










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkEcología en Bolivia
Print version ISSN 1605-2528On-line version ISSN 2075-5023
Ecología en Bolivia vol.49 no.2 La Paz Sept. 2014
Nota
Legitimidad en la dispersión de semillas de algarrobo (Prosopis flexuosa, Fabaceae) por zorro andino (Lycalopex culpaeus, Canidae) en el Valle de La Paz (Bolivia)
Legitimacy of algorrobo (Prosopis flexuosa, Fabaceae) seed dispersal by Andean fox (Lycalopex culpaeus, Canidae) in the valley of La Paz (Bolivia)
Diego E. Maldonado1*, Luis F. Pacheco2 & Laura V. Saavedra1
1Carrera de Biología, Universidad Mayor de San Andrés, C/27 Cota Cota, Campus Universitario, La Paz, Bolivia,
*Autor de correspondencia: diegoeduardo_mv@hotmail.com
2Colección Boliviana de Fauna, Instituto de Ecología, Universidad Mayor de San Andrés, Casilla 10077, Correo Central, La Paz, Bolivia
La dispersión de semillas por vertebrados frugívoros frecuentemente implica interacciones mutualistas (Armesto et al. 1987, Castro et al. 1994) y suele ser una fase dentro de un ciclo complejo, cuyo resultado es fundamental para determinar la composición y mantenimiento de las comunidades vegetales (Wang & Smith 2002, Howe & Miriti 2004). La dispersión por endozoocoria puede incrementar las probabilidades de supervivencia y acelerar la germinación de las semillas (Howe & Smallwood 1982).
En el marco del estudio de animales dispersores de semillas deben evaluarse tres atributos: 1) legitimidad del dispersor 2) eficiencia del dispersor y 3) efectividad del dispersor (Bustamante et al. 1992, Bustamante & Canals 1995). La legitimidad se refiere a la viabilidad de las semillas que pasan por el tracto digestivo y puede cuantificarse como la proporción de semillas defecadas viables (Herrera 1989). La eficiencia suele cuantificarse como la proporción de semillas defecadas en sitios donde puedan germinar y establecerse (Reid 1989); mientras que la efectividad puede evaluarse como la proporción de plántulas reclutadas a través de la diseminación de semillas por un dispersor (Reid 1989).
Estudios sobre endozoocoria en carnívoros son escasos, aunque se conoce que estos pueden ser importantes dispersores, principalmente los cánidos (Cossios 2006). Por ejemplo, el zorro andino (Lycalopex culpaeus, Canidae) es un legítimo, pero ineficiente dispersor de semillas de Cryptocarya alba (Lauraceae) y Lithraea caustica (Anacardiaceae) (Bustamante et. al. 1992, León-Lobos & Kalin-Arroyo 1994); aunque actúa como dispersor legítimo y eficiente de Schinus molle (Anacardiaceae) (Castro et. al. 1994, Silva et al. 2005).
En este trabajo reportamos los resultados de dos pruebas que permitieron evaluar la viabilidad de semillas de Prosopis flexuosa (Fabaceae) encontradas en heces de zorro andino, recolectadas en Llacasa (16º39'01.59" S, 68º03'42.06" O), a altitudes entre 3.300-3.600 m, en el municipio de Mecapaca, en la provincia puneña mesofítica de la región andina tropical (Navarro & Ferreira 2007). Recolectamos 32 muestras de heces de zorro en octubre de 2013 y las guardamos en bolsas de papel. En 29 de las muestras identificamos restos vegetales y de esas obtuvimos 491 endocarpios con semillas de P. flexuosa en su interior y 388 semillas que no pudimos identificar. Descartamos 120 semillas por tener el endocarpio perforado, lo cual es signo de depredación por invertebrados. Finalmente, se extraviaron 57 semillas, por lo que utilizamos 316 endocarpios con semillas para las pruebas de viabilidad.
Seleccionamos al azar 52 endocarpios con semillas para la prueba de tetrazolio (Castro et al. 1994). Extrajimos manualmente el endocarpio de las semillas y las remojamos en agua por 24 horas, para retirar el epispermo y dejar al descubierto el embrión y que el tetrazolio pueda actuar. En este proceso y por fallas mecánicas (pérdida del embrión y quiebre de la semilla), se redujo el número a 41 semillas, las que se remojaron enteras en tetrazolio al 0.01% por 48 horas (Ruiz 2009). El 48.8% (20) de los embriones de semillas remojadas se tiñó de color rojo intenso, indicando viabilidad; 39% (16) no presentaron tinción y del 12.2% (5) restante no se pudo identificar el color de tinción. Sin embargo y aunque no estaba planeado, se conservaron las semillas en una caja petri y 82.92% (34) de las 41 semillas remojadas en tetrazolio germinaron, incluyendo 11 de las semillas que no se tiñeron y cuatro de aquellas que no se identificó la tinción (Tabla 1).

Si bien esta prueba es rápida, los resultados parecen ser poco precisos, posiblemente porque la determinación de viabilidad obedece a rasgos cualitativos (patrón e intensidad de la tinción; Ruiz 2009). En nuestro caso, la viabilidad detectada por la prueba de tetrazolio subestimó la viabilidad real y, por tanto, la importancia del zorro en la dispersión de P. flexuosa. Esto coincide con otro estudio que reporta subestimación de la viabilidad de semillas de Masdevallia solomonni (Orchidaceae) por la prueba de tetrazolio (Mamani-Sanchez 2013).
Como alternativa para evaluar viabilidad, se colocaron a germinar semillas con endocarpios encontradas en las heces y trozos de legumbres que se cortaron a partir de legumbres recolectadas al pie de diez árboles. Usamos cuatro tratamientos de germinación, el primero y segundo consistieron en endocarpios con semillas encontrados en las heces y semillas con trozos de legumbre proveniente de legumbres recolectadas al pie de diez árboles, respectivamente. El segundo tratamiento actuó como el primer grupo control. El tercer tratamiento estaba conformado por semillas provenientes de heces a las que se les retiró manualmente el endocarpio. El último tratamiento consistió en semillas que se sacaron de legumbres recolectadas al pie de los árboles, siendo éste el segundo grupo control. Pusimos las semillas a germinar sobre algodón en 48 cajas petri: 12 cajas/tratamiento y 11 semillas o trozos/caja, con excepción de la doceava en la que sólo colocamos 10, por carencia de semillas. Se obtuvo un total de 131 semillas por tratamiento. Las cajas se regaron una vez por semana, entre el 7 de noviembre y el 9 de diciembre de 2013.
Mediante una ANOVA unifactorial confirmamos diferencia en los porcentajes de germinación entre los cuatro tratamientos (F(3,44)= 16.87; P < 0.001). Una prueba a posteriori (Tukey) mostró mayor porcentaje de semillas germinadas en aquellos tratamientos con semillas libres de endocarpios (tratamientos 3 y 4; Fig. 1). En los mismos dos tratamientos (3 y 4), las semillas germinaron más rápido que las semillas de los tratamientos 1 y 2; las que no se despojaron de partes de la legumbre (Fig. 1). Estos resultados se explican simplemente con el hecho de que P. flexuosa tiene frutos indehiscentes.
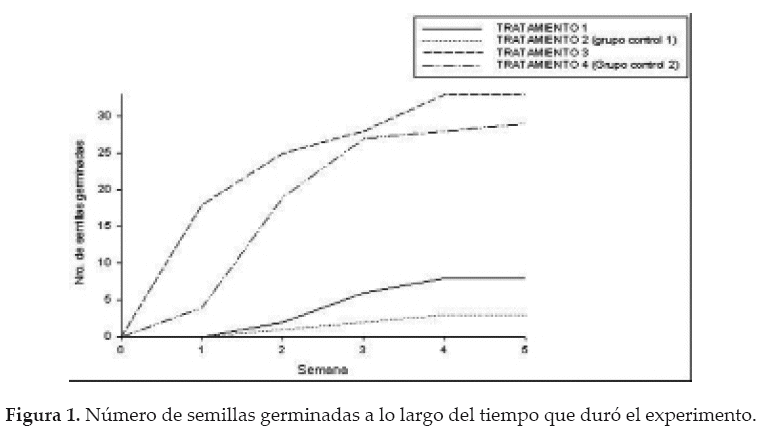
A pesar de que las semillas del tratamiento 1 estaban cubiertas por el endocarpio y las del tratamiento 2 mantenían el pericarpio, no se observó diferencia en el porcentaje de germinación entre estos tratamientos; siendo el endocarpio la parte de la legumbre que representa el principal obstáculo para la germinación de las semillas.
Por otro lado, los resultados indican que no existe diferencia en el porcentaje de germinación entre el tratamiento 1 y 2 y entre los tratamientos 3 y 4, lo que significa que el tracto digestivo del zorro no reduce la viabilidad de semillas de P. flexuosa. Un estudio en Argentina reporta que el zorro gris también defeca semillas viables de P. flexuosa y que estas semillas son defecadas con el endocarpio (Campos & Ojeda 2007).
Previo a nuestro trabajo, se documentó que el paso de semillas por el tracto digestivo del zorro andino tendría un efecto escarificante (por masticación o efecto de enzimas digestivas), que acelera la germinación de semillas de S. molle y C. alba (Castro et. al. 1994). En cambio, el efecto sobre semillas de Lithraea caustica sería lo contrario (Castro et al. 1994, León-Lobos & Kalin-Arroyo 1994). En nuestro caso, el paso de semillas P. flexuosa por el tracto digestivo del zorro no acelera la tasa de germinación, tal y como se ha registrado para el zorro gris en Argentina (Campos & Ojeda 2007).
En conclusión el zorro andino sería un legítimo dispersor para P. flexuosa (Fabaceae) en el Valle de La Paz, ya que la viabilidad de las semillas no es afectada negativamente por su paso a través de su tracto digestivo. Asimismo, es posible que, al consumir las legumbres, el zorro aleje las semillas de potenciales depredadores, que podrían actuar de manera denso-dependiente (Janzen 1970) y de esa manera generar un efecto positivo mayor al evidenciado en este trabajo.
Agradecimientos
Agradecemos a Mariana Da Silva por la colaboración en el reconocimiento de las heces y el material brindado. Emilia García colaboró en la identificación de las semillas. Jorge Quezada y Beatriz Mamani apoyaron para la prueba de tetrazolio. Los comentarios de Sergio Castro, Stella Giannoni y un revisor anónimo mejoraron este manuscrito.
Referencias
Alvarez, J.A. & P.E. Villagra. 2009. Prosopis flexuosa DC. (Fabaceae, Mimosoideae). Kurtziana 35 (1): 49-63.
Armesto, J.J., R. Rozzi, R Miranda & C. Sabag. 1987. Plant/frugivore interactions in South American temperate forests. Revista Chilena de Historia Natural 60(2): 321-336.
Burkart, A. 1976. A monograph of the genus Prosopis (Mimosoideae). Journal of Arnold Arboretum 57(3): 219-249.
Bustamante, R. O., J. A. Simonetti & J. E. Mella. 1992. Are foxes legitimate and efficient seed dispersers? A field test. Acta Oecologica 13(2): 203-208.
Bustamante, R. O. & M. Canals L. 1995. Dispersal quality in plants: how to measure efficiency and effectiveness of a seed disperser. Oikos 73(1): 133-136.
Campos, C. M & R. A. Ojeda. 1997. Dispersal and germination of Prosopis flexuosa (Fabaceae) seeds by desert mammals in Argentina. Journal of Arid Environments 35: 707-714
Castro, S. A, S. I. Silva, P. L. Meserve, J. R. Gutiérrez, L. C. Contreras & F. M Jaksic. 1994. Frugívora y dispersión de semillas de pimiento (Schinus molle) por el zorro culpeo (Pseudalopex culpaeus) en el Parque Nacional Fray Jorge (IV Región, Chile). Revista Chilena de Historia Natural 67(2): 169-176.
Cossios, E. D. 2006. Dispersión y variación de la capacidad de germinación de semillas ingeridas por el zorro costeño (Lycalopex sechurae) en el Santuario Histórico Bosque de Pómac, Lambayeque. Tesis de maestría, Universidad Nacional Mayor de San Marcos, Lima. 81 p.
Herrera, C. M. 1989. Frugivory and seed dispersal by carnivorous mammals and associated. OIKOS 55(2): 250-262.
Howe, H. F. & J. Smallwood. 1982. Ecology of seed dispersal. Annual Review of Ecology and Systematic 13: 201-228.
Howe, H. F. & M. N. Miriti. 2004. When seed dispersal matters. BioScience 54(7): 651-660.
Janzen, D. H. 1970. Herbivores and the number of tree species in tropical forests. American Naturalist 104(940): 501-528.
León-Lobos, P. M. & M. T. Kalin-Arroyo. 1994. Germinación de semillas de Lithraea caustica (Mol.) H. et A. (Anacardiaceae) dispersadas por Pseudalopex spp. (Canidae) en el bosque esclerófilo de Chile central. Revista Chilena de Historia Natural 67(1): 59-64.
Mamani-Sanchez, B. 2013. Germinación asimbiótica en Masdevallia solomonii a partir de diferentes tratamientos de polinización. Tesis de maestría en ecología y conservación, Universidad Mayor de San Andrés, La Paz. 66 p.
Navarro, G. & W. Ferreira. 2007. Mapa de vegetación de Bolivia, escala 1:250.000. Edición digital CD-ROM. The Nature Conservancy (TNC) & Rumbol, Santa Cruz.
Reid, N. 1989. Dispersal of mistletoes by honeyeaters and flowerpeckers: components of seed dispersal quality. Ecology 70(1): 137-145.
Ruiz, M. A. 2009. El análisis de tetrazolio en el control de calidad de semillas. Caso de estudio: cebadilla chaqueña. EEAINTA Anguil Argentina (77): 1-19.
Silva, S., F. Bozinovic, F. M. Jaksic. 2005. Frugivory and seed dispersal by foxes in relation to mammalian prey abundance in a semiarid thornscrub. Austral Ecology 30: 739-746.
Wang, B. C. & T. B. Smith.2002. Closing the seed dispersal loop. Trends in Ecology & Evolution 17(8): 379-384.
Nota recibida el: 07 de Julio de 2014.
Manejada por: Andrea Premoli.
Aceptada el: 01 de Septiembre de 2014.

