











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkEcología en Bolivia
versión impresa ISSN 1605-2528versión On-line ISSN 2075-5023
Ecología en Bolivia vol.53 no.1 La Paz abr. 2018
Artículo
Hábitos alimentarios de la lechuza blanca Tyto alba en un agroecosistema del centro-sur de Chile
Food habits of barn owl Tyto alba in an agroecosystem in southern central Chile
Alfredo H. Zúñiga1,2,*, Víctor Fuenzalida3 & Rodolfo Sandoval4
1Laboratorio de Ecología, Departamento de Ciencias Básicas, Universidad de Los Lagos, Casilla 933, Osorno, Chile. *Autor de correspondencia: zundusicyon@gmail.com
2Programa de Doctorado en ciencias mención conservación y manejo de Recursos Naturales, Universidad de Los Lagos, Puerto Montt, Chile.
3Consultora Ambientes del Sur, Temuco, Chile.
4Red de la Conservación de la Biodiversidad de Nahuelbuta, Contulmo, Chile.
Resumen
Se describe la dieta de la lechuza blanca (Tyto alba) en un agroecosistema ubicado a 20 km al sur de Mulchén, en el centro-sur de Chile. Ésta fue estudiada a través de la colecta sistemática de egagrópilas y posterior análisis de los restos de presas no digeridos presentes en éstas en laboratorio. Se destacó la presencia predominante de mamíferos (roedores; 86.74%), seguido por aves (passeriformes; 9.6%) y en una menor proporción, por invertebrados (3.6%). Son discutidos los elementos ecológicos que intervienen en el patrón dietario observado.
Palabras clave: Abrothrix, Chile, Dieta, Lagomorfos, Roedores.
Abstract
The Barn owl (Tyto alba) diet is described in an agroecosystem located at 20 km South of Mulchén, in south-central Chile. This was studied through the systematic collection of pellets and further analysis in laboratory. It was emphasized the predominant presence of mammals (rodents; 81.9%), followed by birds (passerines; 9.6%), and to a lesser extent, by invertebrates (3.6%). Ecological elements involved in this pattern are discussed.
Keywords: Abrothrix, Chile, diet, lagomorphs, rodents.
Introducción
La lechuza blanca (Tyto alba Scópoli 1769) es una rapaz nocturna de la familia Tytonidae, que cuenta con una distribución cosmopolita (Clark et al. 1978), a través de la cual vive en diversos hábitats, donde consume un amplio espectro de presas. Este espectro varía de acuerdo a las diversas latitudes en que las cuales esta rapaz está presente, lo que da cuenta de su flexibilidad ecológica (Smith et al. 1972, Vargas et al. 2002, Hernández-Muñoz & Macina 2011, Brito et al. 2015). En Chile, se encuentra la subespecie tuidara, la que se encuentra a lo largo de todo el territorio nacional (Pavez 2004), donde depreda sobre diversas especies de roedores, así como marsupiales y aves, cuya riqueza de especies está determinada por un gradiente latitudinal. Existen diversos estudios que documentan la diversidad dietaria de la lechuza en este continuo geográfico (Ebensperger et al. 1991, Correa & Roa 2004, Muñoz-Pedreros et al. 2016, Valladares et al. 2016).
A pesar de la flexibilidad ecológica de la lechuza, son escasos los estudios que den cuenta del proceso de transformación del paisaje, los que acontecen fundamentalmente a través de cambios en la cobertura de la vegetación arbustiva y arbórea. En el centro-sur de Chile, esta situación se va conformando de una manera sistemática y gradual en el tiempo (Echeverría et al. 2008), explicada principalmente por la consolidación local de las actividades forestales y agrícolas. En este contexto los agroecosistemas, cuya tecnificación productiva implica una extrema simplificación de la cobertura vegetal, trae como consecuencia una reducción de la biodiversidad en sus diversas formas (Benton et al. 2003), lo que llevaría a la conformación de una comunidad con pocas especies generalistas. Este hecho reviste especial relevancia para los depredadores, debido a la relación existente entre las prácticas agrícolas y la diversidad de presas, con consiguientes efectos sobre los depredadores (Arlettaz et al. 2010). Así, la lechuza podría adaptarse a este escenario concentrando su espectro alimentario en unas pocas especies que aún persistan en este agroecosistema, o bien modificar esta conducta a través del consumo de especies de gran adaptabilidad. El objetivo del presente estudio es documentar los hábitos alimentarios de T. alba en un agroecosistema del centro-sur de Chile, donde se compara la frecuencia de consumo de las distintas presas con su respectiva biomasa, lo que permitiría determinar su importancia relativa en el total observado.
Metodología
Área de estudio
El área de estudio es un agroecosistema que se encuentra a 20 km al sur de la ciudad de Mulchén, centro-sur de Chile (37°49'42''S, 72°14'51''W). Posee un clima húmedo (Di Castri & Hajek 1976) y desde el punto de vista vegetacional se clasifica como bosque caducifolio (Gajardo 1994). En la actualidad, esta zona presenta en forma predominante cultivos de avena y lupino, a través de los cuales se encuentran pequeños parches de bosque nativo, distribuidos en forma dispersa (Fig. 1).
Métodos
Durante la primavera de 2016 se recorrió el área de estudio en búsqueda de regurgitados (egagrópilas), los cuales fueron identificados por su morfología (Muñoz-Pedreros & Rau 2004), siendo además confirmados por la detección de árboles utilizados como perchas, hecho que coincide además con el avistamiento posterior de los individuos en el lugar. Fueron detectadas siete de estas perchas en el área de estudio, las cuales presentaron entre sí una distancia promedio de 100 metros. Las egagrópilas fueron depositadas en bolsas de papel, para su análisis posterior en laboratorio.
En laboratorio, se desmenuzaron las egagrópilas en forma manual, siendo los restos posteriormente identificados a nivel de especie a través de su observación con una lupa electrónica. La identificación estuvo basada principalmente en claves que dan cuenta de la morfología de las mandíbulas en el caso de roedores (Reise 1973, Pearson 1995), y de colecciones de referencia (provenientes del Laboratorio de Ecología Aplicada y Biodiversidad de la Universidad Católica de Temuco) para el resto de los taxa. Para la diferenciación entre conejos y liebres en el caso de los lagomorfos, se examinaron las escamas de sus pelos, cuya estructura es diagnóstica para cada especie (Day 1966).
Para cuantificar el espectro trófico de la lechuza se determinó la frecuencia relativa que cada item presentó en relación al total de egagrópilas analizadas (Rau 2000). Para estimar la diversidad de presas consumidas en el área de estudio, se calculó el índice de amplitud dietaria de Levins (β), el cual es un indicador de la amplitud del espectro de presas consumida por un depredador en función de 0 a n ítemes disponibles (Levins 1968). La desviación estándar de este índice fue estimada a través del método Jackkinfe (Jaksic & Medel 1987). Para evaluar el efecto de la biomasa de las presas en el patrón dietario se utilizó el método de las isoclinas tróficas (Kruuk & De Kock 1981), el cual ha sido aplicado para aves rapaces (Arriagada et al. 2011). En forma secundaria, se utilizó la media geométrica de los pesos de las presas (Muñoz-Pedreros et al. 2018). Los valores de los pesos de las presas fueron obtenidos a través de Muñoz-Pedreros & Gil (2009) y Norambuena & Riquelme (2014) para mamíferos y aves, respectivamente. En el caso de los roedores indeterminados, se utilizó la media aritmética de los pesos de las especies identificadas y para lagomorfos, se consideró lo determinado por Muñoz-Pedreros et al. (2018), debido a la evidencia de que en este grupo, las lechuzas depredan principalmente sobre juveniles (Simonetti & Fuentes 1982). La nomenclatura de las especies de roedores se basará en Muñoz-Pedreros & Gil (2009).
Resultados
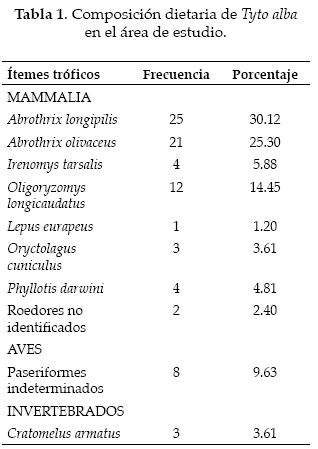
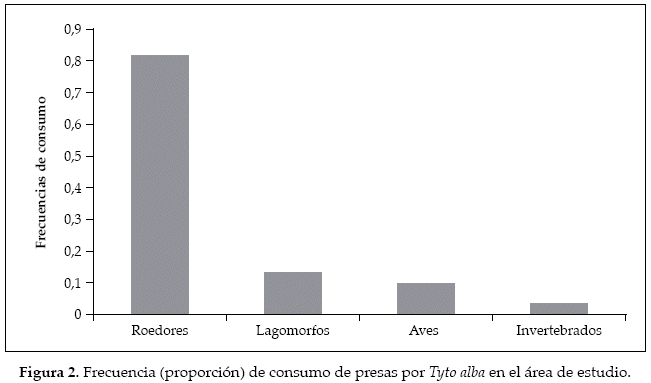
Se colectó un total de 56 egagrópilas, en las cuales se registraron 83 presas distribuidas en 10 categorías. De éstas, se destacaron los roedores que ocuparon una frecuencia del 86.74% del total (Tabla 1; Fig. 2), con un total de cinco especies, principalmente las del género Abrothrix, donde A. longipilis y A. olivaceus fueron las más consumidas. Por otra parte los lagomorfos, un grupo exótico y que está representado en el área de estudio por la liebre europea Lepus europaeus y el conejo, Oryctolagus cuniculus, fueron el otro grupo observado de mamíferos, presentando un 4.81% del porcentaje de consumo. Las aves fueron el segundo taxa representado como presa de T. alba, siendo el orden de las paseriformes el único grupo observado. Finalmente los artrópodos, representados esencialmente por el grillo rojo Cratomelus armatus, fueron los que conforman la menor proporción de la dieta de la lechuza. La amplitud dietaria observada fue de β=5.18±1.54.
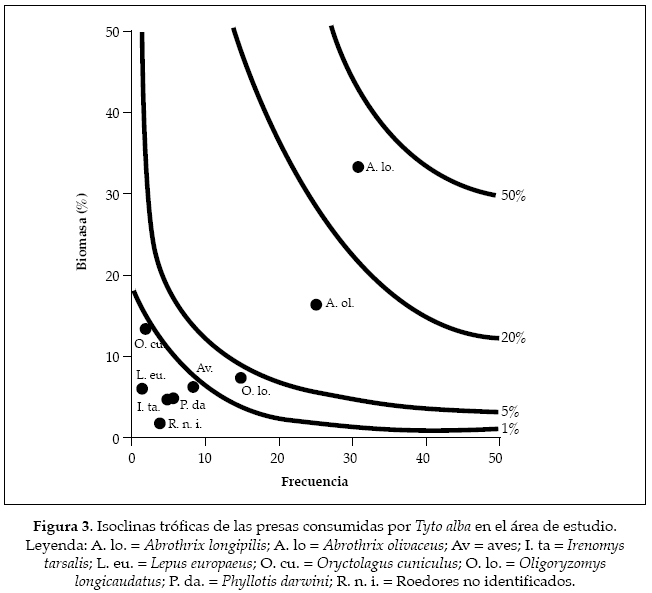
Al representar el efecto de la biomasa de las presas consumidas por la lechuza a través de las isoclinas tróficas, se puede apreciar que los lagomorfos ocupan la isoclina del 50%, mientras que los roedores ocuparon la inmediatamente inferior, es decir la del 20% (Fig. 3). Por otra parte, las aves ocuparon la isoclina más baja (1%) y los artrópodos, al representar un porcentaje mínimo en términos de biomasa en el total, no fueron incorporados en el análisis; asimismo, el valor de la media geométrica del peso de las presas fue de 37.19 gramos.
Discusión
El espectro trófico observado de la lechuza ocupa un amplio número de presas, donde los roedores ocupan la mayor parte de los ítemes que compone, lo cual es consistente con estudios similares (Rau et al. 1985, Muñoz & Murúa 1990, González et al. 2004). Estre patrón se ve reforzado a partir de la media geométrica observada, ya que este valor se sitúa en torno a la masa de las especies de roedores observadas. Asimismo, la concentración de dos de estas especies A. olivaceus y A. longipilis en la isoclina del 5 y 20% respectivamente, dan cuenta de la importancia en términos relativos del total del espectro trófico de la lechuza, ya que éstos son los que se encuentran en las isoclinas más elevadas en relación a las otras presas. Por otra parte, la riqueza de roedores observada da cuenta de las especies de mayor representación en esta latitud, lo que da cuenta de la flexibilidad espacial de éstas para utilizar agroecosistemas (Muñoz-Pedreros & Gil 2009), donde las variaciones de abundancia de las distintas especies estarían asociadas con dinámicas reproductivas específicas, asociadas tanto por factores endógenos como ambientales (Murúa & González 1986), lo cual se evidenciaría en la tasa de capturas. Además, la ventana temporal donde se realizaron las actividades coinciden con los episodios reproductivos, y por consiguiente de reclutamiento de juveniles (Murúa & González 1986, Muñoz-Pedreros & Gil 2009), lo que permite la representatividad de la diversidad de la comunidad de roedores local.
En términos de riqueza de especies, la conversión del bosque nativo original a un agroecosistema ha traído como consecuencia una sensible reducción en la ocurrencia de una importante proporción de roedores, ya que esta condición espacial afecta sus requerimientos específicos de hábitat (Muñoz-Pedreros & Gil 2009). A pesar de lo anterior, la aparición de la laucha arbórea (Irenomys tarsalis) en el espectro trófico de la lechuza permite suponer un gran esfuerzo de búsqueda por parte de T. alba a través del área de estudio, ya que este roedor posee hábitos arborícolas (Muñoz-Pedreros & Gil 2009), y los árboles disponibles se encuentran en los parches de bosque (Fig. 1), lo que sugiere que la lechuza blanca desplegaría parte de sus recorridos a lo largo de estos sectores. Este hecho implicaría para la lechuza un importante esfuerzo de búsqueda, considerando el ámbito de hogar que presenta (Hafidzi et al. 2012; Thomsen et al. 2014), y los lugares donde ha sido observado que anida, lo cual reviste especial importancia considerando el mosaico que conforman los parches de bosque nativo en torno a la matriz agrícola. Por otra parte, es destacable la ausencia del monito del monte (Dromiciops gliroides) en el espectro trófico observado, el cual es un marsupial de hábitos fundamentalmente arbóreos y que además, es una recurrente presa de T. alba en los bosques del sur de Chile, lo que pone de manifiesto las restricciones ecológicas que posee esta especie para utilizar los agroecosistemas, debido a sus altos requerimientos de cobertura arbórea y arbustiva, así comoa su reducido ámbito de hogar (Fontúrbel et al. 2010), lo que limita sus probabilidades de desplazamiento entre los parches de hábitat.
El consumo de lagomorfos en la dieta de la lechuza obedecería a limitaciones conductuales por parte de esta rapaz (Simonetti & Fuentes 1982), a lo cual debe adicionarse el hecho de que el tamaño promedio de la lechuza (470 g; Pavez 2004), restringe la captura sólo a los individuos juveniles (Simonetti & Fuentes 1982). Este hecho contrasta con otras localidades (Aliaga et al. 2004), donde especies de mayor tamaño, los cuyes (Cavia sp.) presentan una alta depredación, contrastando no obstante el hecho de que en este último caso las presas son nativas y por ende un mayor tiempo evolutivo en términos de interacciones depredador-presa. A pesar de lo anterior, los lagomorfos en el área de estudio ocupan una importante fracción de la biomasa total consumida (un poco menos del 10%), a pesar de presentar una baja frecuencia de captura, lo que supone una importante recompensa en términos energéticos. En comparación con los roedores, se evidencia que los lagomorfos presentan una menor importancia al ser comparados en las isoclinas tróficas, lo cual sugiere que los primeros resultan ser más fácilmente capturables. A pesar de esto último y dentro de la lógica estructural de los agroecosistemas, resulta interesante que los lagomorfos presenten en una alta ocurrencia en ambientes altamente antropizados (Jaksic & Fuentes 1988; Teta et al. 2012), lo cual sugiere que esta baja de consumo pueda ser consecuencia de una partición de nicho con otros depredadores (Zúñiga, datos no publicados). No obstante lo anterior, resulta de importancia evaluar la abundancia de los lagomorfos en el área de estudio (Batholomew 1970), para evaluar hipótesis relacionadas con respuestas funcionales de consumo.
Conclusiones
Tyto alba presenta un espectro trófico concentrado en roedores, siendo sus principales preferencias dos especies de un total de cinco, A. olivaceus y A. longipilis, las que exhiben la mayor flexibilidad ecológica en relación a la baja complejidad de la vegetación propia de un agroecosistema. Por otra parte, la baja ocurrencia de lagomorfos en este espectro sugiere que no existiría una selectividad por este grupo, lo cual es atribuible a un mayor esfuerzo de captura al considerar el mayor tamaño de éstos en relación a los roedores. Resulta de especial importancia estimar las abundancias de ambos grupos de presas para evaluar los efectos de su disponibilidad en el patrón alimentario observado.
Agradecimientos
Al laboratorio de Ecología Aplicada y Biodiversidad de la Universidad Católica de Temuco, por la facilitación de sus colecciones; a dos revisores anónimos que mejoraron consistentemente el manuscrito.
Referencias
Aliaga-Rossel, E. & T. Tarifa. 2005. Cavia sp. como principal presa de la lechuza de campanario (Tyto alba) al final de la estación seca en una zona intermedia al norte del Departamento de La Paz, Bolivia. Ecología en Bolivia 40: 35-42.
Arlettaz, R., M. Kräbenbühl, B. Almasi, A. Roulin & M. Schaub. 2010. Wildflower areas within revitalized agricultural matrices boost small mammal populations but not breeding barn owls. Journal of Ornithology 151: 553-564.
Arriagada, A.M., J.L. Arriagada, L.A. Baessolo & C.G. Suazo. 2008. Dieta estival del águila (Geranoetus melanoleucus) en la Región de Aysén, Patagonia chilena. Ecotrópicos 24: 164-171.
Bartholomew, B. 1970. Bare zone between California shrub and grassland communities: the role of animals. Science 170: 1210-1212. [ Links ]
Benton, T.G., J.A. Vickery & J.D. Wilson. 2003. Farmland biodiversity: is habitat heterogeneity the key? Trends in Ecology and Evolution 18: 182-188.
Brito, J., H. Orellana-Vásquez, H., Cadena-Ortiz, R. Vargas, G.M. Pozo-Zamora & J. Curay. 2015. Mamíferos pequeños en la dieta de la lechuza Tyto alba (Strigiformes: tytonidae) en dos localidades del occidente de Ecuador, con ampliación distribucional de Ichthyomys hydrobates (Rodentia: Cricetidae). Papéis Avulsos de Zoologia 55: 261-268.
Clark, R.J., D.G. Smith & L.H. Kelso. 1978. Working bibliography of owls of the world: summaries of currents taxonomy and distributional status. National Wildlife Federation, Washington DC. USA. 319 p.
Day, M. 1966. Identification of hair and feather remains in the gut and feces of stoats and weasels. Journal of Zoology 148: 201-207. [ Links ]
Di Castri, E. & E. Hajek. 1976. Bioclimatología de Chile. Ediciones Universidad Católica de Chile, Santiago, Chile. 159 p.
Ebensperger, L., J. Mella & J. Simonetti. 1991. Trophic-niche relations among Galictis cuja, Dusicyon culpaeus and Tyto alba in Central Chile. Journal of Mammalogy 72: 820-823.
Echeverría, C., D. Coomes, M. Hall & A. Newton. 2008. Spatially explicit models to analyze forest loss and fragmentation between 1976 and 2020 in southern Chile. Ecological Modelling 212: 439-449.
Fontúrbel, F. E., E. A. Silva-Rodríguez, N. H. Cárdenas & J. E. Jiménez. (2010) Spatial ecology of monito del monte (Dromiciops gliroides) in a fragmented landscape of southern Chile. Mammalian Biology 75: 1-9.
Gajardo, R. 1994. La vegetación natural de Chile, clasificación y distribución geográfica. Editorial Universitaria. Santiago. 65 p. [ Links ]
González, D., M. Auset, O. Skewes & R. A. Figueroa. 2004. Variación estacional en el consumo de roedores por la lechuza de campanario (Tyto alba) en un área suburbana de Chillán, centro-sur de Chile. Hornero 19: 61-68.
Hafdzi, M. N., M. H. Hamzah & M. S. Jamaluddin. 2003. Ranging behaviour of Tyto alba in ricefield from radio telemetry studies. Journal of Malasyan Applied Biology 32: 47-51.
Hernández-Muñoz, A. & C.A. Mancina. 2011. La dieta de la lechuza (Tyto alba) (Aves: Strigiformes) en hábitats naturales y antropogénicos de la región central de Cuba. Revista Mexicana de Biodiversidad 82: 217-226.
Jaksic, F. & R. Medel. 1987. El acuchillamiento de datos como método de obtención de intervalos de confianza y prueba de hipótesis para índices ecológicos. Medio Ambiente 8: 95-103.
Jaksic, F. & E. Fuentes. 1988. El conejo español: ¿un convidado de piedra? pp. 88-101, En: Fuentes, E. & S. Prenafeta (eds.). Ecología del Paisaje en Chile Central: Estudios sobre sus Espacios Montañosos. Ediciones Universidad Católica de Chile, Santiago.
Kruuk, H. & L. De Kock. 1981. Food and habitat of badgers (Meles meles L.) on Monte Baldo, northern Italy. Zeitschrift fur Saugertierkunde 46: 295-301.
Levins, R. 1968. Evolution in a changing environment. Princeton University Press, Nueva Jersey. 120 p. [ Links ]
Muñoz,A. & R. Murúa. 1990. Control of small mammals in a pine plantation (central Chile) by modification of the habitat of predators (Tyto alba, Strigiforme, and Pseudalopex sp., Canidae). Acta Oecologica 11: 251-261.
Muñoz-Pedreros, A. & J. Rau. 2004. Estudios de egagrópilas en aves rapaces. pp. 265-279. En: Muñoz-Pedreros, A. & J. Yáñez (eds.). Aves Rapaces de Chile. Centro de Estudios Agrarios y Ambientales Ediciones, Valdivia.
Muñoz-Pedreros, A. & C. Gil. 2009. Orden Rodentia. pp. 93-157. En: Muñoz-Pedreros, A. & J. Yáñez (eds.). Mamíferos de Chile. Centro de Estudios Agrarios y Ambientales Ediciones, Valdivia.
Muñoz-Pedreros, A., C. Gil, J. Yáñez, J. R. Rau & P. Möller. 2016. Trophic ecology of two raptors, barn owl (Tyto alba) and whitetail kite (Elanus leucurus), and possible implications for biological control of hantavirus reservoir in Chile. Wilson Journal of Ornithology 128: 385-397.
Muñoz-Pedreros, A., J. Yáñez, H.V. Norambuena & A. Zúñiga 2018. Diet, dietary selectivity and density of South american grey fox, Lycalopex griseus, in Central Chile. Integrative Zoology 13: 46-57.
Murúa, R. & L.A. González. 1986. Regulation of numbers in two rodent species. Revista Chilena de Historia Natural 59: 193-200.
Norambuena, H. & J. Riquelme. 2014. Profesor Dr. Francisco Behn Kuhn (1910-1976), biografía y catálogo de su colección de aves chilenas. Centro de Estudios Agrarios y Ambientales Ediciones, Valdivia.
Quintana, V. 2008. Diversidad de micromamíferos de la Región de la Araucanía, sur de Chile. Gestión Ambiental 16: 41-52. [ Links ]
Pavez, E. 2004. Descripción de las aves rapaces chilenas. pp. 29-103, En: Muñoz-Pedreros, A. & J. Yáñez (eds.) Mamíferos de Chile. Centro de Estudios Agrarios y Ambientales Ediciones, Valdivia.
Pearson, O. 1995. Annotated keys on near Nahuel Huapi National Park or Lanin National Park, southern Argentina. Mastozoología Neotropical 2: 99-148. [ Links ]
Rau, J. 2000. Métodos de ecología trófica. pp. 397-406. En: Muñoz-Pedreros, A. & J. Yáñez (eds.) Mamíferos de Chile. Centro de Estudios Agrarios y Ambientales Ediciones, Valdivia.
Rau, J., D. Martínez & J. Yáñez. 1985. Dieta de la lechuza blanca (Strigiformes) en el sur de Chile. Boletín del Museo Regional de la Araucanía 2: 134-135.
Reise. D. 1973. Clave para la determinación de los cráneos de marsupiales y roedores chilenos. Gayana 27: 1-20. [ Links ]
Simonetti, J.A. & E.R. Fuentes. 1982. Microhabitat use by European rabbits (Oryctolagus cuniculus) in central Chile: are adult and juvenile pattern the same? Oecologia 54: 55-57.
Smith, D., C. Wilson & H. Frost. 1972. Seasonal food habits of barn owls in Utah. Great Basin Naturalist 32: 230-234.
Teta, P., C. Hercolini & G. Cueto. 2012. Variation in the diet of western barn owls (Tyto alba) along an urban-rural gradient. Wilson Journal of Ornithology 124: 589-596.
Thomsen, S. K., C. E. Kroeger, P. H. Bloom & L. Harvey. 2014. Space use and homerange size of barn owls on Santa Barbara Island. Monographs of the Western North America 7: 339-347.
Valladares, P., N. Urrutia, N. Álvarez, O. Orellana & S. Alvarado. 2016. Dieta de la lechuza blanca (Tyto alba Scópoli 1769) en el valle de Copiapó, desierto de Atacama, Chile. Interciencia 41: 114-119.
Vargas, J., C. Landaeta & J.A. Simonetti. 2002. Bats as a prey of barn owls (Tyto alba) in a tropical savanna in Bolivia. Journal of Raptor Research 36: 146-148.
Manejado por Javier López de Casenave
Recibido en: 21 agosto 2017
Aceptado en: 3 diciembre 2017

