









 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO  uBio
uBio 
 Permalink
PermalinkEcología en Bolivia
versão impressa ISSN 1605-2528versão On-line ISSN 2075-5023
Ecología en Bolivia vol.50 no.1 La Paz abr. 2015
Artículo
Efecto de la altitud y el micrositio sobre plántulas reforestadas de Polylepis lanata (Rosaceae) en el noroeste de Cochabamba, Bolivia: implicaciones para su restauración ecológica
Effect of altitude and microsite on reforested seedlings of Polylepis lanata (Rosaceae) in Northwestern Cochabamba, Bolivia: implications for its ecological restoration
Ariel Isaías Ayma-Romay1*, Gladys Soto-Rojas1,2 & Pedro Lovera3
1Fundación para la Autogestión del Medio Ambiente (FUPAGEMA) - Proyecto de Manejo y Restauración de Bosques en Independencia de la Foundation Rufford Small Grants. Jacinto Anaya # 1769, Cochabamba, Bolivia, * Autor para correspondencia: ariel.isaias.aymar@gmail.com
2Escuela de Ciencias Forestales, Universidad Mayor de San Simón, Casilla 447, Cochabamba, Bolivia.
3Gobierno Autónomo Municipal de Independencia, Av. Independencia c. 6 de agosto. Cochabamba, Bolivia.
Resumen
La reforestación de Polylepis lanata (Rosaceae) está limitada por el pobre conocimiento de las condiciones ambientales que necesitan sus plántulas para sobrevivir y crecer. El objetivo de la investigación fue determinar cómo el efecto de la altitud y las condiciones de micrositio influyen en la sobrevivencia y crecimiento de las plántulas reforestadas. Se implementó un experimento de reforestación de plántulas extraídas del bosque, en 15 parcelas distribuidas en cinco pisos altitudinales en un rango de 2.690-3.900 m (tres parcelas por piso altitudinal). El lugar de estudio fue en la comunidad de Pajchanti, al noroeste de Cochabamba, Bolivia. Se evaluaron la sobrevivencia y crecimiento de las plántulas en dos periodos, a los 14 y 51 meses después de la reforestación, respecto a la altitud y micrositio (cobertura de arbustos, piedras, hierbas, musgos, pastos, profundidad de humus y pendiente). Los datos se analizaron con el ajuste de modelos lineares generalizados y la selección de modelo e inferencia multi-modelo basado en el criterio de información de Akaike. Los modelos seleccionados sugieren que el incremento de altitud y la cobertura de arbustos fueron las variables más importantes para mejorar la sobrevivencia. Por otro lado, la profundidad de humus, cobertura de piedras y arbustos fueron los más importantes para mejorar el crecimiento. Las reforestaciones de la especie pueden realizarse en altitudes intermedias (3.000-3.600 m), pero asociados a micrositios seguros que protejan a las plántulas de las condiciones ambientales que posiblemente sean estresantes afuera del bosque, debido a la perturbación humana del suelo y la vegetación.
Palabras clave: Facilitación, Límites de bosque, Nodriza, Reforestación, Restauración.
Abstract
The reforestation of Polylepis lanata (Rosaceae) seedlings is limited by the poor knowledge about the environmental conditions that needs to survive and grow. The objective of the study was to determine how altitude and microsite affect seedlings survival and growth. An experiment was implemented with seedlings gathered from the forest and planted on 15 plots distributed in five altitudes (2,690-3,900 m; three replicates per altitude). In each plot, seedlings were planted in heterogeneous microsites. The study was carried out in the community Pajchanti in Northwestern Cochabamba, Bolivia. Survival and growth were assessed 14 and 51 months after planting. We measured altitude of plots and microsites around of each plant (cover shrubs, stones, moss, herbs, grassland, depth of humus and slope). The data analysis was conducted through the fit of generalized linear models, model selection and inference multi-model based in Akaike Information Criteria. The altitude and shrub cover had a positive effect on seedlings increasing their probability of survival; both were the most important variables. The humus, cover of stones and shrubs around the seedlings also improved the growth rates. The reforestations should be carried out in middle altitudes, 3,000 to 3,600 m, but in safe microsites which protect to the seedlings from a possible environmental stress outside of the forest, due to human disturbance of soil and vegetation.
Keywords: Facilitation, Forestation, Nurse, Restoration, Tree line.
Introducción
Los árboles de queñua - Polylepis lanata (Kuntze) M. Kessler & Schmidt-Leb. son endémicos de Bolivia (Navarro 1997). Estos son considerados en peligro de extinción, igual que otras especies del género debido a la tala y la quema (Meneses & Beck 2005, Ayma-Romay et al. 2009). La reforestación de esta especie es importante debido a los múltiples bienes y servicios ecosistémicos que brindan a las poblaciones locales (Fjeldså & Kessler 2004). Sin embargo, aún existen pocas investigaciones o experiencias al respecto.
En las montañas la altitud y las condiciones de micrositio pueden desempeñar un papel muy importante para la distribución y crecimiento de las plántulas arbóreas (Körner & Paulsen 2004). Por un lado, las condiciones ambientales cambian con la altitud, estableciendo condiciones óptimas y marginales para su distribución y crecimiento; dependiendo la especie, altitudes muy bajas o altas pueden generar condiciones ambientales estresantes, marcando límites para su establecimiento (Körner & Paulsen 2004, Smith et al. 2009, Anthelme & Dangles 2012). Por otro lado, los componentes bióticos y abióticos del micrositio alrededor de las plántulas arbóreas pueden aminorar o incrementar el estrés ambiental del entorno, influyendo de manera importante en la sobrevivencia o crecimiento de las plántulas. Por ejemplo, un micrositio seguro puede conducir a las especies vegetales hacia lugares más estresantes ambientalmente, porque pueden reducir el estrés ambiental o suministrar más recursos (Fowler 1988, Körner & Paulsen 2004).
Aun no se comprende cómo puede influir la altitud y micrositio en las plántulas de P. lanata para establecerse. Por un lado, estudios realizados con otras especies de Polylepis dan cuenta que estas especies pueden sobrevivir y crecer con amplia distribución si se evitan las perturbaciones de origen humano (fuego, pisoteo y ramoneo de ganado) (Kessler et al. 2007, Renison et al. 2013). Por ejemplo, se han observado actuales manchas de bosques maduros de P. lanata entre 2.800-4.000 m de altitud, sugiriendo amplia distribución altitudinal y la capacidad de establecerse en diferentes condiciones ambientales (Fjeldså & Kessler 2004). También existe evidencia de que plántulas de regeneración de P. incana, P. pauta de Ecuador (Cierjacks et al. 2007) y plántulas de reforestadas de P. australis del centro de Argentina (Marcora et al. 2013) pueden expandirse más arriba o abajo de la distribución actual de los bosques.
Por otro lado, otros estudios dan cuenta que especies de Polylepis requieren sitios microclimáticamente especiales para protegerse del estrés ambiental de la montaña (Gosling et al. 2009, Williams et al. 2011), condición que les dificultaría expandirse en amplios rangos altitudinales o ambientales. Por ejemplo, lugares de topografía accidentada con buen drenaje, mayor humedad y temperaturas más cálidas fueron muy importantes para el establecimiento de varias especies del género (Simpson 1986). Lugares con piedras también protegieron a las plantas de estas especies brindando calor en las noches frías y mayor humedad en el suelo en días calientes (Körner & Paulsen 2004). Además, la vegetación de pastizales y otros árboles alrededor de las plántulas facilitaron el establecimiento y crecimiento de P. australis, reduciendo condiciones estresantes, como la alta radiación, temperaturas extremas y mejorando la infiltración de la lluvia (Enrico et al. 2004, Renison et al. 2005, Landi & Renison 2010). El objetivo de esta investigación fue determinar el efecto de la altitud y las condiciones de micrositio en la sobrevivencia y crecimiento de las plántulas reforestadas de P. lanata en la localidad de Pajchanti (Cochabamba, Bolivia), para así recomendar algunas técnicas de reforestación de la especie con criterios ecológicos. La pregunta de investigación fue: ¿Cómo varía la sobrevivencia y crecimiento de las plántulas de esta especie en diferentes altitudes de la montaña y en distintos micrositios fuera del bosque?
Metodología
Área de estudio
La comunidad de Pajchanti se encuentra en el municipio de Independencia, provincia Ayopaya al noroeste de Cochabamba, Bolivia (17º 5' Sur y 66º 49' Oeste). El área muestra diferentes condiciones climáticas y de vegetación a lo largo del gradiente altitudinal de la montaña de 2.500-4.200 m (Fig. 1), propias de los Yungas bolivianos (distrito Yungas del Cotacajes) con transición o contacto con la Puna Peruana (distrito del Tunari). El bioclima es pluviestacional y cambia de altitudes más bajas hacia más altas, de mesotropical a orotropical, con temperaturas y precipitaciones promedio anual, respectivamente, de 14ºC y 894 mm (2.500 m) hacia 7.4ºC y 560 mm (4.200 m). Las heladas son poco frecuentes. La vegetación nativa a lo largo del gradiente altitudinal ha sido modificada al menos 50% para habilitar áreas agrícolas y de pastoreo (Ayma-Romay et al. 2009). La vegetación del piso montano de 2.500-2.900 m está dominada por arbustos de los géneros Baccharis y Berberis. El piso altimontano (2.900-3.500) presenta manchas de bosque alto de Podocarpus glomeratus y Weinmannia microphylla, arbustos de Baccharis, Berberís y Barnadesia y pastizales (Festuca dolichophylla) con arbustos dispersos. En la ceja de monte, de 3.500-3.800 m, predominan pastizales, manchas de bosque bajo y árboles dispersos de P. lanata. En el piso alto andino de 3.800-4.200 m predominan pastizales, áreas rocosas y solamente algunos árboles dispersos de P. lanata y otros arbustos (Navarro 2005, Ayma-Romay et al. 2007).

El uso de suelo no es muy antiguo, existen evidencias de que el bosque de 2.500-3.800 m no fue deforestado hasta 1960. Sin embargo, es posible que en ceja de Yungas y piso altoandino, la deforestación fuera más antigua para el pastoreo de ovejas y llamas (Ayma-Romay et al. 2009). Actualmente, las manchas de bosque están siendo protegidas por la comunidad, raramente existen incendios y tala de P. lanata desde hace una década debido a mejoras de reglas de uso; además no es una especie preferida para uso maderable (Ayma-Romay 2012). Las evidencias de erosión del suelo son escasas. La ganadería es extensiva, rotativa y de moderada intensidad (obs. pers.).
La especie del estudio pertenece a la familia Rosaceae. Es endémica de Bolivia y fue elevada recientemente a rango de especie Polylepis lanata (Kuntze) M. Kessler & Schmidt-Leb. por Kessler et al. (2005). En el país es considerada en peligro de extinción, se conoce muy poco sobre su distribución y estado de conservación (Meneses & Beck 2005). Es un árbol de ocho metros de altura, de tronco retorcido, ramificado, corteza laminada, hojas compuestas imparipinadas de siete foliolos y con pelos largos y lanosos al envés. Las flores y semillas son pequeñas y se polinizan por el viento (Fjeldså & Kessler 2004). Esta especie ha sido observada en el Parque Nacional Tunari (Kessler et al. 2005), en la provincia Ayopaya (norte del municipio de Independencia) del departamento de Cochabamba y en el sur del municipio de Inquisivi del departamento de La Paz (Navarro 2005). En Independencia, estos árboles forman pequeñas manchas de bosque de dosel abierto o árboles dispersos de 3.300-3.600 m alrededor de pastizales, en contacto o asociados con especies de ceja de Yungas, como Vallea stipularis, Alnus acuminata, P. glomeratus, W. microphylla y otras (Fjeldså & Kessler 2004, Ayma-Romay et al. 2009). Ayudan en la captación agua de la neblina, protegen suelos frágiles de los deslizamientos, forman hábitats para otras especies endémicas, proveen leña, tintes para tejidos, medicinas y madera, entre otros (Bach et al. 1999, Ayma-Romay et al. 2009).
Métodos
Se plantaron a raíz desnuda 150 plántulas de P. lanata de 3.75 cm de altura±DE 1.77. Estas fueron extraídas de diferentes partes del bosque de Pajchanti a una altitud de 3.400 m y se plantaron en 15 parcelas de 17 x 17 m afuera del bosque (con pastizales y arbustos dispersos) repartidas en cinco pisos altitudinales (Altitud A = 2.747 m, B = 3.005 m, C = 3.300 m, D = 3.590 m y E = 3.850 m) y tres repeticiones por piso. En cada parcela se plantaron diez plántulas distanciadas aproximadamente tres metros, en hoyos de 20 x 20 cm, cada una en condiciones de micrositio particulares. Las plántulas no recibieron riego ni otros tratamientos.
La plantación se realizó en verano (temporada de lluvia). Los dos primeros años las parcelas estuvieron clausuradas con alambre de púas para evitar el ingreso del ganado, a partir del segundo fueron abiertas a solicitud de la comunidad, lo que no tuvo muchas repercusiones porque todas tuvieron el mismo tratamiento y las perturbaciones fueron muy bajas (hubo muy bajas señales de ganadería).
Evaluación de campo
Se realizaron evaluaciones a los catorce y 51 meses después de la reforestación de plántulas (ambas después de la temporada alta de lluvias). Estos periodos de evaluación estuvieron sujetos a nuestras posibilidades operativas y financieras de realizar el trabajo de campo. El primer periodo de evaluación fue desde la plantación (febrero de 2009) hasta los 14 meses (abril de 2010) y el segundo periodo de 14-51 meses (abril de 2013). Se evaluaron la sobrevivencia, crecimiento y micrositio de cada plántula en cada periodo. Para la sobrevivencia se registró cada planta si estaba viva o muerta, obteniendo una variable de respuesta binomial (planta viva =1; planta muerta = 0) y para el crecimiento se obtuvo la diferencia de altura de cada planta entre periodos, la altura se midió desde el cuello del tallo hasta el ápice de la rama más larga y dominante (esta variable fue continua).
El micrositio de cada planta se evaluó como el ambiente más cercano a un metro de distancia de cada plántula formando un círculo de 3.14 m2. En esta área se estimaron el porcentaje de cobertura ocupado por musgos y hojarasca, pastizal, hierbas, arbustos, piedras y suelo descubierto, sumando 100%. También a 20 cm de distancia de cada plántula, con orientación al azar, se midieron la profundidad de humus y hojarasca en centímetros y la pendiente en porcentaje.
El micrositio de las plántulas en el primer periodo de evaluación fue dominado por la cobertura de pastos F. dolichophylla (46%), musgos y hierbas (30%), escasa cobertura de arbustos (10%), piedras (8%) y suelo desnudo (6%). Para el segundo periodo no cambio mucho, siguieron predominando los pastos (58%), musgos y hierbas (30%) con escasa cantidad de arbustos (7%), piedras (7%) pero sin suelo desnudo ya que fue cubierto por las hierbas, musgos y pastos. La pendiente fue en promedio 5% y la profundidad de humus y hojarasca muy escasa o a veces ausente.
Análisis de datos
Se realizó un análisis exploratorio con el ajuste de modelos lineales generalizados (MLG) (Dobson 1990), la selección de modelos basados en el AIC (Criterio de Información Akaike) (Akaike 1973) modificado para pequeñas muestras AICc (Hurvich & Tsai 1989) y un proceso de inferencia multi-modelo a partir del mejor conjunto de modelos seleccionados a través de índices derivados del AICc (Johnson & Omland. 2004, Symonds & Moussalli 2011, Burnham et al. 2011).
Para el ajuste de MLG se partió de modelos saturados que incluyeron la sobrevivencia (planta viva; planta muerta) o crecimiento como variables de respuesta, en función de factores de bloque (3 niveles), altitud (cinco niveles) y las variables continuas de micrositio que no mostraron alta colinealidad (como covariables). Los modelos que correspondieron a sobrevivencia se ajustaron con la distribución de errores de la familia binomial y vínculo de enlace logístico, y los de crecimiento con la familia gaussiana y vínculo de enlace identidad.
Como fueron dos periodos de evaluación para cada variable de respuesta (S= sobrevivencia y C = crecimiento) el análisis partió del ajuste de los siguientes cuatro modelos saturados, por separado:
S (0-14 meses)= Bloque + Altitud + Arbustos + Musgos + Pendiente + Piedras + Humus
S (14-51 meses) = Bloque + Altitud + Arbustos +Pendiente + Piedras
C(0-14 meses)= Bloque + Altitud + Musgos + Arbustos + Pendiente + Piedras + Humus
C (14-51 meses) = Bloque + Altitud + Arbustos + Pendiente + Piedras + Humus
Las variables con alta colinealidad fueron removidas de los modelos. Entre estos: la cobertura de pastizales, por su alta correlación negativa con arbustos, hierbas y musgos, los musgos o hierbas por su correlación positiva con arbustos y a veces con piedras y humus por su correlación con arbustos. Estas correlaciones fueron inspeccionadas con coeficientes de correlación de Pearson y el factor de inflación de varianza (Fox & Monette 1992, Zuur et al. 2010). La variable de suelo descubierto fue removida porque no registro datos para el segundo periodo.
Los datos de las variables continuas, crecimiento, piedras, arbustos, pendiente y humus no tuvieron una distribución normal y mostraron sobre-dispersión, pero decidimos no realizar transformaciones de los mismos, para no complicar la interpretación de los parámetros de los modelos (Zuur et al. 2010). Además, se sostiene que la estadística paramétrica es muy robusta, incluso violando el supuesto de normalidad (Understood 1997).
Bajo la perspectiva de selección de modelos basado en AIC, a partir de cada modelo saturado según correspondía, se generaron 2n modelos alternativos con todas las combinaciones posibles de las n variables regresoras más uno del intercepto (por ejemplo, del modelo saturado de sobrevivencia del primer periodo, se generaron 28 = 256 modelos alternativos). De acuerdo con sus valores AICc los modelos fueron posicionados con un "ranking", así modelos alternativos que tuvieron un valor de AICc más pequeño fueron considerados mejores modelos (ya que proporcionaron más evidencia para explicar la variación de la variable de respuesta dado los datos y tuvieron menor número de variables, principio de parsimonia). Debido a que la selección de un solo mejor modelo ocultaba a otros posibles modelos igual o cercanamente buenos, se realizó un procedimiento de inferencia multi-modelo (Johnson & Omland 2004), que consistió realizar inferencias del mejor conjunto de modelos. El mejor conjunto de modelos tuvieron una diferencia (Δ AICc) menor a dos respecto al modelo con menor AICc (Burnham et al. 2011). La inferencia comenzó comparando entre modelos el peso Akaike (wi, que mide la probabilidad, en escala 0 -1, de que un Xi modelo sea el más próximo y con más evidencia para explicar mejor la variable de respuesta dado los datos y el conjunto de modelos seleccionados) y la Proporción de Evidencia (ER = que mide para el mejor modelo, cuantas veces es más probable que sea el mejor respecto a otros del conjunto de modelos). Asimismo, continuó con la comparación de la Importancia Relativa de cada variable en el mejor conjunto de modelos (I, que se entiende como la probabilidad, en escala 0-1, que una variable sea parte del mejor modelo) (Symonds & Moussalli 2011). La exploración de los datos y el análisis estadístico se realizó en el ambiente R - Project versión 3.1.1 desarrollado por la R Foundation for Statistical Computing, apoyados en paquetes instalados por defecto en el programa "car, MASS" (R Core Team 2014) y el paquete "glmulti" para realizar la programación de selección de modelo y cálculos para la inferencia multi-modelo (Calcagno & Mazancourt 2010).
Para comprender mejor como varió la probabilidad de sobrevivencia (modelo logístico) o crecimiento de las plántulas (modelo lineal) en función a las variables predictoras continuas o categóricas más importantes en el mejor conjunto de modelos, se hicieron modelaciones de cada una en función a la de respuesta, por separado, utilizando los parámetros o coeficientes de los mejores modelos donde éstas estuvieron presentes (Underwood 1997). Si la variable para modelar fue continua, para ésta se creó una serie de valores, en el rango de sus datos observados, si fue factor, cada nivel fue asignado con el valor de su coeficiente. Cuando hubo más de una variable en el modelo, estas fueron asignadas con un valor fijo, por ejemplo, promedio de sus valores observados para variables continuas, y cero para niveles de factores (Grueber et al. 2011). Este procedimiento se realizó de manera manual en MS- Excel 2012.
Resultados
Sobrevivencia
De 150 plántulas reforestadas, a los catorce meses sobrevivieron 80 individuos (53% del total) y a los 51 meses, fueron 51 (34% del total). La sobrevivencia fue mayor en pisos altitudinales intermedios en ambos periodos de evaluación; el promedio de sobrevivencia fue de 40, 60, 70, 63.3 y 33% (primer periodo) y de 25, 66, 71, 84 y 50% (segundo periodo), según los diferentes niveles de altitud (de abajo hacia arriba) (Fig. 2).
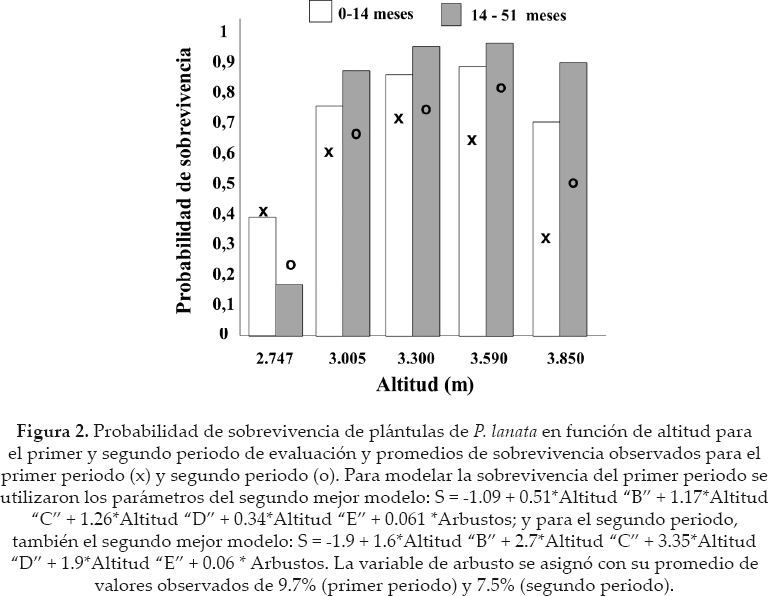
El mejor conjunto de modelos para explicar la sobrevivencia del primer periodo de evaluación estuvo formado por seis modelos y para el segundo periodo por cuatro. Aunque todos estos modelos fueron los más parsimoniosos y tuvieron más evidencia para explicar los datos, existieron algunas diferencias entre sus pesos Akaike y ER. Para ambos periodos de evaluación de sobrevivencia, los modelos con un ER igual o menor 1.4 respecto al mejor modelo, fueron los más simples, parsimoniosos y los que incluyeron a la altitud y cobertura de arbustos como variables con un efecto positivo en la sobrevivencia (ver signos de los niveles de altitud y arbustos en los modelos) (Tabla 1) y las más importantes (Tabla 2). Además para el primer periodo, un modelo incluyó a la pendiente, también como una variable que influyó positivamente en la sobrevivencia. Los modelos con un ER igual o mayores a 1.8, fueron versiones más complejas, en las cuales, las variables de arbustos o la altitud fueron complementadas por variables de menor importancia (Tablas 1 y 2), como el efecto positivo del humus, el efecto positivo de piedras, a veces por un efecto positivo de la pendiente (primer periodo) o negativo de la misma para el segundo periodo (Tabla 1). Estos últimos modelos tuvieron menor probabilidad para ser los mejores modelos (comparar pesos Akaike y ER entre modelos (Tabla 1).
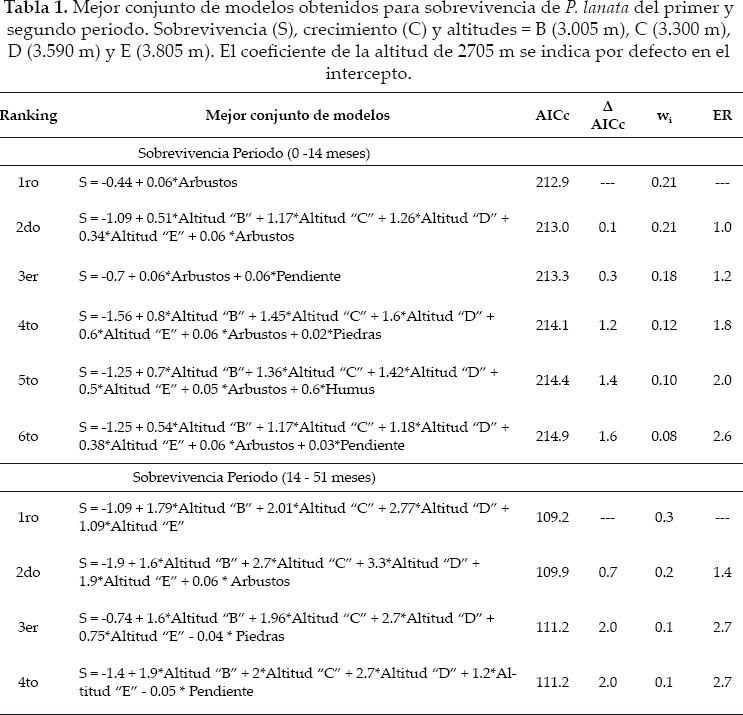
Si bien la altitud y los arbustos fueron las variables más importantes, se debe notar que los arbustos fueron más importantes en la primera fase de establecimiento de las plántulas, y la altitud la más importante en el segundo periodo (Tabla 2). Variables como el bloque, musgos y hierbas fueron removidas del mejor conjunto de modelos de sobrevivencia. Pero como los musgos y a veces las hierbas tuvieron correlación positiva con los arbustos, no se debería descartar su posible función junto con los arbustos.
La modelación de sobrevivencia en función de la altitud fue unimodal en ambos periodos de evaluación, mayor probabilidad de sobrevivencia en altitudes intermedias y menor probabilidad en los extremos. Sin embargo, la probabilidad de sobrevivencia en la altitud más alta es mayor que en la inferior (Fig. 2). El signo negativo del coeficiente del primer término de los modelos donde se incluye altitud, sugiere que la altitud inferior disminuyó la sobrevivencia, respecto a las altitudes superiores que la aumentan (Tabla 1).
La modelación también sugiere que la cobertura de arbustos mejoró la sobrevivencia de manera logística. Coberturas leves e intermedias < 35 % llegaron a mejorar la probabilidad de sobrevivencia hasta 90%. Plántulas sin cobertura de arbustos tuvieron una probabilidad de sobrevivencia menor, de 40 o 50%, para el primer y segundo periodo de evaluación, respectivamente (Fig. 3).

Crecimiento
Las plántulas a 51 meses del experimento alcanzaron una altura promedio de 27.42 cm ± DE 24.53 (aproximadamente 6.45 cm.año1). Para el primer y segundo periodo, el mejor conjunto de modelos estuvieron formados por tres modelos, respectivamente. El mejor modelo del primer periodo incluyó a la profundidad de humus y a la cobertura de piedras, como variables que influyeron positivamente en el crecimiento de las plántulas (ver signo del parámetro en los modelos) (Tabla 3, Figs. 4 -5) y además fueron los más importantes en el conjunto de modelos seleccionados (Tabla 2).
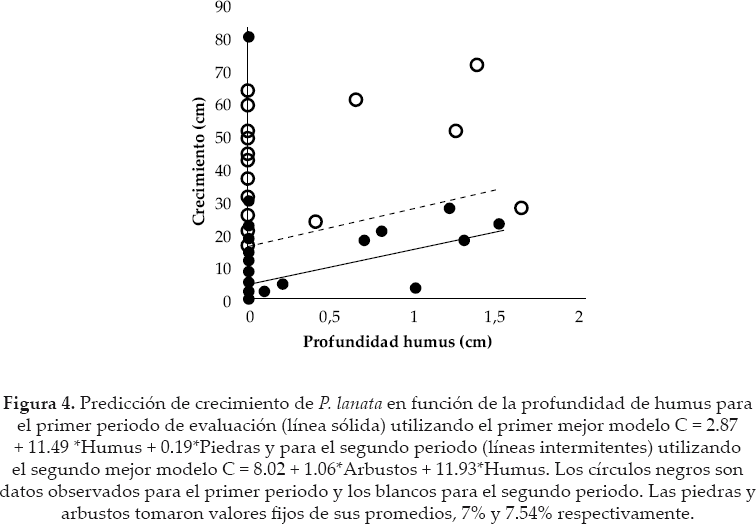
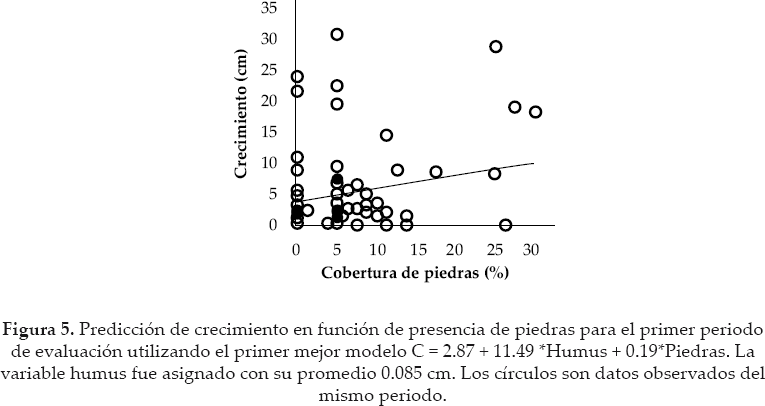
Los otros modelos fueron más complejos que adicionaron a la pendiente o musgos como variables explicativas, pero estos tuvieron menos evidencia que el primero.
En el segundo periodo, el primer y segundo mejor modelo tuvieron prácticamente similar probabilidad para ser el mejor modelo (ver semejanzas de pesos Akaike y ER); ambos sugieren un efecto positivo de arbustos y humus para el crecimiento de las plántulas (ver signos de los parámetros de los modelos) (Tabla 3, Figs. 4 y 6).


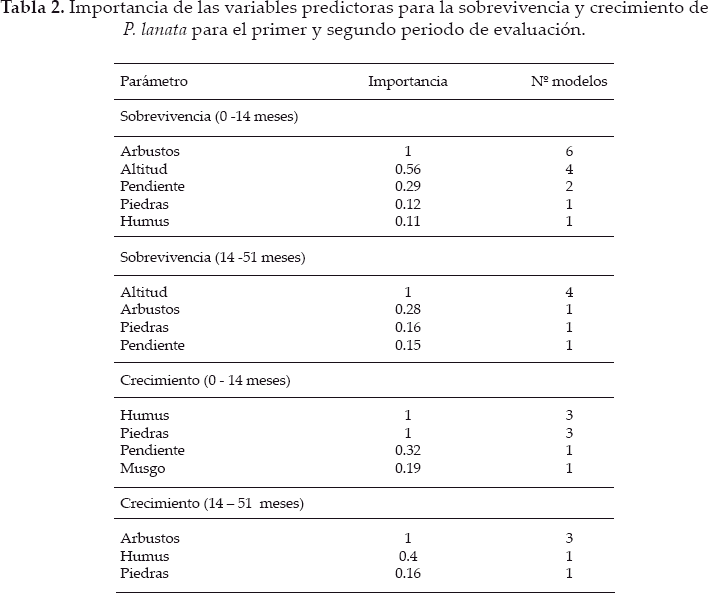
Los factores de altitud, bloque, hierbas fueron totalmente removidos del mejor conjunto de modelos, en ambos periodos. La profundidad de humus y la cobertura de piedras fueron igual de importantes en el primer periodo de evaluación, seguido con menor importancia por la pendiente y los musgos. En el segundo periodo, los más importantes fueron arbustos y humus (Tabla 2).
Discusión
Gradiente altitudinal y micrositio
Las plántulas de P. lanata pueden sobrevivir más arriba o abajo del rango altitudinal de los actuales bosques maduros, acompañado de micrositios seguros que facilitan su sobrevivencia y crecimiento. Existen evidencias para afirmar que altitudes intermedias(3.005-3.590m),seguido de más altas (3.805 m) sean mejores que las altitudes más bajas (2.747 m) del gradiente altitudinal. En otros estudios, las plántulas de otras especies de P. australis de Argentina (Marcora et al. 2013), P. incana y P. pauta de Ecuador (Cierjacks et al. 2007) tuvieron similar potencialidad para expandirse fuera de los límites altitudinales actuales de los bosques maduros, pero igual mostrando una distribución unimodal. Esto podría ser posible, porque las condiciones ambientales estresantes para especies de Polylepis, como temperaturas promedio por debajo de 6.5 ºC, vientos fuertes, alta radiación y heladas frecuentes en las montañas tropicales empezarían arriba de 3.900 o 4.200 m de altitud (Kessler et al. 2007, Anthelme & Dangles 2012). Por otro lado, temperaturas altas, mayores a 15 ºC que provocan estrés hídrico a las plántulas (Marcora et al. 2013) empezarían a partir de altitudes inferiores a 2.700 m (Navarro 2005). Es más probable que estas condiciones ambientales estresantes no estuvieron presentes en los pisos altitudinales intermedios (3.000 a 3.600 m), pero posiblemente tuvieron un efecto en los extremos del gradiente altitudinal estudiado (principalmente el inferior).
Al parecer la potencialidad de las plántulas de P. lanata para expandirse hacia otras altitudes fuera del bosque, bajo las condiciones actuales de suelo y vegetación, también está facilitada por micrositios seguros, formados principalmente por arbustos, piedras y presencia de humus, que incrementan la probabilidad de sobrevivencia, y principalmente sus tasas de crecimiento; esto ya fue observado por otros estudios (Körner & Paulsen 2004, Anthelme & Dangles 2012).
Aunque esto no es compartido por algunos investigadores que suponen que las especies de Polylepis pueden expandirse por un amplio rango altitudinal y en diferentes condiciones micro-climáticas (Fjeldså & Kessler 2004, Kessler et al. 2007, Renison et al. 2013), algunos otros apoyan lo observado, porque atribuyen que las condiciones micro-climáticas especiales son un factor muy importante para el establecimiento de especies de Polylepis, ya que brindan ambientes menos fríos, con menos radiación y más humedad (Simpson 1986, Smith et al. 2009). La necesidad de micrositios por parte de las plántulas de P. lanata, puede ser explicado porque afuera del bosque el estrés ambiental causado por la modificación antrópica del suelo y la vegetación sea importante, además de las condiciones climáticas adversas en los extremos altitudinales (Renison et al. 2006, Anthelme & Dangles 2012). Se conoce ampliamente que los suelos deforestados con historia de uso ganadero y de fuego están sujetos a alta radiación, altas y bajas temperaturas, poca infiltración de agua, alta evapotranspiracióny poca materia orgánica limitando el establecimiento de plántulas. Por ejemplo, la presencia de pastizales (Festuca dolichiphylla) y chuchurrumi (Eryngium rauhianum) presentes en el área de estudio son indicadores de estas condiciones estresantes (Navarro 2005). Aunque el área no es de largo uso, y su grado de degradación no es tan fuerte como en otras áreas de los Andes, al parecer serían suficientes para causar estrés ambiental para las plántulas. Similares condiciones, aunque con signos de degradación más severas del suelo, en el centro de Argentina, también limitaron el establecimiento de plántulas reforestadas de P. australis (Renison et al. 2005, 2006, Torres et al. 2008).
Facilitadores para la sobrevivencia de P. lanata
Arbustos
En muchos estudios ya se han encontrado interacciones positivas de los arbustos para la sobrevivencia de los árboles, porque estos mejoran las condiciones abióticas para las plántulas arbóreas; por ejemplo, reducen las temperaturas extremas (altas y bajas), la alta radiación (dando sombra a las plántulas durante algunas horas en el día), reducen el estrés hídrico y protegen a las plantas de daños físicos provocados por el viento entre otros (Castro et al. 2002, 2004, Gómez-Aparicio et al. 2004). Este es uno de los primeros reportes de facilitación de arbustos en una especie de Polylepis. Por otro lado, en el centro de Argentina se evidenció que las plántulas de P. australis estaban establecidas cerca de árboles adultos de la misma especie, en bosques de dosel ralo, sugiriendo facilitación (Enrico et al. 2004). Asimismo, el mayor crecimiento de P. australis estuvo asociado a mayor cobertura de pastizales en áreas más degradadas (Renison et al. 2005). Lo probable es que grados intermedios o ralos de cobertura de arbustos faciliten la sobrevivencia de las plántulas fuera del bosque; ya que se ha observado que coberturas densas de árboles maduros de Polylepis u otros árboles pueden ejercer competencia por recursos (principalmente luz) reduciendo las densidades y tasas de crecimiento de plántulas (Enrico et al. 2004, Fjeldså & Kessler 2004, Kessler et al. 2007, Cierjacks et al. 2008).
Pendiente y piedras
Aunque la pendiente y las piedras no influyeron de modo tan importante en la sobrevivencia de las plántulas, podría ser necesario hacer notar que estas tuvieron un efecto positivo en el primer periodo de evaluación y generalmente con un leve efecto negativo en el segundo. Por un lado, las pendientes inclinadas son lugares más húmedos, tienen menor exposición solar y las temperaturas bajas son menos probables que en lugares planos (Fjeldså & Kessler 2004, Renison et al. 2006). Las piedras pueden reducir la evaporación del suelo e incrementar la infiltración del agua, reduciendo la escorrentía en lugares inclinados y dando mayor humedad en el suelo para las plantas (Peters et al. 2008). Los mismos autores encontraron que las piedras pueden ofrecer protección física a cactus de ecosistemas áridos reduciendo altas temperaturas del suelo en el día y bajas temperaturas en las noches y prolongando la humedad del suelo después de una lluvia. Renison et al. (2005) también encontró mayor sobrevivencia y crecimiento de plántulas de P. australis en lugares pedregosos y en otro estudio en el norte de Chile se encontró un efecto positivo de rocas grandes con Polylepis tarapacana (Kleier & Lambrinos 2005). Por otro lado, el efecto negativo de estos puede ser explicado porque pendientes más elevadas tienen mayor escorrentía y más estrés hídrico cuando tienen poca vegetación asociada; las piedras también pueden ser indicadores de erosión y suelos pobres en nutrientes, dificultosos para el enraizamiento y almacena calor en días secos y calientes provocando marchitez de las plántulas arbóreas (Pimentel 2007).
Facilitadores para el crecimiento
Humus y hojarasca
A pesar de que el suelo de los micrositios evaluados presentaron poca profundidad de humus (máximo 1.7 cm de profundidad), se evidenciaron beneficios para el crecimiento de las plántulas. Los beneficios del humus y hojarasca para las plantas son muy conocidos y utilizados en la agricultura. Por ejemplo, su color oscuro puede almacenar calor en lugares fríos, reduciendo el daño por heladas en las noches a las plantas, aportar nutrientes micro y macro nutrientes al suelo (Stevenson 1994); así como el ácido húmico, que mejora el crecimiento de las raíces y parte foliar de las plantas, facilitando la absorción de nutrientes, el intercambio de micro y macro nutrientes, los procesos de fotosíntesis, vigor de la planta, la captación y almacenamiento del agua entre otros (Chen & Aviad 1990, Stevenson 1994).
En Argentina, plántulas reforestadas de P. australis también tuvieron mejor crecimiento en suelos mejor preservados oscuros, con mayor cantidad de materia orgánica que en suelos degradados (Renison et al. 2004, Torres et al. 2008). Enrico et al. (2004) encontró árboles de P. australis más altos y menos arbustizados en suelos mejor preservados con materia orgánica que en degradados. Incluso los árboles de la misma especie en suelos mejor preservados produjeron semillas con más capacidad germinativa y sus plantas provenientes tuvieron mejor sobrevivencia y crecimiento (Renison et al. 2004). Algo muy importante si se desea que las plántulas reforestadas se reproduzcan, cumpliendo esta vital función para la especie.
Arbustos y piedras
También se encontró que los arbustos pudieron mejorar el crecimiento de las plántulas, ya que al parecer estos pueden proteger físicamente a la elongación de nuevas partes vegetativas de las plántulas de la radiación; además porque pueden aportar materia orgánica al suelo (Castro et al. 2002). Hay que hacer constar que los arbustos y humus tuvieron una moderada correlación (datos no mostrados en este estudio), lo que sugiere que ambos pueden ser muy importantes. Las piedras pudieron tener la misma función que los arbustos; pero además como se relacionan con mayor infiltración y humedad, pudieron dar mayor disponibilidad de agua facilitando el crecimiento de las plántulas.
Recomendaciones para reforestar P. lanata
En Bolivia, la reforestación de P lanata no es considerada atractiva debido a su baja sobrevivencia y crecimiento en comparación con especies exóticas del género Eucalyptus y Pínus (Fjeldså & Kessler 2004, Ayma et al. 2009). Sin embargo, es importante conocer las condiciones ambientales (altitudes y micrositios) para la especie y adaptar las técnicas de reforestación a estas. Contrariamente, en esquemas convencionales de reforestación diseñados para monocultivos, los arbustos son vistos como competidores y son eliminados, lugares con piedras y de topografía accidentada son poco preferidos porque hacen difícil las operaciones de plantación (Pimentel 2007). Asimismo, las plántulas en el área de reforestación mantienen distancias parecidas unas con otras, y así no se considera la heterogeneidad ambiental del área.
Plantar plántulas de la especie entre 3.000 y 3.600 m de altitud puede ser recomendado como óptimo; sin embargo, buscar micrositios seguros durante la plantación puede ser muy relevante. Esto puede obtenerse dirigiendo las plantaciones cerca de arbustos que ocupan ralamente el área de plantación, como Berberís, Brachyotum, Baecharis, Miconia entre otros (Navarro 2005) o plantando estas especies u otros tipos de vegetación con anticipación si el área estuviera desprovista de estos (Landi & Renison 2010). También se pueden buscar lugares con piedras o colocarlas alrededor de las plántulas. Serán importantes lugares con suelos oscuros con presencia de humus o incorporar materia orgánica alrededor de las plántulas. El uso de micorrizas (Martino et al. 2011) o acido húmico (Chen & Aviad 1990) también podrían ayudar a mejorar el suelo.
Agradecimientos
Gracias a la Fundación Rufford Small Grants por su apoyo financiero a las iniciativas de restauración de los bosques de P lanata en el municipio de Independencia. A la comunidad de Pajchanti por ayudar en las reforestaciones. Al Ph. D. Daniel Renison, quién nos motivó a desarrollar la investigación y realizó recomendaciones sobre el manuscrito. Asimismo a las excelentes observaciones y recomendaciones del editor y revisores de la revista.
Referencias
Akaike, H. 1973. Information theory as an extensión of the máximum likelihood principle. pp. 267-281. En: Petrov, B.N. & F. Csaki (eds.). Second International Symposium on Information Theory. Akademiai Kiado, Budapest.
Anthelme, F. & O. Dangles. 2012. Plant-plant interactions in tropical alpine environments. Perspectives in Plant Ecology Evolution and Systematics 14: 363-372.
Ayma-Romay, A.I. 2012. Instituciones y autogobernanza del bosque común de una comunidad aymara en los Andes bolivianos. Tesis de maestría en áreas silvestres y conservación de la naturaleza, Universidad de Chile, Santiago. 98 p.
Ayma-Romay, A.I., R. Villavicencio, E. Rocha, N. Zambrana, F. Valdivia, O. Oseo, 2009. Bosques de Independencia: Pautas ecológicas y conocimiento local para el manejo tradicional. Fundación Rufford Small Grants y Gobierno Municipal de Independencia, Cochabamba. 120 p.
Bach, K., Fjelsá, S., M. Herzog, M. Kessler, J. Rasmussen & G. Swason. 1999. Cocapata and Sailapata: People and biodiversity in a bolivian mountain valley. Centre for Research on Cultural and Biological Diversity of Andean Rainforest, Rønde. 340 p.
Burnham, K., D. Anderson & K. P. Huyvaert. 2011. AIC model selection and multimodel inference in behavioral ecology: Some background, observations, and comparisons. Behavioral Ecology and Sociobiology 65: 23-35.
Calcagno, V. & C. de Mazancourt. 2010. glmulti: An R Package for Easy Automated Model Selection with Generalized Linear Models. Journal of Statistical Software 34 (12): 1-29.
Castro, J., R. Zamora, J. A. Hódar & J. M. Gómez. 2002. Use of shrubs as nurse plants: a new technique for reforestation in Mediterranean mountains. Restoration Ecology 10: 297-305.
Chen, Y. & T. Aviad. 1990. Effects of humic substances on plant growth. pp. 161-186. En: MacCarthy P, C. E. Clapp, R.L. Malcolm & P. R. Bloom (eds.) Humic Substances in Soil and Crop Sciences: Selected Readings. Soil Science Society of America, Madison.
Cierjacks, A., K. Wesche & I. Hensen. 2007. Potential lateral expansión of Polylepis forest fragments in central Ecuador. Forest Ecology and Management 242: 477-486.
Cierjacks, A., N. K. Rühr, K. Wesche & I. Hensen. 2008. Effects of altitude and livestock on the regeneration of two tree line forming Polylepis species in Ecuador. Plant Ecology 194: 207-221.
Dobson, A. J. 1990. An introduction to generalized linear models. Chapman & Hall, Londres. 115 p.
Enrico, L., G. Funes & M. Cabido. 2004. Regeneration of Polylepis australis Bitt. in the mountains of central Argentina. Forest Ecology and Management 190: 301-309.
Fjeldså, J. & M. Kessler. 2004. Conservación de la biodiversidad de los bosques de Polylepis de las tierras altas de Bolivia. Editorial FAN, Santa Cruz. 180 p.
Fowler, N. L. 1988. What is a safe site? Neighbor, litter, germination date and patch effects. Ecology 69: 947-961.
Fox, J. & Monette, G. 1992. Generalized collinearity diagnostics. Jasa 87:178-183.
Gómez-Aparicio, L., R. Zamora, J. M. Gómez, J. A. Hódar, J. Castro & E. Baraza. 2004. Applying plant facilitation to forest restoration: a meta-analysis of the use of shrubs as nurse plants. Ecological Applications 14:1128-1138.
Gosling, W. D., J. A. Hanselman, C. Knox, B. G. Valenda, & M.Bush. 2009. Long-term drivers of change in Polylepis woodland distribution in the central Andes. Journal of Vegetation Science 20:1041-1052.
Grueber, C. E., S. Nakagawa, R. J. Laws & I. G. Jamieson. 2011. Multimodel inference in ecology and evolution. Journal of Evolutionary biology 24: 699-711.
Hurvich, C. & C. Tsai. 1989. Regression and time series model selection in small samples. Biometrika 76: 297-307.
Johnson, J. B. & K. S. Omland. 2004. Model selection in ecology and evolution. Trends in Ecology and Evolution 19 (2): 101 -108.
Kessler, M. & Schmidt-Lebuhn. 2005. Taxonomical and distributional notes on Polylepis (Rosaceae). Organisms, Diversity & Evolution 6: 67-69.
Kessler, M, J. Bóhner & J. Kluge. 2007. Modelling tree height to assess climatic conditions at tree lines in the Bolivian Andes. Ecological Modelling 207: 223-233.
Kleier, C. & J. Lambrinos. 2005. The importance of nurse assodations for three tropical alpine life forms. Arctic, Antarctic and Alpine Research 37: 331-336.
Körner, C. & J. Paulsen. 2004. Aworld-wide study of high altitude treeline temperatures. Journal of Biogeography 31: 713-732.
Landi, M.A. & D. Renison. 2010. Forestación con Polylepis australis en suelos erosionados de las sierras grandes de Córdoba. Ecologia austral 20: 47-55.
Marcora, P. I., D. Renison, A. I. País-Bosch, M. R. Cabido & P. A. Tecco. 2013. The effect of altitude and grazing on seedling establishment of woody species in central Argentina. Forest Ecology and Management 291: 300-307.
Martino, J., C. Urcelay & D. Renison. 2011. Crecimiento y colonización micorrícica de Polylepis australis Bitter (Rosaceae) en suelos con distinta historia de pastoreo. Kurtziana 36: 69-77.
Meneses, R. & S.Beck. 2005. Especies amenazadas de la flora de Bolivia. Herbario Nacional de Bolivia. La Paz. 34 p.
Navarro, G. 2005. Provincia biogeográfica de los Yungas, pp. 279- 339. En: Navarro, G. & M. Maldonado (eds.). Geografía ecológica de Bolivia. Centro de Ecología y Difusión Simón I. Patino, Santa Cruz.
Peters, E. M., C. Martorell & E. Ezcurra. 2008. Nurse rocks are more important than nurse plants in determining the distribution and establishment of Globose cacti (Mammillaria) in the Tehuacán Valley, México. Journal of Arid Environments 72: 593-601.
Pimentel, L. 2007. Sistemas manuales de preparación del terreno con fines forestales. Universidad Autónoma de Chapingo. Chapingo. 49 p.
R Core Team, 2014. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. http://www.R-project.org/. [ Links ]
Renison, D., A. M. Cingolani & D. Schinner. 2002. Optimizing restoration of Polylepis australis woodlands: when, where and how to transplant seedlings to the mountains. Ecotropica 8: 219-224.
Renison, D., I. Hensen & A. M. Cingolani. 2004. Anthropogenic soil degradation affects seed viability in Polylepis australis mountain forests of central Argentina. Forest Ecology and Management 196: 327-333.
Renison, D., A. M. Cingolani, R. Suarez, E. Menoyo, C. Coutsiers, A. Sobral & I. Hensen. 2005. The restoration of degraded mountain woodlands: Effects of seed provenance and microsite characteristics on Polylepis australis seedling survival and growth in central Argentina. Restoration Ecology 13:129-137.
Renison, D., I. Hensen, R. Suarez & A. M. Cingolani 2006. Cover and growth habit of Polylepis woodlands and shrublands in the mountains of central Argentina: human or environmental influence? Journal of Biogeography 33: 876-887.
Renison, D., G. A. E. Cuyckens, S. Pacheco, G. F. Guzmán, H. R. Grau, P. Marcora, G. Robledo, A. M. Cingolani, J. Domínguez, M. Landi, L. Bellis & I. Hensen. 2013. Distribución y estado de conservación de las poblaciones de árboles y arbustos del género Polylepis (Rosaceae) en las montañas de Argentina. Ecología austral 23: 27-36.
Simpson, B., 1986. Speciation and spedalization of Polylepis in the Andes, pp. 304 - 316. En: Vuillemier, F. & M. Monasterio (eds.). High altitude tropical biogeography. Oxford University Press, Nueva York.
Smith, W.K., M. J. Germino, D. M. Johnson & K. Reinhardt. 2009. The altitude of alpine treeline: A bellwether of climate change effects. Botanic Review 75:163-190.
Stevenson, F. J. 1994. Humus chemistry: Génesis, composition, reactions. Wiley, New Jersey. 512 p.
Symonds, M. & A. Moussalli. 2011. A brief guide to model selection, multimodel inference and model averaging in behavioural ecology using Akaike's information criterion. Behavioral Ecology and Sociobiology 65:13-21.
Torres, R.C., D. Renison, I. Hensen, R. Suarez & L. Enrico. 2008. Polylepis australis' regeneration niche in relation to seed dispersal, site characteristics and livestock density. Forest Ecology and Management 254: 255-260.
Underwood, A. J. 1997. Experiments in ecology: their logical design and interpretation using analysis of variance. Cambridge University Press, Reino Unido. 68 p.
Williams, J., W. Gosling, S. Brooks, A. Coe & C. Xu. 2011. Vegetation, climate and fire in the eastern Andes (Bolivia) during the last 18,000 years. Palaeogeography Palaeoclimatology, Palaeoecology 312: 115-126.
Zuur, A. E. Leño & C. Elphick. 2010. A protocol for data exploration to avoid common statistical problems. Methods in Ecology and Evolution 1: 3-14.
Artículo recibido en: 29 de Agosto de 2014.
Manejado por: Ramiro López.
Aceptado en: 25 de Marzo de 2015.