











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkEcología en Bolivia
versión impresa ISSN 1605-2528versión On-line ISSN 2075-5023
Ecología en Bolivia v.52 n.1 La Paz mayo 2017
Artículo
Análisis multi-temporal de mosaico de etapas sucesionales del bosque chaqueño de Arenales, Parque Nacional Kaa-Iya del Gran Chaco
Multi-temporal analysis of a vegetation mosaic form by a serial stages of the Chacoan forest on well drained soils, Kaa-Iya del Gran Chaco National Park
Carlos Pinto1 & Erika Cuéllar Soto2*
1 Fundación Amigos de la Naturaleza, FAN, Santa Cruz Bolivia.
2University of Oxford, Dept Zoology, Wildlife Conservation Research Unit, Recanati Kaplan Centre, Tubney OX13 5QL, Oxon, United Kingdom, * Autor de correspondencia: erika.cuellar71@gmail.com
Resumen
Según estudios previos la vegetación original de la zona sureste del Parque Nacional Kaa-Iya fue el bosque chaqueño de arenales. Sin embargo, actualmente el paisaje está conformado por un mosaico de etapas seriales o sucesionales en diversos grados de madurez. Para visualizar y cuantificar situaciones de retracción o invasión de comunidades de vegetación, desarrollamos un estudio de los cambios estructurales de la vegetación en un área determinada a través del tiempo. Basados en imágenes satelitales y utilizando herramientas de los sistemas de información geográfica producimos tres mapas de vegetación luego de una clasificación supervisada para tres décadas 1975, 1984 y 1999. Además realizamos los cálculos de superficie por comunidad vegetal para cada año y comparamos estos entre años. Observamos la mayor retracción (91.5%) de la comunidad de pajonal herbazal y el aumento de formaciones arbustivas leñosas. Esperamos que la información obtenida en este trabajo contribuya a entender la pérdida de estos ecosistemas abiertos y el hábitat para especies dependientes de las pampas chaqueñas y promover así un mejor manejo de las mismas.
Palabras clave: Análisis multi-temporal, Arborización, Gran Chaco, Imágenes satelitales, Mapas de vegetación.
Abstract
According to previous studies the original vegetation of Southeast corner of the Kaa-Iya National Park was the Chacoan forest on well drained soils. However, the current landscape is formed by a mosaic of serial stages in different degrees of maturity. In order to visualize and quantify situations of retraction or invasion of vegetation communities, we developed a study of the structural changes of the vegetation in a determined area over time. We used satellite images and geographic information systems tools to produced three vegetation maps after a "supervised classification" for 3 decades 1979, 1984 and 2009. We calculated the extension of each plant community for each year and compare these between years. We observed the greatest retraction (91.5%) of the grassland community and the increase of woody shrub formations. We hope that the information obtained in this work will help to understand the loss of open ecosystems and the habitat for species dependent on the Chacoan grasslands and promote a better management of them.
Keywords: Gran Chaco, Multi-temporal analysis, Satellite images, Vegetation maps, Woody encroachment.
Introducción
El Gran Chaco de Bolivia constituye una unidad fisiográfica, que es continuación de la llanura beniana y comprende parte de los departamentos de Santa Cruz, Chuquisaca y Tarija en la región oriental y sur oriental del país (Navarro 2002). Gran parte de la vegetación natural del Gran Chaco está relativamente preservada en Bolivia, pero la sobrecarga animal, la agricultura mecanizada y la explotación petrolera presentan amenazas para la integridad de estos ecosistemas a mediano plazo. Asociado al uso de la tierra con fines de producción ganadera está la incidencia del fuego que se utiliza como el tratamiento más barato para la eliminación de los desechos del desmonte transformados para agricultura o ganadería (Zerda 2002). El fuego es también utilizado para favorecer el rebrote de los pastos naturales y usarlos como alimento para el ganado.
Según Navarro (2002), la vegetación original de la zona estudiada, llamada comúnmente Arenales de Yanahigua era bosque chaqueño de arenales (serie de Acacia emilioana y Schinopsis cornuta) en las crestas y laderas de las dunas, cambiando en las depresiones interdunares de sustratos arcillosos mal drenados, a bosque chaqueño de mampuesto mal drenado (serie de Cordia bordasii y Tabebuia nodosa). Actualmente, el paisaje está conformado por un mosaico de etapas seriales o sucesionales en diversos grados de madurez y con diferentes estructuras como respuesta al comportamiento diferencial del fuego según la estructura y composición de la vegetación y las diferencias locales de relieve (Navarro 2002). Las etapas seriales o sucesionales en el Gran Chaco boliviano están caracterizadas mayormente por la invasión de especies arbustivas y semiarbustivas sobre la vegetación herbácea y con gramíneas, relegando así a una superficie cada vez más reducida de pastizales comúnmente llamado pampas chaqueñas (Navarro 2002).
El concepto de sucesión inicialmente desarrollado por Warming (1896) y Cowles (1901) y posteriormente explicado en una teoría de sucesión de vegetación por Clements en 1904 y 1916 (véase Glenn-Lewin et al. 1992) está basado en el establecimiento de una nueva especie debido a la colonización y la posterior modificación del medio ambiente. La teoría Clementsiana basada en el equilibrio ha sido ampliamente criticada (por ejemplo, Gleason 1927, Connell & Slatyer 1977, Westoby et al. 1989, Fernández-Giménez & Allen-Díaz 1999), básicamente porque argumentan que los modelos teóricos basados en el equilibrio y las medidas de manejo de los recursos basados en ellos no han podido predecir el comportamiento de sistemas naturales complejos (Connell & Sousa 1983). Sin embargo, el legado de los primeros conceptos de sucesión desvelan un modelo de facilitación mediante la invasión secuencial y la modificación del hábitat de reemplazo (véase Johnson & Miyanishi 2007). Debido a la modificación del hábitat, se cree que los entornos cambiantes (naturales y/o inducidos por el hombre) tienen profundas implicaciones para la persistencia y estabilidad de ecosistemas (Fryxell et al. 2004).
Aunque existe evidencia de que en las últimas décadas el impacto de las perturbaciones humanas ha aumentado en las sabanas áridas y semiáridas (Archer et al. 1995), hay notablemente poca información sobre los cambios resultantes y las prácticas asociadas de uso de la tierra en biomas subtropicales, especialmente en bosques secos estacionales (Janzen 1988, Redford et al. 1990, Pennington et al. 2000).
Para entender mejor la transformación de las comunidades vegetales en la zona suroeste del Parque Nacional Kaa-Iya del Gran Chaco (PNKI de aquí en más) y brindar información que pudiera ser aplicada a un manejo adecuado de las mismas es que desarrollamos un estudio de los cambios estructurales de la vegetación a través del tiempo y en un área determinada, utilizando imágenes satelitales y herramientas de los sistemas de información geográfica (Killeen1998). La imagen de satélite a diferencia de la fotografía aérea permite su manipulación digital con el fin de mejorar el contraste para resaltar diferentes tipos de superficies y proporcionar un estimativo de las características físicas de estas. También pueden ser utilizadas en el estudio cuantitativo (almacenamiento de datos) geográfico y temático de una región en particular (Lillesand et al. 1994).
El principal objetivo de este estudio fue identificar, cuantificar y comparar las diferentes comunidades (o etapas sucesionales) que conformaron el mosaico de vegetación entre 1975 y 1999. Finalmente esperamos que la información obtenida en este trabajo sea aplicada al momento de estudiar la pérdida de hábitat para especies dependientes de áreas abiertas y promover así un mejor manejo de las mismas.
Métodos
Área de estudio
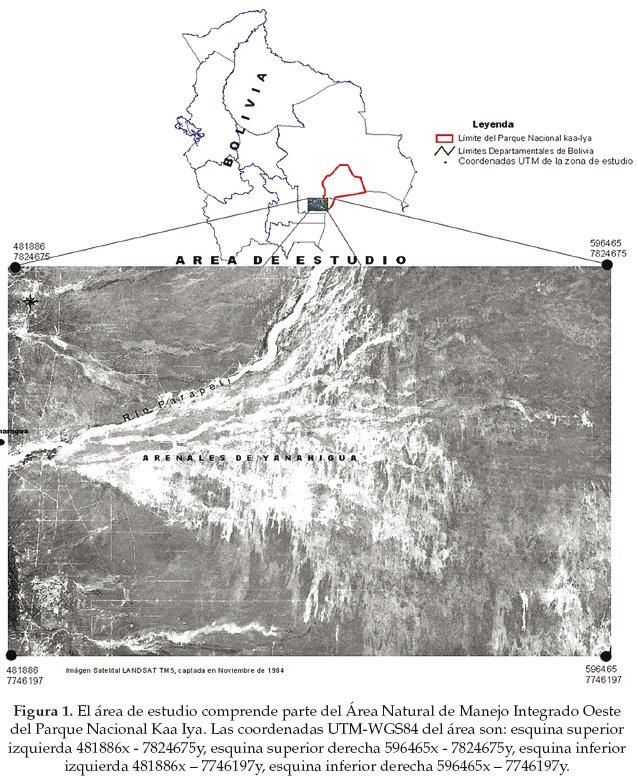
Topográficamente el Chaco boliviano es una llanura, con la presencia de colinas, lomas y pequeñas serranías. Se caracteriza por los escasos ríos, los suelos varían desde arenosos hasta arcillosos en distancias cortas Ramella & Spichger (1989). El área de estudio se encuentra ubicado al sur este del Municipio de Charagua (Provincia Cordillera del Departamento de Santa Cruz). Es un área conocida generalmente como Arenales de Yanahigua que limita al norte con el río Parapetí, al sur con la ruta Transchaco, al este con el Paraguay y al oeste con la vía férrea Yacuiba Santa Cruz. El área que comprende el presente estudio forma parte del Área Natural de Manejo Integrado al oeste del Parque Nacional Kaa Iya. La altitud promedio es de 500 m con las coordenadas UTM-WGS84 que son: esquina superior izquierda 481886x - 7824675y, esquina superior derecha 596465x - 7824675y, esquina inferior izquierda 481886x - 7746197y, esquina inferior derecha 596465x - 7746197y. La superficie total del área de estudio alcanza aproximadamente los 900.000 ha (Fig. 1).
Clasificación de las imágenes satelitales
En el proceso de clasificación de las diferentes comunidades de vegetación seguimos la clasificación de Navarro (2002).
Realizamos la clasificación utilizando imágenes LANDSAT correspondientes a las escenas 230-074 del sensor MSS (1975) y TM (1984, 1999). El siguiente paso fue georeferenciar las imágenes LANDSAT mediante imágenes ortorectificadas (tomando en cuenta puntos estratégicos entre ambas imágenes como: cruce de caminos, ríos, comienzo de una elevación, localidades antropogénicas). Una vez georeferenciadas las distintas imágenes para cada año de estudio, procedimos al levantamiento de datos de campo.
Una vez realizada la toma de datos en campo, procedimos al trabajo de gabinete, para esto utilizamos los puntos de control tomados en el campo de las diferentes unidades de vegetación. Para poder definir la firma espectral de cada unidad de vegetación utilizamos el programa ERDAS® (luego de realizar el mejoramiento, realce y combinación de las bandas, ajustes de los histogramas, contrastes y finalmente la clasificación de las imágenes). Para definir las unidades de vegetación estudiamos las bandas 3, 4 y 5 de las imágenes satelitales correspondientes a los colores rojo, verde y azul, respectivamente.
Una vez definidas las unidades de vegetación y su respectiva firma espectral procedimos a la clasificación no supervisada de las imágenes con 150 clases y una interrelación de 50 puntos entre cada clase, utilizando el modelo de clasificación de ERDAS®, el cual toma en cuenta la firma espectral de cada píxel y permite una clasificación de los píxeles con mayor resolución. Luego de la clasificación en 150 clases, realizamos el sorteo de cada una de las clases para ver en qué tipo de vegetación llegaban a clasificarse.
Finalmente realizamos el análisis multitemporal de cambios de cobertura por sobre posición de imágenes a través del tiempo (para los años 1975, 1984 y 1999).
Resultados
Seis unidades de vegetación fueron evaluadas en el área de estudio que son detalladas a continuación:
a) Pajonal herbazal: Presenta una estructura de sabana herbácea abierta, con árboles y arbustos dispersos, resistentes a las quemas o regenerados a partir de rebrote de tocón. Esta unidad es caracterizada por el predominio de herbáceas y subfrútices pioneras como Alternanthera albida, Aristida mendocina, Eragrostis chiquitanensis, Paspalum chaseanum, Heteropogon melanocarpus, Rhynchosia burkartii, Gomphrena perennis, Sida argentina, Staelia virgata, Turnera krapovickasii, Gaya tarijensis, Mimosa spp., Passiflora foetida y Pappophorum krapovickasii.
b) Matorral: Representa etapas sometidas a mayor frecuencia de fuegos en la zona, donde predominan especies leñosas o subfruticosas de 0.5-1.7 m de altura, con menor cobertura de especies arbóreas y arbustivas. Especies características de esta etapa son Caesalpinia argentina, Lantana grisebachii, Lantana micrantha, Lippia sp, Senna chloroclada y Zexmenia aspilioides. Esta formación típicamente se halla dominada por el acerillo (Lippia sp.), pero existen áreas un poco más evolucionadas sucesionalmente donde abunda el lanza-lanza (Senna chloroclada).
c) Arbustedas de pata de buey: Son agregaciones relativamente menos quemadas que presentan mayor cobertura de arbustos y árboles del bosque original, dominando los primeros, cuya regeneración se ve aparentemente favorecida por el fuego. Entre éstas tenemos a Chloroleucon chacoense, Abutilon herzogianum, Bougainvillea spp., Mimosa castanoclada, Mimosa detinens, Aloysia virgata, Schinopsis cornuta, mezclados con individuos sobrevivientes de Aspidosperma quebracho-blanco, Agonandra excelsa, Sideroxylon obtusifolium y Zizipus mistol entre los más frecuentes.
d) Bosque chaqueño de Arenales: Son bosques caracterizados principalmente por la presencia y abundancia del urundei o sotillo (Schinopsis cornuta) en el nivel de emergentes y Acacia emilioana, Chloroleucon chacoense y Senna chloroclada en el dosel y sotobosque arbustivo. e) Bosque chaqueño medianamente drenado: Son bosques espinosos bajos, densos, con emergentes dispersos, constituidos casi enteramente por especies chaqueñas, sin mayor influencia de la flora de las regiones biogeográficas que contacta.
f) Bosque chaqueño ribereño: Se encuentra sobre sustratos predominantemente arenosos y franco areno-limosos que condicionan la presencia de comunidades vegetales distintas de las del siguiente subsistema, entre ellas el algarrobal freatófilo y los tuscales pioneros. Por ser reservorios de agua permanentes a semipermanentes en una amplia zona de bosques secos en la llanura chaqueña, revisten de importancia fundamental para las poblaciones humanas nativas, así como para especies de fauna silvestre que se concentran aquí durante el largo y marcado periodo seco.
En base a la clasificación anterior, diseñamos tres mapas de vegetación correspondientes al año 1975 (Fig. 2), 1984 (Fig. 3) y 1999 (Fig. 4). La cuantificación de las superficies por cada unidad vegetal se detalla en las tablas 1 (para 1975), tabla 2 (1984) y tabla 3 (1999).
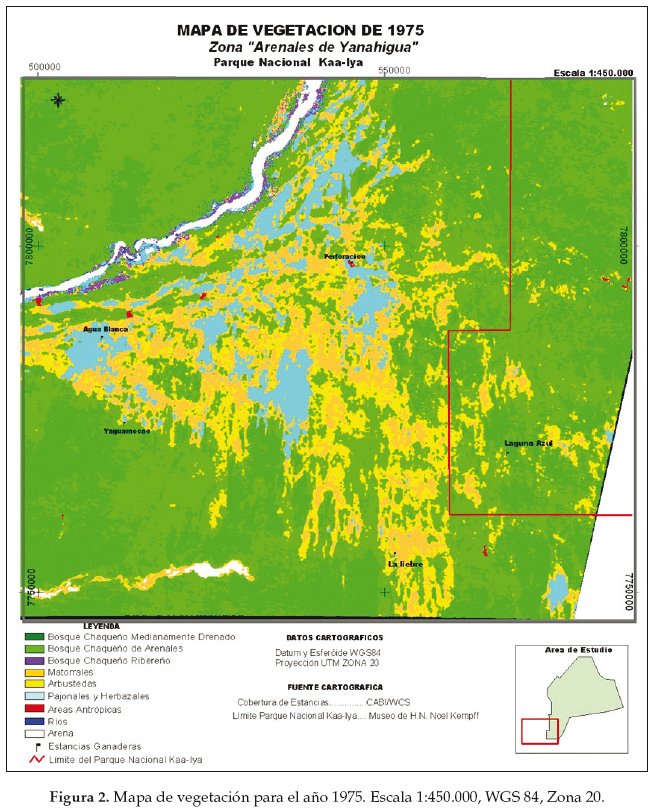
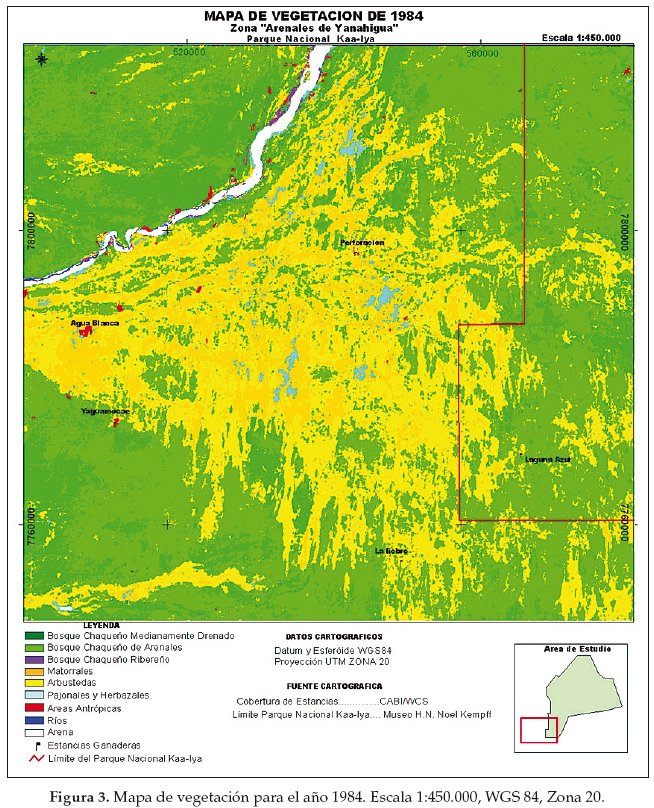
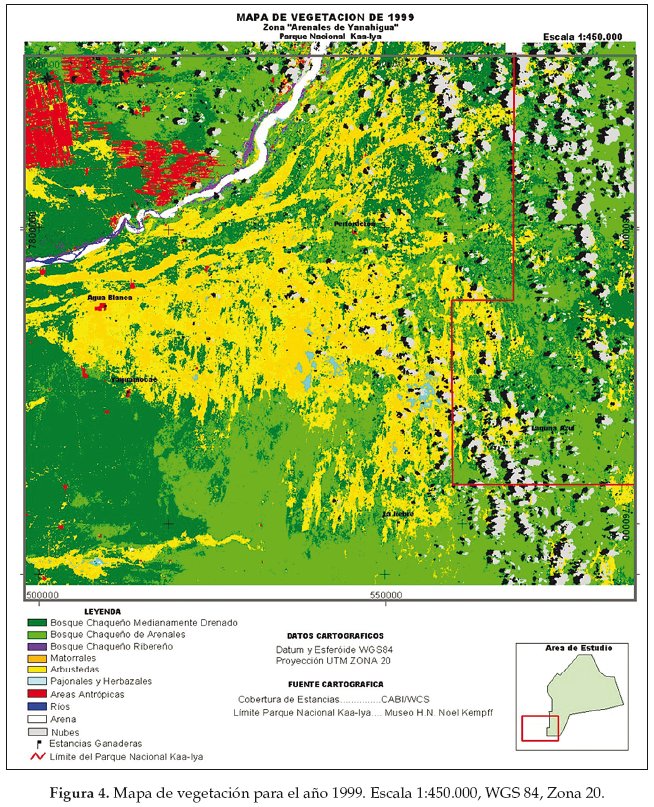
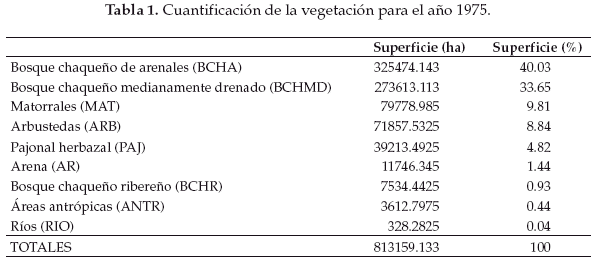
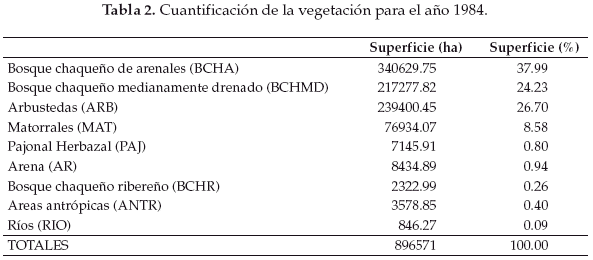
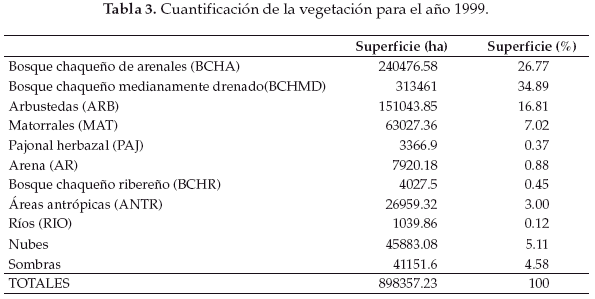
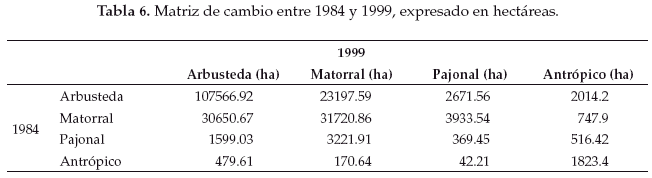
Llamativamente en la comparación cuantitativa entre 1975 y 1999 observamos la mayor retracción (91.5%) de la unidad de vegetación representada por el pajonal herbazal característico de las pampas chaqueñas (Tablas 4-7).



Discusión
Los cambios estructurales en las comunidades de vegetación pueden asociarse con diferentes tipos de perturbaciones (naturales y/o inducidos), que pueden actuar solos o en combinación para modificar y establecer nuevas estructuras (Krebs 2001). Aunque el objetivo de este trabajo no fue determinar los factores que promovieron los cambios estructurales en la vegetación, coincidimos con (Navarro & Fuentes 1999, Navarro 2002, Navarro et al. 2011) que el actual mosaico de unidades de vegetación observado en nuestro estudio es un ejemplo de una sucesión dinámica, tomando en cuenta el establecimiento de las principales etapas de las comunidades de vegetación que preceden al establecimiento de especies leñosas.
A pesar de las discusiones teóricas, no hay consenso en cuanto a cuáles mecanismos son los más importantes para promover la sucesión de la vegetación (Tilman 1985). Sin embargo, consideramos que sería importante mencionar (para tenerlos en cuenta y considerarlos en futuros estudios) los potenciales determinantes que podrían estar causando la invasión de especies leñosas en ambientes áridos y semiáridos incluyendo zonas similares a nuestra área de estudio. El rango de posibles variables reguladoras involucradas hace que sea difícil para cualquier modelo único explicar todo el proceso de sucesión y se ha argumentado que para crear un modelo comprensivo que describa una secuencia sucesional es necesario considerar información adicional como historia natural y fisiología de las plantas tempranas y tardías de la sucesión (Walker & Chapin 1987), lo cual podría ser un tema relevante en futuros estudios. Aparentemente las causas más frecuentemente citadas sobre la invasión leñosa incluyen factores globales tales como el cambio climático que estarían desestabilizando la dinámica de las comunidades de vegetación (Wigley et al. 2010). Entre ellos, los niveles elevados de CO2 podrían tener un efecto positivo significativo en el establecimiento exitoso de plantas leñosas y favorecer así la invasión de arbustos, con lo que se empiezan a cerrar los ecosistemas previamente dominados por gramíneas y herbáceas (Bond & Midgley 2000). Además, los modelos globales de cambio climático predicen que las plantas C3 crecerán más rápido que las C4, como consecuencia de mayores niveles de CO2 en la atmósfera (Ward 2010), aunque un estudio previo de Van Auken (2000) menciona que los niveles elevados de CO2 atmosférico no siempre explican exclusivamente la invasión de arbustos en pastizales semiáridos.
Otros de los mecanismos candidatos es la precipitación anual media (MAP) que es considerado como el principal factor que afecta la distribución y dinámica de la vegetación leñosa (Sankaran 2005). Esto último es basado en el argumento de que cuando la MAP es inferior a 650 mm, la cubierta de vegetación leñosa en las sabanas está restringida por la lluvia. Por el contrario, en las zonas donde el MAP es superior a 650 mm, la lluvia deja de ser un factor limitante y se requieren perturbaciones como el fuego y la herbivoría para evitar el cierre de la cubierta leñosa y, por lo tanto, promover la coexistencia de árboles y pastizales. Otros factores importantes relacionados con la invasión leñosa son las actividades antrópicas, particularmente las altas densidades de ganado y el consecuente sobrepastoreo y los cambios en la frecuencia del fuego utilizados para mantener áreas abiertas para el ganado (Jeltsch et al. 2000, Bond & Midgley 2001, Bond et al. 2003, Sankaran et al. 2004, Bond-Lamberty et al. 2007, Ward 2010). Relacionado con esto último, hay varios estudios que demuestran que el sobrepastoreo y el cambio en los regímenes de fuego pueden transformar la composición de la vegetación de los pastizales semiáridos mediante la sustitución de plantas leñosas palatables y especies herbáceas perennes por especies leñosas y anuales no palatables (Chambers et al. 1999, Janssen et al. 2004), lo cual se observa también en nuestra área de estudio.
Conclusiones
Nuestro estudio permitió obtener por primera vez una evaluación cuantitativa del área de cada uno de los diferentes estados sucesionales en y entre los años estudiados. El mayor cambio observado fue una retracción de alrededor de 91.5% de pastizales/herbazales entre 1975 y 1999 por la arborización de las zonas abiertas.
Aunque no analizamos las causas inmediatas de la retracción y/o invasión de los diferentes estadios sucesionales, sugerimos que los cambios observados mediante las imágenes satelitales se deban probablemente a una combinación de factores que deberían ser estudiados a detalle como: la frecuencia de fuego, la introducción de ganado, la precariedad de la infraestructura para hacer un buen manejo de pasturas para el ganado, la prohibición total del uso de fuego para manejo de pastizales por la cercanía de las estancias al Parque Kaa-Iya.
Finalmente, la modificación del paisaje por la invasión de especies leñosas en áreas abiertas con pastizales nativos sugiere, entre otros aspectos, la pérdida de hábitats para especies dependientes de pastizales y sabanas. Sugerimos que nuestros resultados sean tomados en cuenta para entender las posibles implicancias causadas por la retracción de pampas chaqueñas observada en nuestro estudio sobre la fauna dependiente de estos ecosistemas.
Agradecimientos
Agradecemos a Gregorio Castro, Jorge Segundo y al personal del Parque Kaa-Iya del Gran Chaco por su inestimable apoyo. Este trabajo fue parte de la tesis de licenciatura de CP, con el apoyo de la Sociedad de Conservación de la Vida Silvestre, Gordon y la Fundación Betty Moore dentro del Programa de Conservación de los Andes Amazónicos (AACP) "Conservación de Paisajes Amazónicos". Otros patrocinadores que apoyaron el proyecto fueron: Sheared Earth, Fondo Whitley Fund for Nature, Fundación Kaa-Iya.
Referencias
Archer, S, D.S. Schimel & E.A. Holland. 1995. Mechanisms of shrubland expansion -land use, climate or CO2. Climatic Change 29(1): 91-99.
Bond, W.J. & G.F. Midgley. 2000. A proposed CO2-controlled mechanism of woody plant invasion in grasslands and savannas. Global Change Biology 6(8): 865-869.
Bond-Lamberty, B., S.D. Peckham, D.E. Ahl & S.T. Gower. 2007. Fire as the dominant driver of central Canadian boreal forest carbon balance. Nature 450: 89-97.
Bond, W.J., W. Stock & S. Archibald. 2003. Fire vs. mammal grazers and the nature of zululand savannas. South African Journal of Botany 69(2): 225-226.
Bond, W.J. & J.J. Midgley. 2001. Ecology of sprouting in woody plants: the persistence niche. Trends in Ecology & Evolution 16(1): 45-51.
Chambers, F.M., D. Mauquoy & P.A. Todd. 1999. Recent rise to dominance of Molinia caerulea in environmentally sensitive areas: new perspectives from palaeoecological data. Journal of Applied Ecology 36(5): 719-733.
Cowles, H. C. 1901. The physiographic ecology of Chicago and vicinity. Botanical Gazetter 31: 73-108. [ Links ]
Connell, J.H. & R.O. Slatyer. 1977. Mechanisms of succession in natural communities and their role in community stability and organization. American Naturalist 111(982): 1119-1144.
Connell, J.H. & W.P. Sousa. 1983. On the evidence needed to judge ecological stability or persistence. American Naturalist 121(6): 789-824.
Fryxell, J.M., J.F. Wilmshurst & A R.E. Sinclair. 2004. Predictive models of movement by Serengeti grazers. Ecology 85(9): 2429-2435.
Fernandez-Gimenez, M.E. & B. Allen-Diaz. 1999. Testing a non-equilibrium model of rangeland vegetation dynamics in Mongolia. Journal of Applied Ecology 36(6): 871-885.
Glenn-Lewin, D.C., R.K. Peet & T.T. Veblen (eds.). 1992. Population and community biology series, No. 11. Plant succession: Theory and prediction. Population and Community Biology Series. Chapman and Hall, Nueva York. 352 p.
Gleason, H.A. 1927. Further views on the succession-concept. Ecology 8: 299-326. [ Links ]
Janssen, M.A., J.M. Anderies & B.H. Walker. 2004. Robust strategies for managing rangelands with multiple stable attractors. Journal of Environmental Economics and Management 47(1): 140-162.
Johnson, E. A. & K. Miyanishi (eds.). 2007. Plant disturbance ecology: the process and the response. Academic Press, Oxford. 720 p.
Janzen, D.H. 1988. Tropical ecological and biocultural restoration. Science 239(4837): 243-244. [ Links ]
Jeltsch, F., G.E. Weber & V. Grimm. 2000. Ecological buffering mechanisms in savannas: a unifying theory of long-term tree-grass coexistence. Plant Ecology 150(1-2): 161-171.
Killeen, T. J. 1998. Vegetation and flora of Parque Nacional Noel Kempff Mercado, RAP (Rapid Assessment Program) A biological Assessment of Parque Nacional Noel Kempff Mercado, Bolivia. Technical Reports. Conservation International, Washington DC. 85 p.
Killeen, T.J., E. García & S.G. Beck. 1993. Guía de árboles de Bolivia. Herbario Nacionalde Bolivia, Missouri Botanicakl Garden, La Paz. 966 p.
Krebs, C.J. 2001. Ecology: the experimental analysis of distribution and abundance. Benjamin Cummings, San Francisco, California. 608 p. [ Links ]
Lillesand, T.M., R.W. Kiefer & J. Chipman. 1994. Remote sensing and image interpretation Wiley. John Wiley & Sons, Nueva York. 763 p.
Navarro, G. & A. Fuentes. 1999. Geobotánica y sistemas ecológicos de paisaje en el Gran Chaco de Bolivia. Revista Boliviana de Ecología y Conservación Ambiental 5: 25-50.
Navarro, G., J.A. Molina & S. Vega. 2011. Soil factors determining the change in forests between dry and wet Chacos. Flora 206(2): 136-143.
Navarro, G. 2002. Ecología estructural y dinámica de áreas quemadas en el PN-ANMI Kaa-Iya del Gran Chaco. pp 5-74. En: Guerrero, J. (ed.), Ecología del Fuego en el Parque Nacional y Área Natural de Manejo Integrado Kaa-Iya del Gran Chaco. Proyecto Kaa-Iya, Santa Cruz.
Pennington, R. T., D.E. Prado & C.A. Pendry. 2000. Neotropical seasonally dry forests and Quaternary vegetation changes. Journal of Biogeography 27(2): 261-273.
Ramella, L. & R. Spichiger. 1989. Interpretación preliminar del medio físico y de la vegetación del Chaco Boreal: Contribución al estudio de la flora y de la vegetación del Chaco. Candollea 44(2): 639-680.
Redford, K.H., A. Taber & J.A. Simonetti. 1990. There is more to biodiversity than the tropical rain-forests. Conservation Biology 4(3): 328-330.
Sankaran, M., J. Ratnam & N.P. Hanan. 2004. Tree-grass coexistence in savannas revisited - insights from an examination of assumptions and mechanisms invoked in existing models. Ecology Letters 7(6): 480-490.
Sankaran, M., N.P. Hanan, R.J. Scholes, J. Ratnam, D.J. Augustine, B.S. Cade, J. Gignoux, S.I. Higgins, X. Le Roux, F. Ludwig et al. 2005. Determinants of woody cover in African savannas. Nature 438(7069): 846-849.
Tilman, D. 1985. The resource-ratio hypothesis of plant succession. American Naturalist. 125(6): 827-852. [ Links ]
Van Auken, O. W. 2000. Shrub invasions of North American semiarid grasslands. Annual Review of Ecology and Systematics 31: 197-215. [ Links ]
Walker, L.R. & F.S. Chapin. 1987. Interactions among processes controlling successional change. Oikos 50(1): 131-135.
Warming, J.E.B. 1896. Lehrbuch der ökologischen Pflanzen-geographie. Gebruder Bornträger, Berlín. 412 p. [ Links ]
Ward, D. 2010. A resource ratio model of the effects of changes in CO2 on woody plant invasion. Plant Ecology 209(1): 147-152. [ Links ]
Westoby, M., B. Walker & I. Noy-Meir. 1989. Opportunistic management for rangelands not at equilibrium. Journal of Range Management 42(4): 266-274.
Wigley, B.J., W.J. Bond & M.T. Hoffman. 2010. Thicket expansion in a South African savanna under divergent land use: local vs. global drivers? Global Change Biology 16(3): 964-976.
Zerda, H. 2002. La degradación en los bosques secos del Chaco argentino. Investigación forestal en acción 9(1): 3-5. [ Links ]
Artículo recibido en: 13 de marzo de 2017
Manejado por: Katya Romoleroux
Aceptado en: 13 de abril de 2017

