









 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO 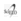 Similares em
SciELO
Similares em
SciELO  uBio
uBio 
 Permalink
PermalinkEcología en Bolivia
versão impressa ISSN 1605-2528versão On-line ISSN 2075-5023
Ecología en Bolivia vol.50 no.1 La Paz abr. 2015
Artículo
Comunidades vegetales de los bofedales de la Cordillera Real (Bolivia) bajo el calentamiento global
Plant communities of high-Andean wetlands of the Cordillera Real (Bolivia) in the face of global warming
Susi Loza Herrera1,2*, Rosa. I. Meneses3 & Fabien Anthelme1,2,3
1Instituto de Ecología, Universidad Mayor San Andrés, Casilla 10077 - Correo Central, La Paz, Bolivia.
*Autor para correspondencia: s.lozaherrera@gmail.com
2Institut de Recherche pour le Développement (IRD), UMR AMAP, Boulevard de la Lironde, TA A-51/PS2, 34398 Montpellier Cedex 5, Francia.
3Museo Nacional de Historia Natural, Herbario Nacional (LPB), Cota Cota, Casilla 8706, La Paz, Bolivia.
Resumen
Los humedales altoandinos (bofedales) son uno de los más amenazados frente al calentamiento global. Dada la estrecha relación entre bofedales y agua, el retroceso glacial podrá causar su contracción. Con tres proxis del cambio climático (área de bofedales, influencia glaciar y altitud) se infirió indirectamente cómo éstos pueden afectar a las comunidades vegetales. Las hipótesis fueron 1) que la reducción del área del bofedal podría afectar negativamente la diversidad de plantas - la influencia glaciar y la altitud podrían afectar ésta relación por su efectos en la heterogeneidad ambiental y la diversidad - y 2) que el retroceso glacial puede afectar indirectamente la diversidad a través de cambios en las especies dominantes. Se midió la diversidad α y β aditiva de plantas en 20 bofedales (>4.400 m) de la Cordillera Real (Bolivia). Se encontraron 63 especies (riqueza especifica: 5-22/cuadrante 1 m2). Variaciones del área e influencia glacial no se relacionaron con cambios en la vegetación, pero la diversidad fue menor a mayor altitud. Las hojas de Oxychloe andina son más resistentes al estrés ambiental por su mayor masa foliar seca, a diferencia de Distichia spp., indicando que los bofedales dominados por O. andina son más secos. Además tienen más especies que también se pueden encontrar en las laderas. Mientras, las comunidades de Distichia spp. son menos tolerantes al déficit hídrico. Si el retroceso glacial reducirá el agua en las próximas décadas, el bofedal de O. andina podrá ser más abundante a expensas de los de Distichia spp., generando desafíos para la conservación.
Palabras clave: Bofedal, Calentamiento climático, Distichia muscoides, Oxychloe andina, Retroceso glacial.
Abstract
High-Andean wetlands (bofedales) are one of the most threatened ecosystems in the face of global warming. Given the close relationship between bofedales and water, it is expected that glacial retreat will cause their gradual shrinkage. With three proxies of climate change (bofedal area, glacier influence and elevation) we inferred how these changes may affect plant communities. The hypotheses were 1) that loss in the area of bofedales should reduce plant diversity - the glacier influence and elevation could affect this relationship through effect on environmental heterogeneity and diversity - and 2) glacial retreat may indirectly affect diversity through changes in dominant species. We measured α and β additive diversity of plants in 20 bofedales (>4.400 m) in the Cordillera Real (Bolivia). Sixty three species were found (species richness: 5-22 /plot 1 m2). Variations in bofedales área and glacial influence weren't correlated with vegetation changes whereas diversity reduced at higher elevation. In reference to our second hypothesis, leaves of Oxychloe andina were shown more resistant to drought stress because of its higher leaf dry mass content (LDMC) than the other cushions, indicating that the bofedales dominated by O. andina may be drier and the communities are dominated by species that also can be found in drier environments. In contrast, communities of Distichia spp. are less tolerant to water déficit. If glacial retreat reduces water availability in the coming decades, the bofedales of O. andina may be more abundant at the expense of Distichia spp. creating challenges for the biodiversity conservation.
Key-words: High Andean peatlands, Distichia muscoides, Glader retreat, Global warming, Oxychloe andina.
Introducción
Los bofedales o jok'os (nombre aymara) son un tipo particular de humedales de turbera, característicos de la zona altoandina y puneña dentro de los Andes subtropicales y tropicales, situados a elevaciones entre 3.200-5.000 m (Estenssoro 1991, Squeo et al. 2006). Se caracterizan por estar formados por plantas en forma de cojines abombados a planos (Ostria 1987, Estenssoro 1991). En la Cordillera Real (Bolivia), los cojines están representados principalmente por Distichia muscoides y Oxychloe andina, mientras que Distichia filamentosa, Plantago tubulosa y Phylloscirpus deserticola pueden ser dominantes sólo localmente (Meneses 2012). Hada el sur de los Andes Centrales, se puede encontrar D. muscoides pero dominan O. andina y Patosia clandestina (Squeo et al. 2006, Ruthsatz 2012).
En medio de esta matriz de vegetación emergen y/o atraviesan cuerpos de agua (pozas y arroyos) (Gonzales et al. 2014).
Los bofedales cumplen importantes funciones ecológicas y socioeconómicas. En comparación a otras formaciones vegetales típicas de alta montaña, poseen una gran diversidad biológica tanto a nivel de riqueza especifica como de endemismo (plantas, aves, anfibios, peces, microcrustáceos (Telleria et al. 2006, Coronel et al. 2007, Meneses 2012, Flores 2013, Anthelme et al. 2014, Maldonado 2014). Ayudan a controlar y regular la corriente de agua, reduciendo la erosión que puede disminuir la calidad del agua (Alzérreca et al. 2001, Coronel et al. 2007, Squeo et al. 2006, Benavides et al. 2013). Los bofedales además pueden almacenar carbono orgánico (Segnini et al. 2010) y tienen la capacidad de regular las emisiones de CO2 a través del secuestro de carbono atmosférico (Buytaert et al. 2011). En efecto, 1 m2 de O. andina secuestra en un año la misma cantidad de CO2 que al menos 10 m2 de Sphagnum (briofito que forma las turberas de la zona templada; Earle et al. 2003).
En términos socioeconómicos, los bofedales son importantes porque proveen forraje rico en proteínas y nutrientes, muy apetecido por los camélidos sudamericanos domesticados (llamas, alpacas; Pacheco 1998) y silvestres (vicuñas). Un estudio con collares GPS mostró que, en época seca, las vicuñas pasan hasta seis horas por día en los bofedales de Sajama (Javier Cañarí, datos no publicados, proyecto BIO-THAW). Los bofedales también poseen alta capacidad para almacenar agua, funcionan como esponjas que liberan el agua cuando las precipitaciones reducen drásticamente en época seca (Squeo et al. 2006, Benavides et al. 2013, Zeballos 2013). Para optimizar estas funciones ecosistémicas, las comunidades locales utilizan el riego para ampliar los bofedales (Alzérreca et al. 2001, Verzijil & Guerrero 2013, Villarroel et al. 2014) enfatizando su rol clave en la conservación de estos humedales.
Existen varios aspectos que ponen en riesgo todas éstas funciones ecosistémicas que cumplen los bofedales. En primer lugar, como los bofedales se localizan dentro de una matriz seca (insularidad continental) son susceptibles a la fragmentación (Anthelme et al. 2014). Al mismo tiempo, enfrentan varias amenazas como la extracción de turba (Estenssoro 1991), actividad minera (Meneses 2012), la canalización y desvío del agua (Pacheco 1998), pastoreo de ganado mayor y menor que dañan fuertemente los bofedales al no dejar regenerar a las plantas (Alzérreca & Luna 2001, Meneses 2012,); así como la sobrecarga de camélidos (Alzérreca et al. 2001, Squeo et al. 2006).
Planteamos la hipótesis de que el cambio climático podría ser otra amenaza importante para la conservación de los bofedales a largo plazo. De hecho, la temperatura promedio en los Andes ascendió 0.1°C por década en los últimos 60 años (IPCC 2013), causando el retroceso acelerado de los glaciares tropicales andinos, con una pérdida del 40% de la masa glacial en los últimos 40 años (Rabatel et al. 2013). Por la aceleración del calentamiento, esta tendencia podría aumentar en las próximas décadas, particularmente en latitudes bajas y a mayor altitud (Bradley et al. 2006). Esta pérdida de volumen y masa de hielo se podría traducir en un aumento significativo de la escorrentía anual solo durante unos pocos años y posterior descenso. Después de un umbral crítico (dependiendo del tamaño del glaciar), la descarga anual descenderá hasta el final de la influencia del glacial sobre el caudal de salida (Milner et al. 2009, Baraer et al. 2012).
Dada la estrecha relación entre bofedales y agua (Earle et al. 2003, Squeo et al. 2006, Anderson et al. 2011, Ruthsatz 2012) se espera que el retroceso glacial afecte negativamente estos humedales, causando su paulatina contracción por desecamiento (Earle et al. 2003, Chimner & Karberg 2008). Esta relación fue observada en un análisis multitemporal de 401 humedales (vegas y bofedales) de la Cordillera Real, donde se detectó el 50% de tendencia del retroceso de estos humedales (entre 1984 y 2009, Zeballos 2013).
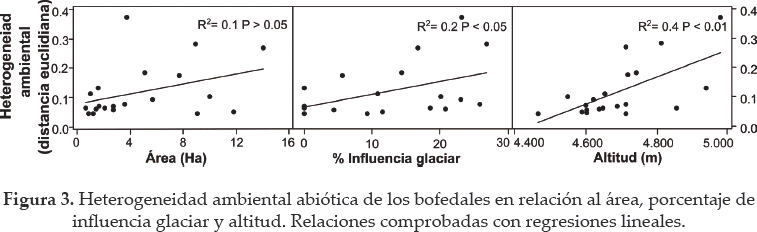
Siguiendo la lógica de reemplazar espacio por tiempo, se utilizaron tres proxis del cambio climático - área de los bofedales, influencia glaciar y altitud - para inferir de forma indirecta como éstos cambios pueden afectar a las comunidades vegetales (Meneses et al. 2014). Consecuentemente, se eligieron bofedales de modo que representen un gradiente de áreas superficies (0-14 ha), diferente porcentaje de influencia glaciar (0-27%) y diferente altitud (4.445-5.045 m).
Nuestra primera hipótesis de trabajo es que la reducción del área de los bofedales puede afectar negativamente su diversidad vegetal (Preston 1962, MacArthur & Wilson 1963) porque en áreas más grandes existe mayor heterogeneidad ambiental, posibilitando mayor diversidad de hábitats y nichos para las especies (Palmer 2007). Dentro de esta hipótesis, tomamos en cuenta que la influencia glaciar podría afectar ésta relación a través de su efecto positivo en la turbidez del agua que estimula mayor heterogeneidad ambiental (Quenta 2013) Asimismo, esperamos que la altitud podría influir sobre la relación área-diversidad porque la diversidad y altitud están negativamente correlacionadas en ambientes altoandinos (Sklenář & Jørgensen 1999).
La segunda hipótesis es que la variación en la disponibilidad de agua en los bofedales bajo un escenario de calentamiento podría afectar la diversidad de plantas a través de cambios en la abundancia de las plantas dominantes (Distichia muscoides, D. filamentosa, O. andina, Plantago tubulosa y Phylloscirpus deserticola). Pues, en ambientes estresados las interacciones positivas entre plantas juegan un papel importante en la estructuración y dinámica de las comunidades de plantas (Callaway 1995, Cavieres et al. 2014).
Métodos
Área de estudio
Los bofedales en estudio se encuentran en cinco cuencas glaciales de la Cordillera Real (Departamento de La Paz, Bolivia), entre 4.445-5.045 m, incluidos desde el valle glacial de Hichu Khota (16°3'44.78"S, 68°18'3.86"W) hasta la zona de influencia del nevado Huayna Potosí (16°16'55.70"S, 68°6'37.36"W; Fig. 1). Esta área tiene alta relevancia socio-económica principalmente para la provisión de agua a las ciudades de La Paz y El Alto (Ramírez 2008) y porque proveen forraje durante todo el año para la ganadería camélida (Alzérreca et al. 2001). Además presenta características ambientales relativamente homogéneas, permitiendo revelar patrones comunes (Soruco et al. 2005). Las precipitaciones en época húmeda (diciembre a marzo) llegan a 410.4 mm y el resto del año es seco (184 mm) (2000-2009 en la Estación Tuni, Empresa Pública Social de Agua y Saneamiento, EPSAS). La temperatura media en época húmeda es 6.4°C y en época seca 4.5°C (Soruco et al. 2005).
Diseño de estudio
Se eligieron 20 bofedales por encima de los 4.400 m de elevación para evitar la influencia de otros disturbios de origen antropogénico, como el sobrepastoreo y minería que se encuentran frecuentemente debajo de esta altitud (Ostria 1987). Asimismo, para evitar efectos de variables de confusión solo se trabajó en bofedales planos dentro de la categorización propuesta por Squeo et al. (2006). Todos los bofedales se encuentran en la ladera oeste de la Cordillera Real a excepción de tres sitios en Huayna Potosí (HP5, HP6, HP7) y uno en Hichu Khota (HK1).
Para detectar el efecto del área, de la influencia glaciar y de las variaciones altitudinales -tres indicios indirectos de los efectos del cambio climático- se estudiaron bofedales con diferentes áreas (entre 0.5 y 14 ha), con diferente porcentaje de influencia glaciar (entre 0 y 27%) (más detalles en la sección recolección de datos en laboratorio) y con diferentes altitudes (entre 4.989 y 4.459 m; Meneses et al. 2014).
En cada bofedal, se evaluaron 10 cuadrantes de 1 m2 (N = 200). Se eligieron los cuadrantes con al menos 50% de cobertura de las especies estructurantes representativas del área de estudio: Distichia muscoides, D. filamentosa, Oxychloe andina, Plantago tubulosa y Phylloscirpus deserticola.
Recolección de datos en campo
Diversidad y tolerancia al estrés ambiental
En este trabajo se consideraron dos niveles de diversidad: alfa (a través de la riqueza específica) y diversidad beta aditiva, que indica el recambio de especies y se calcula en base a la riqueza total y a la riqueza promedio por bofedal. Así, en cada cuadrante de 1 m2, se registraron las especies vegetales presentes y su porcentaje de cobertura.
Para estimar el estrés ambiental que pueden tolerar las comunidades vegetales de bofedales, se midió el contenido de masa seca en hojas de los diferentes cojines dominantes (LDMC; ver Meneses et al. 2014 para más detalles). El LDMC se puede utilizar como un indicio de tolerancia al estrés ambiental porque se correlaciona negativamente con el crecimiento potencial relativo de las plantas (Cornelissen et al. 2003) y es recomendado para usar como proxy de la fertilidad de los suelos (Hodgson et al. 2011). Así, los valores altos de LDMC se relacionan con mayor tolerancia al estrés ambiental.
Medidas ambientales
Se midieron variables ambientales en el campo para estudiar como las variaciones del área, de la influencia glaciar y de altitud cambiaron los parámetros locales. Primeramente, se cavaron pequeños pozos (profundidad aproximada de 20 cm) adyacente a cada cuadrante evaluado, en medio de los tepes de vegetación, donde se midió in situ con un equipo multiparamétrico (TESTR35 marca OAKTON, Malasia) la temperatura, el pH y la conductividad eléctrica del agua en los mismos sitios de evaluación de plantas. Luego, en cada pozo se recolectó 500 ml de agua en frascos plásticos estériles para evaluar el contenido de nitrógeno, fósforo y potasio (nutrientes N, P, K). Posteriormente, se obtuvieron muestras del sustrato presente debajo de cada especie dominante para su posterior análisis de NPK y porcentaje de materia orgánica.
Posteriormente, dentro de cada cuadrante se midió la compactación (kPa) con un compactómetro (Agratronix, Streetboro modelo OH 44241, USA) a través de cinco mediciones distribuidas uniformemente en cada uno.
Recolección de datos en laboratorio LDMC
Inmediatamente al llegar al laboratorio se seleccionaron 20 hojas enteras jóvenes (fotosintéticamente más productivas) de cada especie colectada. Posteriormente las hojas fueron guardadas en sobres de papel sabana etiquetados para secarlos en el horno a 65°C durante 72 horas hasta que las hojas perdieron toda el agua. Finalmente se registró el peso seco de las hojas.
Medidas ambientales
Se determinó la cantidad de agua dentro del sustrato (suelo) (% humedad) mediante la medición del peso fresco y peso seco (luego de secarlas en horno a 65°C por 72 horas) a través de la fórmula: Porcentaje de humedad=[(Ph-Ps) Ps-1] 100, donde Ph es el peso del suelo húmedo y Ps, el peso del suelo seco (g).
La determinación de nutrientes NPK y materia orgánica fue realizada por el Laboratorio de Calidad Ambiental (LCA, del Instituto de Ecología-Universidad Mayor de San Andrés en La Paz). Las muestras de agua extraídas de los pequeños pozos cavados se analizaron directamente. Mientras, para las muestras de hojas de las especies dominantes primeramente se seleccionaron hojas de los tepes extraídos en campo, se secaron en horno a 65°C durante 72 horas y posteriormente se molieron en mortero. Las muestras de sustratos también se secaron y molieron bajo las mismas condiciones. Posteriormente en el LCA, el nitrógeno total se determinó por el método estándar de Kjeldahl, el fósforo se cuantificó por el método del molibdato. Para determinar el potasio total se utilizó espectrofotometría de absorción atómica, previa extracción del potasio con ácido.
La conductividad eléctrica es sensible a los cambios de temperatura. Para evitar sesgos en las mediciones, se aplicó un factor de corrección (FC) = 0.0408 (25°C-X) + 0.7944, donde: X es la temperatura de la medición de conductividad in situ. Posteriormente para calcular la conductividad estandarizada a 25°C se aplicó la fórmula: (FC) (CE), donde FC es el factor de corrección y CE la conductividad eléctrica medida in situ.
El área y el porcentaje de influencia glaciar en los bofedales se obtuvieron del trabajo de Quenta (2013), que calculó la influencia glaciar a través del porcentaje de cobertura glaciar en la cuenca, para lo cual inicialmente se delimitaron los ríos de cada valle con un Modelo Digital de Elevación (imagen DEM, de 30 m de altura) y a partir del punto GPS de cada bofedal con fines metodológicos se delimitaron 20 microcuencas (una por cada bofedal). La delimitación de ríos y cuencas se realizó en el programa SAGA (System Automated Geoscientific Analyses). Posteriormente, cada cuenca se intersectó con la cobertura glaciar en el programa ArcMap, donde se calculó el área de cada cuenca y la cobertura glaciar con la herramienta de cálculo de área, finalmente se calculó el índice de influencia glaciar, donde el porcentaje de glaciar incluida en el área de cada cuenca es el porcentaje de influencia glaciar por bofedal (para más detalles ver Gonzales et al. 2014).
Análisis de datos
La diversidad de plantas se cuantificó a través de la riqueza específica (índice de diversidad α) y mediante la diversidad β aditiva que indica el recambio de especies y se aplicó la fórmula: β= Υ-α (Veech et al. 2002), donde Υ es la riqueza de plantas a nivel de todo el bofedal y a es la riqueza promedio de los 10 cuadrantes por bofedal.
Para determinar la heterogeneidad ambiental abiótica se calculó la distancia euclidiana (Quinn & Keough 2002) con lo que se cuantificó cuan disímiles o diferentes son dos cuadrantes en términos de las variables ambientales medidas. La distancia euclidiana se calculó en base a los siguientes parámetros medidos en cada cuadrante: pH, conductividad eléctrica, nitrógeno, fósforo y potasio total del sustrato y del agua y, porcentaje de humedad. Su fórmula es:
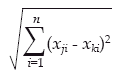
Donde, Xji = el valor de la variable ambiental i en el cuadrante j y Xki = el valor de la variable ambiental i en el cuadrante k.
Posteriormente, se promediaron las distancias euclidianas entre pares de cuadrantes para tener una medida de heterogeneidad por bofedal. Este índice puede tomar valores desde cero y no tiene límite superior, entonces puede tomar valores muy grandes; por tanto se estandarizaron los datos dividiendo el valor obtenido entre el máximo para tener valores de 0-1, donde los valores cercanos a uno indican mayor heterogeneidad (Quenta 2013). Posteriormente, en el programa Minitab versión 15 se corrieron regresiones lineales utilizando como variables dependientes la riqueza, la diversidad beta aditiva y el promedio de las distancias euclidianas (heterogeneidad ambiental), y como variables independientes el área de los bofedales, el porcentaje de influencia glaciar y la altitud.
Para determinar el efecto de las especies vegetales dominantes (Distichia muscoides, D. filamentosa, Oxychloe andina, Plantago tubulosa y Phylloscirpus deserticola) sobre la diversidad de plantas se aplicó un análisis de correspondencia canónica (CCA) en el programa CANOCO versión 4.5, utilizando como única variable explicativa categórica a la especie dominante. La significancia estadística del modelo generado se analizó mediante la prueba de Monte Carlo con 999 permutaciones (Leps & Smilauer 2003). Posteriormente estos resultados se corroboraron con pruebas de ANOVA de una vía. Con los datos fisicoquímicos, la compactación, coeficiente de variación de la compactación, nutrientes en hojas y agua se corrieron ANOVAS de una vía para determinar variaciones significativas, posteriormente los resultados se corroboraron a través de pruebas de Tukey. En los ANOVAS se excluyeron las muestras donde dominaban P. tubulosa y P. deserticola porque su tamaño de muestra era bajo (N<10).
Resultados
En total, se encontraron 63 especies de plantas vasculares, distribuidas en 17 familias, de las cuales las más abundantes fueron Asteraceae y Poaceae (12 y 14 especies, respectivamente; Anexo 1). La riqueza varió desde 5 hasta 22 especies por cuadrante de 1 m2. En todos los cuadrantes evaluados la cobertura relativa de la vegetación siempre fue superior al 90%.
Efectos del área, influencia glaciar y altitud
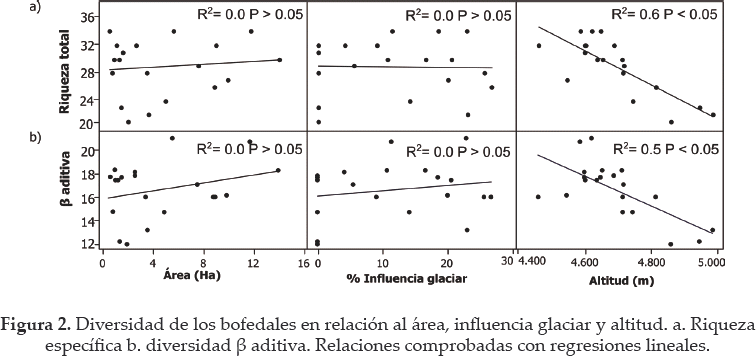
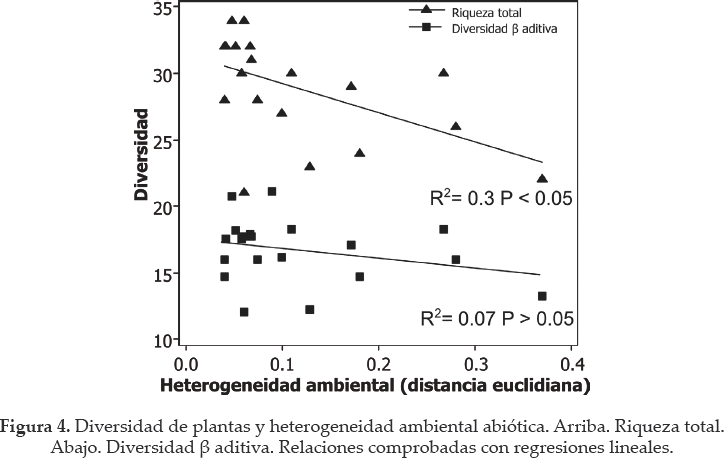
Los análisis de regresión no mostraron relación entre la riqueza de plantas y el área de los bofedales (R2= 0.0; P > 0.05) ni entre la riqueza y la influencia glaciar (R2= 0.0; P > 0.05) (Fig. 2a). Sin embargo, la riqueza y diversidad beta redujeron significativamente a medida que los bofedales estaban a mayor altitud (R2 = 0.6; P < 0.05 y R2= 0.5; P < 0.05, respectivamente, Fig. 2b). La heterogeneidad ambiental tampoco mostró relación significativa con el área (R2 = 0.1; P > 0.05) pero si se relacionó positivamente con la influencia glaciar y la altitud (Fig. 3). Se encontró que la riqueza específica redujo en bofedales más heterogéneos (Fig. 4).
Efectos de la especie dominantes sobre la diversidad de plantas
El análisis CCA generó un modelo significativo de composición de plantas en relación con la especie dominante (inercia total de los ejes canónicos: 0.72, P < 0.01; Fig. 5).
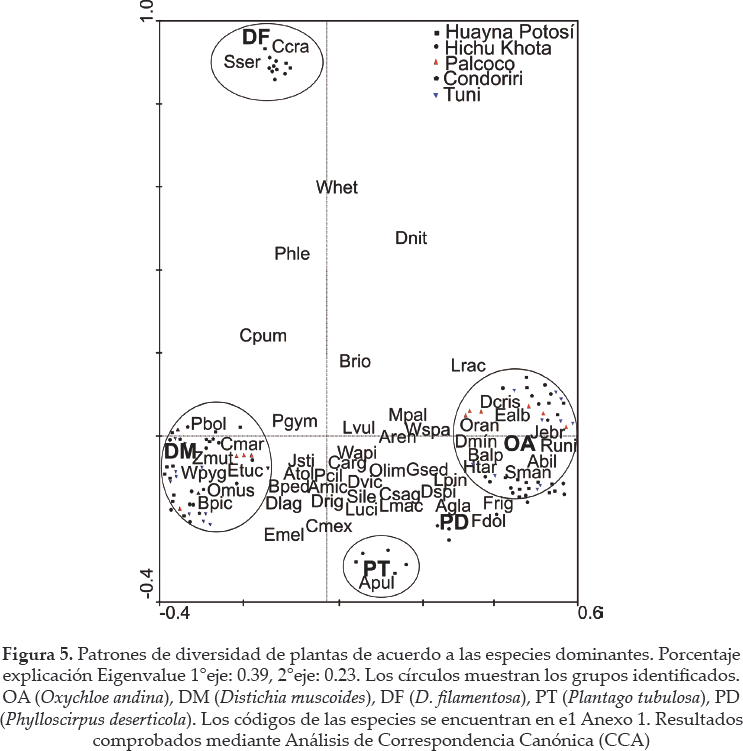
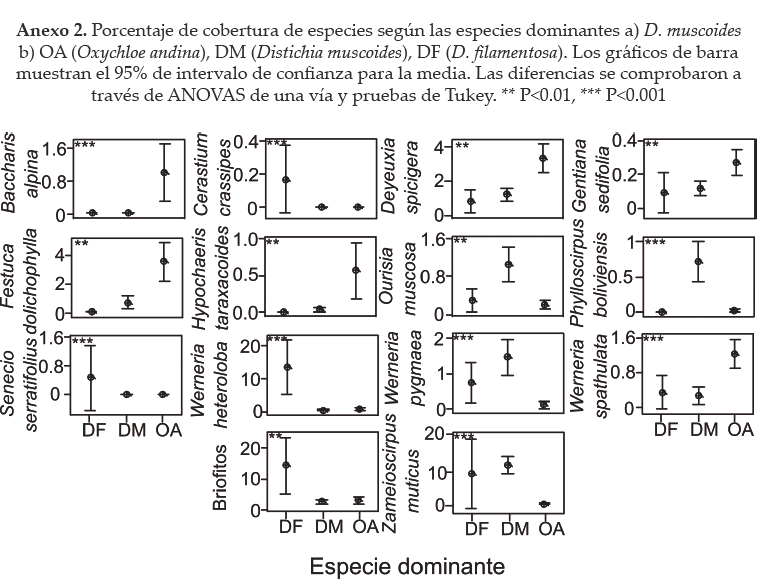
El CCA mostró que cuatro especies: Werneria pygmaea (Wpyg), Ourisia muscosa (Omus), Phylloscirpus boliviensis (Pbol) y Zameioscirpus muticus (Zmut)) son características de los bofedales dominados por D. muscoides (DM). Siete especies son características de los sitios dominados por O. andina (OA): Werneria spathulata(Wspa), Hypochaeris taraxacoides (Htar), Gentiana sedifolia (Gsed), Deyeuxia specigera (Dspi), Festuca dolicophylla (Fdol), F. rigescens (Frig) y Baccharis alpina (Balp). Mientras, en los sitios dominados por D. filamentosa (DF) sólo tres especies parecen características: Cerastium cf. crassipes (Cera) Senecio serratifolius (Sserr) y Werneria heteroloba (Whet). Estos resultados se corroboraron con pruebas de ANO VA (ver Anexo 2).
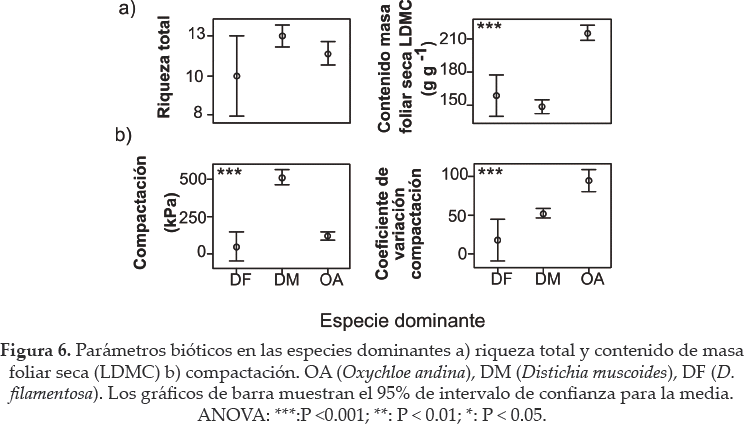
Entre otros parámetros bióticos, la riqueza especifica no varió significativamente entre las especies dominantes (ANOVA: P > 0.05) (Fig. 6a). El LDMC fue significativamente mayor en los cojines de O. andina (OA) a comparación de Distichia spp. (ANOVA: P < 0.001; OA: 215 ± 31.5 g g1; DM: 147.5 ± 31.3 g g1; DF: 157 ± 27.7 g g1) (Fig. 6a). Los cojines de D. muscoides (DM) fueron netamente más compactos que los de D. filamentosa (DF) y O. andina (OA) (ANOVA P < 0.001; OA: 117.5 ± 122.4 kPa; DM: 517.2 ± 259.2 kPa; DF: 46 ± 154.9 kPa), mientras los cojines de O. andina mostraron una variabilidad de compactación mayor al resto de los otros cojines (Fig. 6b).
De las variables abióticas, siete cambiaron significativamente entre especies dominantes (Fig. 7). Destacan, el mayor contenido de nutrientes NPK en hojas de Distichia spp. a comparación de O. andina (OA): fósforo total (ANOVA P < 0.01; OA: 2.941,7 ± 804,6 mg/1; DM: 3.707 ±1.261 mg/1; DF: 3.600 ± 953 mg/1), nitrógeno total (ANOVA P < 0.001, OA: 1.9 ± 0.3 mg/1; DM: 2.5 ±0.5 mg/1; DF: 2.6 ± 0.3 mg/1) (Fig. 7d) y potasio total (ANOVA P < 0.01; OA: 31.530 ± 12.965 mg/1; DM: 42.506 ± 12.630 mg/1; DF: 40.167 ± 3.339 mg/1) (Fig. 7d).
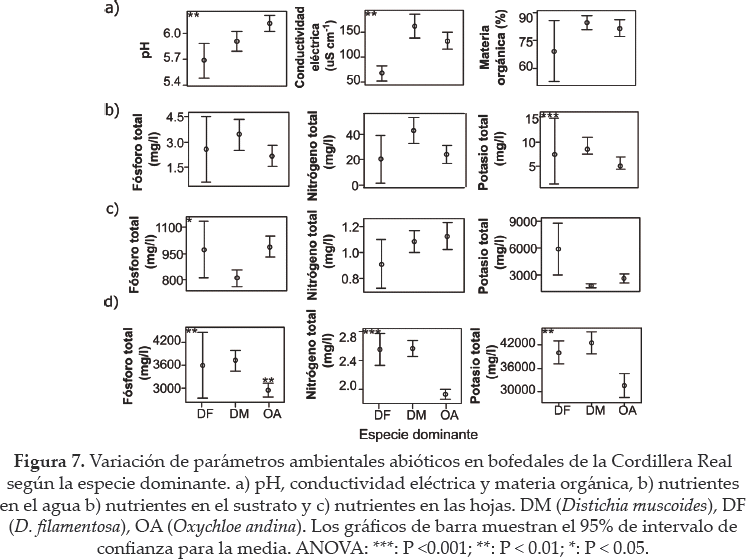
Discusión
En contraste con la primera hipótesis de nuestro trabajo y con la teoría de las islas (Mac Arthur & Wilson 1963) no hay relación entre el área y la diversidad de especies de plantas. En concordancia con este resultado, tampoco se encontró que los bofedales más grandes son los abióticamente más heterogéneos. Contrario a lo esperado, la riqueza de especies redujo en los sitios más heterogéneos. Dentro del posible escenario de reducción del área de los bofedales como consecuencia del retroceso glaciar y la falta de agua disponible (Zeballos 2013), el área no parece tener influencia directa sobre la diversidad de las plantas. La ausencia de la relación riqueza-área se podría explicar por el hecho que todos los bofedales generan su propio sustrato orgánico, a veces de varios metros de materia orgánica formada durante varios siglos (Benavides et al. 2013). Esta acumulación puede causar la homogeneización de los parámetros ambientales, disminuyendo así la diversidad beta o recambio de especies (Kikvidze et al. en prensa). De acuerdo con esta interpretación, los bofedales más jóvenes, i.e. los que no han logrado totalmente esta homogenización ambiental, tendrían menor riqueza específica, como se mostró en este trabajo con los bofedales ubicados a mayor altitud. Igualmente, los que están más influenciados por la proximidad de los glaciares son probablemente más jóvenes y más heterogéneos en términos abióticos porque el glaciar aporta agua con mayor turbidez al sistema, apoyando los resultados de Quenta (2013).
La influencia glaciar no se limita a estimular mayor heterogeneidad. Al contrario, su aporte hídrico es importante principalmente en época seca (que generalmente dura cinco meses en la región de estudio, sin embargo en los últimos años se extiende hasta ocho meses, registro de 1995-2008 en la estación de COBEE). De este modo, los flujos sub-superficiales provenientes del deshielo glaciar permiten mantener bofedales vigorosos (Verzijl & Quispe 2013). Consecuentemente, bajo los escenarios actuales de retroceso glaciar, aumentará la vulnerabilidad de los bofedales por su mayor dependencia de las precipitaciones (Zeballos 2013).
En cambio, los resultados apoyan la segunda hipótesis planteada, de que el retroceso glaciar puede afectar indirectamente la diversidad vegetal a través de cambios en las especies dominantes. De modo interesante, las comunidades características del cojín Oxychloe andina son dominadas por especies que también se pueden encontrar en las laderas de los bofedales, en ambientes más secos, como Festuca dolicophylla, F. rigescens, Baccharis alpina, Deyeuxia spicigera var. spicigera y D. mínima (Ruthsatz 2012). En contraste, las especies características del cojín Distichia muscoides son principalmente típicas del interior de los bofedales, y no tolerantes a la falta de agua dentro del suelo durante época seca. Es el caso de Werneria pygmaea, Ourisia muscosa, Phylloscirpus boliviensis y Zameioscirpus muticus (Meneses 2012). Así, los ambientes dominados por O. andina revelan ser más secos que los dominados por D. muscoides, lo que se confirma con valores de LDMC superiores en O. andina que en D. muscoides. Estos resultados están en concordancia con la literatura existente sugiriendo que O. andina es más tolerante al estrés ambiental, como alta salinidad y sequía (Ruhtsatz 2012) o variación de contenido de agua (Ostria 1987, Meneses 1997). Por su carácter estrés tolerante, esperamos que O. andina también soportaría mejor la presión por pastoreo, al contrario D. muscoides es más susceptible a ser sobrepastoreada por el ganado porque es más nutritiva que O andina (Fig. 7).
Bajo el escenario de incremento de la sequedad dentro de los bofedales, como consecuencia de la disminución de los glaciares, se espera que el calentamiento global previsto en futuras décadas pueda cambiar la dominancia de cojines, desde D. muscoides hasta O. andina.
Estos cambios pueden tener efectos negativos importantes en términos de conservación de la biodiversidad porque las comunidades asociadas específicamente a Distichia spp., que conforman el "corazón" del bofedal y no se encuentran en las laderas, estarían directamente amenazadas. Al contrario, muchas especies asociadas a O. andina se encuentran en las laderas. Entonces, se espera que la contracción del área de los bofedales tenga menor impacto sobre las comunidades de O. andina. Sin embargo, tanto Distichia muscoides, D. filamentosa y Oxychloe andina, están categorizadas como EN PELIGRO a nivel nacional (MMAyA 2012) y las tres especies cumplen funciones esenciales en el bofedal, por tanto requieren ser protegidas para mantener estos humedales.
Conclusiones
Se mostró que la diversidad en plantas de los bofedales altoandinos de la Cordillera Real de Bolivia no estaría afectada directamente por la probable reducción del área como consecuencia del calentamiento global. Sin embargo, los efectos de estos cambios podrían alterar las comunidades vegetales indirectamente, a través de cambios de dominancia de las plantas en cojín (Juncaceae) que estructuran los bofedales, desde O. andina hasta D. muscoides. Dado que éstas dos especies dominantes participan activamente en el funcionamiento/productividad de los bofedales de la Cordillera Real, atributos que se traducen en agua potable y forraje que aprovechan las personas. Sequías más pronunciadas en el contexto de retroceso glaciar podrían llegar a comunidades dominadas por gramíneas (ej. Aciachne pulvinata, Festuca spp.) y especies como Phylloscirpus deserticola, que no tienen la misma capacidad de almacenar agua y formar suelos orgánicos. El paso siguiente seria cuantificar con precisión cuánta productividad y servicios par a el ser humano se tendrá cuando O. andina y Distichia spp. sean reemplazadas por éstas especies.
Agradecimientos
Agradecemos a Ariel Lliully y Arely Palabral por su ayuda en el trabajo de campo e identificación de plantas, a Julieta Acho por su ayuda en el trabajo de laboratorio, a Jaime Chincheros por compartir su conocimiento sobre los nutrientes en el suelo y plantas, a Estefanía Quenta por proporcionar datos del área e influencia glaciar y a Martin Kraemer por la elaboración del mapa del área de estudio.
Referencias
Alzérreca, H., G. Prieto, J. Laura, D. Luna & S. Laguna. 2001. Características y distribución de los bofedales en el ámbito boliviano. Informe final no publicado. Autoridad Binacional del Lago Titicaca (ALT) y Programa de las Naciones Unidas para el Desarrollo (PNUD), LaPaz. 190 p.
Alzérreca, H & D. Luna. 2001. Manual del ganadero para el manejo de bofedales. Asociación Integral de Ganaderos de Camélidos de los Andes Altos (AIGACAA), La Paz. 40 p.
Anderson E., J. Marengo, R. Villalba, S. Halloy, B. Young, D. Cordero, F. Gast, E. Jaimes feD.Ruiz. 2011. Consequences of climate change for ecosystems and ecosystem services in the tropical Andes. Pp: 1-18. En: Herzog, S., R. Martínez, P. Jørgensen & H. Tiessen (eds.) Climate Change and Biodiversity in the Tropical Andes. Inter-American Institute for Global Change Research (IAI) and Scientific Committee on Problems of the Environment (SCOPE), Saint Louis.
Anthelme, F., D. Jacobsen, P. Macek, R. I. Meneses, P. Moret, S. Beck & O. Dangles. 2014. Biodiversity patterns and continental insularity in the high tropical Andes. Arctic, Antarctic and Alpine Research 46: 811-830.
Baraer, M., B. Mark, J. Mckenzie, T. Condom, T. Bury, J. Huh, K. Portocarrero, C. Gómez & S. Rathay. 2012. Glacier recession and water resources in Perú, Cordillera Blanca. Journal of Glaciology 58:134-150.
Benavides, J. C, D. H. Vitt & R. K. Wieder. 2013. The influence of climate chiange on recent peat accumulation patterns of Distichia muscoides cushion bogs in the high-elevation tropical Andes of Colombia. Journal of Geophysical Research: Biogeosciences 118(4): 1627-1635.
Bradley, R., M. Vuille, H. Diaz & W. Vergara. 2006. Threats to water supplies in the tropical Andes. Science 312:1755-1756.
Buytaert, W., F. Cuesta-Camacho & C. Tobón. 2011. Potential impacts of climate change on the environmental services of humid tropical alpine regions. Global Ecology and Biogeography 20:19-33.
Callaway, R. M. 1995. Positive interactions among plants. The Botanical Review 61(4): 306-349.
Cavieres, L. A., R.W. Brooker, B.J. Butterfield, B.J. Cook, Z. Kikvidze, C. J. Lortie, R. Michalet, F. I. Pugnaire, C. Schöb, S. Xiao, F. Anthelme, R.G. Björk, K. J. M. Didcinson, B. H. Cranston, R. Gavilán, A. Gutierrez-Giron, R. Kanka, J. P. Maalouf, A. F. Mark, J. Noroozi, R. Parajuli, G. K. Phoenix, A. M. Reid, W. M. Ridenour, C. Rixen, S. Wipf, L. Zhao, A. Escudero, B. F. Zaitchik, E. Lingua, E. T. Aschehoug & R. M. Callaway. 2014. Facilitative plant interactions and climate simultaneously drive alpine plant diversity. Ecology Letters 17(2): 193-202.
Chimner, R. A. & J.M. Karberg. 2008. Long-term carbon accumulation in two tropical mountain peatlands, Andes Mountains, Ecuador. Mires and Peat 3(4): 1-10.
Cornelissen, J., S. Lavorel, E. Garnier, S. Diaz, N. Buchmann, D. Gurvich, P. Reich, H. Steege, H. Morgan, M. van der Heijden, J. Pausas & H. Poorter. 2003. A handbook of protocols for standardized and easy measurement of plant fundional traits worldwide. Australian Journal of Botany 51: 335-380.
Coronel, J. S., S. Declerck & L. Brendonck. 2007. High altitude peatlands temporary pools in Bolivia house a high Cladoceran diversity. Wetlands 27 (4): 1166-1174.
Earle L., B. Warner & R. Aravena. 2003. Rapid development of fan unusual peat-accumulating ecosystem in the Chilean Altiplano. Quaternary Research 59:2-11.
Estenssoro, S.1991. Los bofedales de la cuenca alta del valle de La Paz. Pp. 109-121. En: Forno, E. & M. Baudoin (eds.) Historia Natural de un Valle en los Andes: La Paz. Instituto de Ecología, Universidad Mayor de San Andrés, Imprenta Quipus, LaPaz.
Flores, A. 2013. Ecomorfología y ecología alimentaria del género Orestias (Pisces Cyprinodontiformes) en la puna xerofítica de la provincia Sud Lípez, Potosí - Bolivia. Tesis de licenciatura en biología, Universidad Mayor de San Andrés, La Paz. 80p.
Gonzales R. K., E. Quenta, J. Molina-Rodriguez, O. Dangles & D. Jacobsen. 2014. Propuesta metodológica para estimar la heterogeneidad ambiental, diversidad y estructura de comunidades acuáticas de pozas de agua en bofedales altoandinos. Ecología en Bolivia 49(3): 56-72.
Hodgson, J. G., G. Montserrat-Martí, M.Charles, G. Jones, P. Wilson, B. Shipley, M. Sharafi, B. E. L. Cerabolini, J. H. C. Cornelissen, S. R. Band, A. Bogard, P. Castro-Diez, J. Guerrero-Campo, C. Palmer, M. C. Pérez-Ron tomé, G. Cárter, A. Hynd, A. Romo-Diez, L. de Torres Espuny & F. Royo-Pla. 2011. Is leaf dry matter content a better predictor of soil fertility than specific leaf area?. Annals of Botany 108(7):1337-1345.
IPCC (Intergovernmental Panel on Climate Change). 2013. Summary for policymakers. pp. 1-29. En: Climate Change 2013: The Physical Science Basis. Contribution of working group I to the 5th assessment report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge y Nueva York. [ Links ]
Kikvidze, Z., R. Brooker, B. Butterfield, R. Callaway, L. Cavieres, B. Cook, C. Lortie, R. Michalet, F. Pugnaire, S. Xiao, F. Anthelme, R. Björk, B. Cranston, R. Gavilán, R. Kanka, E. Lingua, J. Maalouf, J. Noroozi, R. Parajuli, G. Phoenix, A. Reid, W. Ridenour, C. Rixen & C. Schöb. En prensa. The effects of foundation species on community assembly: a global study on alpine cushion plant communities. Journal of Ecological Society of America.
Leps, J. & P. Smilaeur (eds). 2003. Multivariate analysis of ecological data using CANOCO. Cambridge University Press, Cambridge. 267 p.
MacArthur, R.& O.Wilson. 1963. An equilibrium theory of insular zoogeography. Evolution 17: 373-387.
Maldonado, F. 2014. An introduction to the bofedales of the Peruvian high Andes. Mires and Peat 15(5): 1-13.
Meneses, R. I. 1997. Estudio de la vegetación en la zona minera de Milluni (Provincia Murillo, Departamento de La Paz). Tesis de licenciatura en biología, Universidad Mayor de San Andrés, La Paz. 107 p.
Meneses, R. I. 2012. Estado de arte de los bofedales en la Cordillera Real en el área que abarca la cuenca de Khara Khota hasta la cuenca de Choqueyapu. Documento técnico no publicado. Banco Interamericano de Desarrollo (BID), La Paz. 74 p.
Meneses, R. I., S. Loza Herrera, A. Lliully, A. Palabral & F. Anthelme. 2014. Métodos para cuantificar diversidad y productividad vegetal de los bofedales frente al cambio climático. Ecología en Bolivia 49(3): 42-55.
Michalet, F. Pugnaire, S. Xiao, F. Anthelme, R. Björk, B. Cranston, R. Gavilán, R. Kanka, E. Lingua, J. Maalouf, J. Noroozi, R. Parajuli, G. Phoenix, A. Reid, W. Ridenour, C. Rixen & C. Schöb. En prensa. The effects of foundation species on community assembly: a global study on alpine cushion plant communities. Journal of Ecological Society of America.
Milner, A. M., L. E. Brown & D. H. Hannah. 2009. Hydroecological response of river systems to shrinking glaciers. Hydrological Processes 23: 62-77.
MMAyA (Ministerio de Medio Ambiente y Agua). 2012. Libro rojo de la flora amenazada de Bolivia. Vol. I. Zona Andina. La Paz. 583 p. [ Links ]
Ostria, C. 1987. Phytoécologie et paleoécologie de la vallée altoandine de Hichu Kkota (Cordillére oriéntale, Bolivie). Tesis de doctorado, Universidad de París, París. 180 p.
Pacheco, M. 1998. Aprovechamiento de áreas hidromórficas en el altiplano peruano-boliviano. Manejo de bofedales. Cría de alpacas. Cooperación Andina de Fomento y Autoridad Binacional del Lago Titicaca, La Paz, 113 p.
Palabral, A. 2013 Relación de la composición florística y su biomasa subterránea con las variables hidrológicas en los bofedales de Sajama. Documento técnico no publicado. Agua Sustentable, La Paz. 30 p.
Palmer, M. 2007. Species-area curves and the geometry of nature. Pp: 15-31. En: Storch, D., P. Marquet & J. Brown (eds.) Scaling biodiversity, Published by Cambridge University Press, Cambridge.
Preston, F. 1962.The canonical distribution of commonness and rarity: Part I. Ecology 43:185-215.
Quenta, E. 2013. Estructuración espacial de metacomunidades de cladócera en los humedales altoandinos de la Cordillera Real. Tesis de maestría en ecología y conservación, Universidad Mayor de San Andrés, La Paz. 42 p.
Quinn, G. P. & M. J. Keough. 2002. Experimental design and data analysis for biologists. 1ra. Edic. Cambridge University Press, Cambridge. 537 p.
Rabatel, A., B. Francou, A. Soruco, J. Gómez, B. Cáceres, J. L. Ceballos, R. Basantes, M. Vuille, J.-E. Sicart, C. Huggel, M. Scheel, Y. Lejeune, Y. Arnaud, M. Collet, T. Condom, G. Consoli, V. Favier, V. Jomelli, R. Galarraga, P. Ginot, L. Maisincho, J. Mendoza, M. Ménégoz, E. Ramirez, P. Ribstein, W. Suarez, M. Villads & P. Wagnon. 2013. Current state of glaciers in the tropical Andes: a multi-century per spective on glacier evolution and climate change. Cryosphere 7(1): 81-102.
Ramírez, E. 2008. Impactos del cambio climático y gestión del agua sobre la disponibilidad de recursos hídricos para las ciudades de La Paz y El Alto. Revista Virtual REDESMA 2(3): 49-61.
Ruthsatz, B. 2012. Vegetación y ecología de los bofedales altoandinos de Bolivia. Phytoecologia 42:133-179.
Segnini, A., A. Posadas, R. Quiroz, D. M. B. P Milori, S. C. Saab, L. M. Neto & C. M. P. Vaz. 2010. Spectroscopic assessment of soil organic matter in wetlands from the high Andes. Soil Science Sodety of American Journal 74(6): 2246-2253.
Sklenář, P. & P. M. Jørgensen. 1999. Distribution patterns of páramo plants in Ecuador. Journal of Biogeography 26: 681-691.
Squeo, F., B. Warner, R. Aravena &D. Espinoza. 2006. Bofedales: high altitude peatlands of the central Andes. Revista Chilena de Historia Natural 79: 245-255.
Soruco A., T. Berger, J. Mendoza, A. Rabatell, B. Francou, A. Machaca, F. Rojas, T. Alexandrovna, P. Wagnon, J. Philippe, R. Fuertes, L. Noriega, E. Ramirez & H. Valdivieso. 2005. Mediciones gladológicas, hidrológicas y meteorológicas en los glaciares Zongo - Chacaltaya - Charquini sur. Año hidrológico 2003-2004. Institut de Recherche pour le Développement (IRD) & Instituto de Hidráulica e Hidrología (IHH), La Paz. 147 p.
Tellería, J. L., J. L. Venero & T. Santos. 2006. Conserving birdlife of Peruvian highland bogs: effects of patch-size and habitat quality on species richness and bird numbers. Ardeola 53: 271-283.
Veech, J. A., K. S. Summerville, T. O. Crist & J. C. Gering. 2002. The additive partitioning of spedes diversity: recent revival of an old idea. Oikos 99(1): 3-9.
Verzijl, A. & S. G. Quispe. 2013. The system nobody sees: irrigated wetland management and alpaca herding in the Peruvian Andes. Mountain Research and Development 33(3): 280-293.
Villarroel, E. K., P. L. Pacheco Mollinedo, A. I. Domic, J. M. Capriles & C. Espinoza. 2014. Local management of Andean wetlands in Sajama National Parle, Bolivia: persistence of the collective system in increasingly family-oriented arrangements. Mountain Research and Development 34 (4): 356-368.
Zeballos, G. 2013. Cuantificación de la variabilidaddelaextensióndehumedales, lagunas, y nevados, de la Cordillera Real (15º45' - 16º45' sur, 67º40'- 68º40' oeste), entre 1984 y 2009, empleando imágenes LANDSAT TM. Tesis de grado de licenciatura en geografía, Escuela Militar de Ingeniería Mcal. Antonio José de Sucre, La Paz. 154 p.
Artículo recibido en: 06 de Febrero de 2015.
Manejado por: Blanca León.
Aceptado en: 27 de Abril de 2015.