azar, cayeron en cierta fracción de los parches que es diferente de la fracción en que cayeron los individuos de la otra especie) y esto es plausible. No obstante, no creemos que dentro de las parcelas (que fueron elegidas en función de su uniformidad) exista una distribución tan heterogénea de los recursos. Por otro lado, a fin de exponer una razón más objetiva de por qué pensamos que las asociaciones positivas no dependen de las preferencias compartidas de las especies por ciertos microhábitats, podemos mencionar la distribución y abundancia de B. boliviensis en ambas comunidades. La especie es abundante y se encuentra distribuida por toda la comunidad, incluso en lugares donde no aparecen individuos de las otras especies, lo que sugiere que B. boliviensis podría no encontrarse asociada a ciertos parches con mejores recursos.
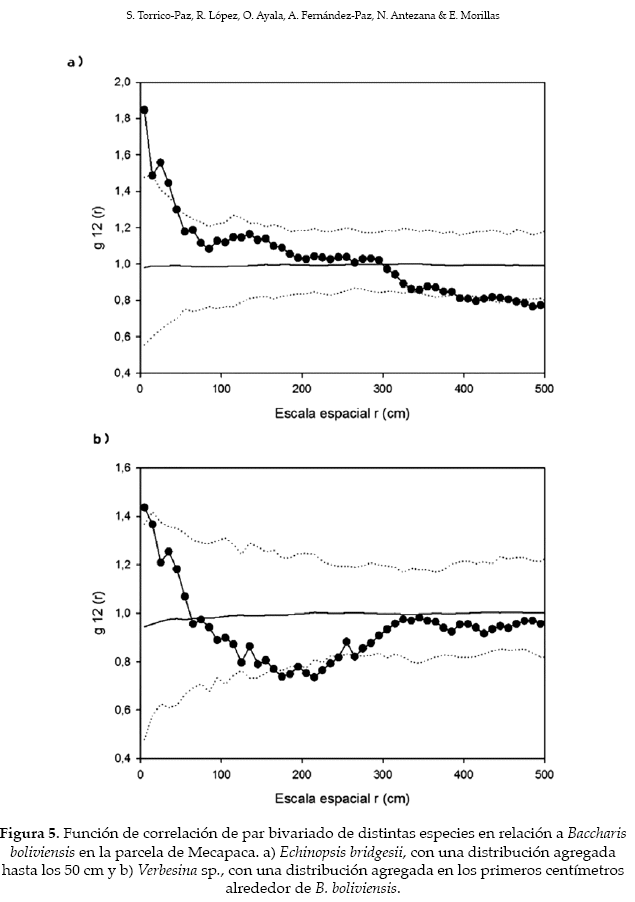

La facilitación, en su connotación clásica, es una interacción mediante la cual una planta (la nodriza) genera una mejora de las condiciones de microhábitat para otras plantas cercanas (las beneficiarias), las cuales pueden completar sus ciclos de vida en ese microambiente favorable (Bronstein 2009). Pero esta interacción también puede suponer la protección de las plantas contra herbívoros (facilitación biótica según Bertness & Callaway 1994, Smit 2009). Nosotros creemos que en los valles de La Paz se produce el segundo tipo de facilitación. Esta conclusión surge a partir de la observación de una importante presión de grandes herbívoros domésticos (vacas, caballos, burros) en estos valles, los cuales tienen una presencia casi constante durante todo el año (R.P.L., obs. pers.). Estos deben moverse en medio de una matriz de B. boliviensis (y en los lugares mejor conservados la matriz también incluye a Prosopis alba y Acacia aroma, que sin embargo tienen bajas densidades en la zona de estudio). Cualquier planta o arbusto que quede fuera de esa matriz corre el riesgo no solo de ser devorada, sino y quizá más importante, de ser pisoteada. Resultaría ventajoso para las otras especies de la comunidad asociarse a los espacios cercanos a B. boliviensis.
Otra línea de evidencia que permite inferir que la asociación positiva relacionada con B. boliviensis se debe a facilitación biótica es que en lugares de mayor aridez que los de nuestra zona de estudio (p.e., la Prepuna), las asociaciones positivas entre plantas no son tan marcadas (López et al. 2007, 2010) como las que nosotros encontramos en este estudio. En la Prepuna, el único tipo de ganado que suele afectar a las plantas es el caprino y evidentemente las plantas corren menos riesgos con un animal pequeño, que come y pesa mucho menos. En la Prepuna, la facilitación parece existir principalmente como proceso para enfrentar el ambiente abiótico.
Dado que concluimos que el patrón de atracción puede deberse a facilitación, quisimos hacer alusión a un estudio anterior que abordaba esta misma interacción con otras especies (Larrea et al. 2005). Estas leguminosas son las dominantes en muchos lugares de estos valles y en todo caso son las especies más altas. Debido al bajo número de individuos, no se pudo evaluar la relación espacial de las leguminosas (Prosopis alba, el algarrobo) y Acacia aroma con las otras leñosas. Larrea et al. (2005) encontraron que algunas especies de leñosas parecen preferir los espacios bajo el algarrobo y que éste ayuda levemente a aumentar la diversidad de la zona. Curiosamente, una de las tres especies que tenían mayor asociación espacial con P. alba fue B. boliviensis (que era más abundante bajo el algarrobo). A diferencia de nuestro caso, el estudio de Larrea y colaboradores se realizó en un lugar cercano al del presente trabajo pero localizado en una terraza aluvial muy amplia con vegetación en mejor condición que en nuestra parcela (tanto en términos de cobertura del algarrobo como del número de especies); lamentablemente esta diversa comunidad fue deteriorada por acciones edilicios.
Basados en la distribución de B. boliviensis en nuestra parcela, queda claro que es una especie que no depende de la presencia de otras especies, y que es más bien un taxón que permite el establecimiento de otras leñosas. Podríamos decir que se trata de una especie clave, ya que parece permitir que ciertas especies sean parte de los ecosistemas de valles secos de La Paz o que al menos tengan abundancias relativamente mayores de las que serían posibles en un ambiente más abierto. El por qué se encontró en mayor abundancia bajo el algarrobo en aquel estudio queda como incógnita. Una posibilidad es que crece mejor en suelos con mayor contenido en nitrógeno, ya que el suelo bajo P. alba tiene una concentración diez veces mayor que en espacios abiertos (López & Valdivia 2007).
Nuestros resultados son tanto más sorprendentes cuanto que lo que se conoce de B. boliviensis indica que se trata de una especie con efectos potencialmente dañinos. Cazón et al. (2000) encontraron que extractos de las hojas de B. boliviensis inhibieron completamente la germinación del cactus Trichocereus pasacana. De todos modos, sus efectos negativos no han sido demostrados en ambientes naturales y es posible que diferentes poblaciones de esta especie tengan una química diferente en función del ambiente en el que evolucionaron. Si esta última posibilidad fuera correcta, no dejaría de llamar la atención que sean justamente las poblaciones del extremo más árido donde crece B. boliviensis las que generen aleloquímicos (generalmente se esperaría que las plantas de los lugares con condiciones físicas menos severas fueran las más agresivas desde esta perspectiva ya que están rodeadas de más especies y de más individuos). Sin embargo, si las poblaciones de B. boliviensis de La Paz poseen las mismas sustancias alelopáticas que las poblaciones argentinas y si estas sustancias son capaces de actuar en condiciones naturales, la asociación de varias especies con B. boliviensis tendría que explicarse por protección contra agentes bióticos. Posiblementees preferible tolerar un ambiente más o menos hostil cerca de B. boliviensis que uno en extremo difícil en espacios más abiertos transitados por grandes animales. Si bien no es imposible que una especie con aleloquímicos sea a la vez una planta facilitadora (Farzam & Ejtehadi 2016), no es muy plausible que sean muchas las especies de plantas que puedan crecer debajo de estas inusuales nodrizas. Sería interesante plantear un estudio de las relaciones químicas entre las especies de este tipo de ambientes.
Otro aspecto que es necesario discutir es lo poco que se sabe acerca de la influencia de los arbustos sobre la distribución y abundancia de hierbas en estos ambientes. En la Prepuna más árida, un estudio (López & Ortuño 2008) encontró una influencia positiva de los arbustos sobre la diversidad. Considerando las similitudes entre la Prepuna y los valles paceños, se podría esperar el mismo patrón en los valles paceños. Es un tema que tendrá que ser evaluado ojalá en el futuro inmediato.
Conclusiones
Los resultados del presente trabajo muestran un patrón agregado en la distribución espacial de las especies vegetales de los valles secos interandinos, tanto en la comunidad completa (en el caso de Collana) como para determinadas especies, siendo pocas las excepciones que tienen un patrón diferente (p.e. Corryocactus melanotrichus). Por otra parte, nosotros encontramos una asociación positiva entre B. boliviensis y otras especies vegetales en ambas parcelas. Se podría llegar a pensar en esta especie como una especie nodriza en este ecosistema. Es más interesante este hallazgo sabiendo que existe un estudio que reporta efectos alelopáticos de esta especie sobre una especie de cactus, por lo que se sugiere realizar más estudios sobre B. boliviensis.
Nuestro estudio solo abarcó las especies arbustivas, dejando de lado a las hierbas circundantes y a los árboles con pequeño tamaño muestral. Queda pendiente, entonces, realizar un futuro estudio sobre la relación arbustos-hierbas y uno que aborde el tema de patrones espaciales que involucre a P. alba y a A. aroma en su relación con B. boliviensis y las otras especies de leñosas. Empezamos a desentrañar la estructura y función de estos ecosistemas, pero esperemos que no sea demasiado tarde, ya que se encuentran seriamente amenazados.
Agradecimientos
Los autores quieren agradecer a Freddy Zenteno, Natalio Roque, Laura Moya y David Villalba por su valiosa ayuda durante el trabajo de campo.
Referencias
Armas, C. & F. I. Pugnaire. 2005. Plant interactions govern population dynamics in a semi-arid plant community. Journal of Ecology 93(5): 978-989.
Beck, S., E. García, L.N. Thompson, R.I. Meneses, F. Zenteno, R.P. López & A. Fuentes. 2015. Paisajes, eco-regiones y vegetación. pp. 113-158. En: M. Isabel, R.I. Meneses & J. Sarmiento (eds.) Historia Natural de un valle en los Andes: La Paz. Instituto de Ecología y Museo Nacional de Historia Natural, La Paz.
Bertness, M. D. & R. Callaway. 1994. Positive interactions in communities. Trends in Ecology & Evolution 9(5): 191-193.
Bronstein, J.L. 2009. The evolution of facilitation and mutualism. Journal of Ecology 97(6): 1160-1170. [ Links ]
Bruno, J. F., J.J. Stachowicz & M.D. Bertness. 2003. Inclusion of facilitation into ecological theory Trends in Ecology & Evolution 18(3): 119-125.
Callaway, R. M. 1995. Positive interactions among plants. The Botanical Review 61(4): 306-349. [ Links ]
Callaway, R. M. 2007. Positive interactions and interdependence in plant communities. Springer Science & Business Media, Dordrecht. 404 p. [ Links ]
Cazón, A., M. de Viana & J.C. Gianello. 2000. Identificación de un compuesto alelopático de Baccharis boliviensis (Aster aceae) y su efecto en la germinación de Trichocereus pasacana (Cactaceae). Revista de Biología Tropical 48(1):47-51.
Connell, J.H. 1961. The influence of interspecific competition and other factors on the distribution of the barnacle Chthamalus stellatus. Ecology 42(1): 710-723. [ Links ]
Farzam, M., & H. Ejtehadi. 2016. Effects of drought and slope aspect on canopy facilitation in a mountainous rangeland. Journal of Plant Ecology 10(4): 626-633.
Flores, J., & Jurado, E. 2003. Are nurse-protégé interactions more common among plants from arid environments? Journal of Vegetation Science 14(6): 911-916.
Holmgren, M., P. Stapp, C.R. Dickman, C. Gracia, S. Graham, J.R. Gutiérrez, C. Hice, F. Jaksic, D. A. Kelt, M. Letnic, M. Lima, B. C. López, P. L. Meserve, W. B. Milstead, G. A. Polis, M. A. Previtali, M. Richter, S. Sabaté & F. Squeo. 2006. Extreme climatic events shape arid and semiarid ecosystems. Frontiers in Ecology and the Environment 4(2): 87-95.
King, T.J. & S.R. Woodell. 1973. The causes of regular pattern in desert perennials. Journal of Ecology 61(3): 761-765.
Larrea-Alcázar, D., R.P. López & D. Barrientos. 2005. The nurse-plant effect of Prosopis flexuosa D.C. (Leguminosae-Mimosoideae) in a dry valley of the BolivianAndes.Ecotrópicos 18(2): 89-95.
Lomolino M.V., B.R. Riddle & J. H. Brown. 2006. Biogeography. Sinauer Associates, Sunderland. 725 p.
López, R. P. & C. Zambrana-Torrelio. 2006. Representation of Andean dry ecoregions in the protected areas of Bolivia: the situation in relation to the new phytogeographical findings. Biodiversity & Conservation 15(7): 2163-2175.
López, R.P. & S. Valdivia. 2007. The importance of shrub cover for four cactus species differing in growth form in an Andean semi-desert. Journal of Vegetation Science 18(2): 263-270.
López, R.P., S. Valdivia, D. de la Quintana & N. Sanjinés. 2007. The role of nurse plants in the establishment of shrub seedlings in the semi-arid subtropical Andes. Oecologia 152(4): 779-790.
López, R.P. & T. Ortuño. 2008. La influencia de los arbustos sobre la diversidad y abundancia de plantas herbáceas de la Prepuna a diferentes escalas espaciales. Ecología Austral 18(1): 119-131.
López, R.P., D. Larrea Alcázar, T. Ortuño, F. Zenteno Ruiz & S. Valdivia. 2010. ¿Qué conocemos sobre la ecología de la Prepuna y otras zonas semiáridas de los andes centrales? Diez años de observaciones. pp. 207-216. En: Paniagua, N., S. Beck, R.L. López & N. Nagashiro (eds.), Biodiversidad y Ecología enBolivia. Instituto de Ecología, Universidad Mayor de San Andrés, La Paz.
Maestre, F. T., F. Valladares & J. F. Reynolds. 2006. The stress-gradient hypothesis does not fit all relationships between plant-plant interactions and abiotic stress: further insights from arid environments. Journal of Ecology 94(1):17-22.
Meserve, P. L., D.A. Kelt, W.B. Milstead & J.R. Gutierrez. 2003. Thirteen years of shifting top-down and bottom-up control. BioScience 53(7): 633-646.
Noy-Meir, I. 1973. Desert ecosystems: environment and producers. Annual Review of Ecology and Systematics 4(1): 25-51. [ Links ]
Polis, G. A., S.D. Hurd, C.T. Jackson & F.S. Piñero. 1997. El Niño effects on the dynamics and control of an island ecosystem in the Gulf of California. Ecology 78(6): 1884-1897.
Pugnaire, F. I., P. Haase, J. Puig de fábregas, M. Cueto, S.C. Clark & L.D. Incoll. 1996. Facilitation and succession under the canopy of a leguminous shrub, Retama sphaerocarpa, in a semi-arid environment in south-east Spain. Oikos 76(3): 455-464.
Smit, C., M. Rietkerk & M.J. Wassen. 2009. Inclusion of biotic stress (consumer pressure) alters predictions from the stress gradient hypothesis. Journal of Ecology 97(6): 1215-1219.
Stoyan, D. & H. Stoyan. 1994. Fractals, random shapes and point fields. John Wiley & Sons, Chichester. 389 p.
Terán, A. 2008. El rol de Prosopis flexuosa y Dodonaea viscosa como plantas nodriza para dos especies de cactus en el valle seco de Mecapaca. Tesis de licenciatura en biología, Universidad Mayor de San Andrés, La Paz. 90 p.
Tielbörger, K. & R. Kadmon. 2000. Temporal environmental variation tips the balance between facilitation and interference in desert plants. Ecology 81(6): 1544-1553.
Wiegand, T. & A. K. Moloney. 2004. Rings, circles, and null-models for point pattern analysis in ecology. Oikos 104(2): 209-229.
Wiegand, T. & K.A. Moloney. 2014. Handbook of spatial point-pattern analysis in ecology. CRC Press. Boca Ratón. 538 p.
Wiegand, T., F. He & S. P. Hubbell. 2013. A systematic comparison of summary characteristics for quantifying point patterns in ecology. Ecography 36(1): 92-103.
Manejado por: Mónica Moraes R.
Recibido en: 9 mayo 2018
Aceptado el: 1 julio 2018

