










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkEcología en Bolivia
Print version ISSN 1605-2528On-line version ISSN 2075-5023
Ecología en Bolivia vol.49 no.2 La Paz Sept. 2014
Artículo
Productividad interanual e interestacional de la biomasa aérea en la Chiquitanía transicional a la Amazonía (Guarayos, Bolivia)
Interseasonal and interannual productivity of above - ground biomass in a transition from Chiquitano to the Amazon (Guarayos, Bolivia)
Erwin Pardo-Toledo, Alejandro Araujo-Murakami, Luzmila Arroyo Padilla, Juan Pablo Heredia, Victor D. Rojas-Landivar, Marcio Flores-Valencia, Zamir Perez, Rebeca Sibler, Luz M. Mendizabal, Meison Vega-Martínez & Luzmarina Moreno
Herbario del Oriente Boliviano, Museo de Historia Natural Noel Kempff Mercado, Universidad Autónoma Gabriel René Moreno, Casilla 2489, Santa Cruz, Bolivia,
Email: erwinp1987@gmail.com, Autor de correspondencia
Resumen
Según investigaciones recientes, el cambio climático acentuará la estacionalidad con sequías prolongadas, lo cual repercutirá drásticamente sobre el almacenamiento de CO2 en los bosques amazónicos. Para determinar su impacto a menor escala, en el presente estudio se obtuvo los datos de densidad, altura total e incremento diamétrico mensual de árboles con DAP ≥ 10 cm en dos parcelas de 1 ha (100 x 100 m) en un bosque húmedo (BH) y bosque seco (BS) en Kenia, Guarayos (Santa Cruz, Bolivia). Estos datos se introdujeron a ecuaciones alométricas para estimar la biomasa y luego obtener la productividad de biomasa aérea (PBA), comparándola entre un año húmedo (2009) y uno seco (2010); asimismo se contrastó entre sus estaciones (húmeda, semiseca, seca, semihúmeda), además de analizar su relación con la precipitación pluvial y temperatura. La PBA interanual fue significativamente mayor en el año húmedo (BH = 40.16 kg/árbol/año; BS = 24.21 kg/árbol/año) respecto al año seco (BH = 35.97 kg/árbol/año; BS = 17.71 kg/árbol/año) (p < 0.05), mientras que la PBA interestacional fue mayor en la estación húmeda/2009 y la menor en la seca/2010, un escenario observado en ambos tipos de bosque. En cuanto a la relación de los factores climáticos con la PBA estacional, se evidenció que la precipitación pluvial tuvo una alta relación con la productividad (BH: rs = 0.91; p < 0.05); (BS: rs= 0.88; p < 0.05). Con lo que se deduce que los periodos con sequías prolongadas afectan significativamente la productividad de biomasa aérea, afirmándose que el clima juega un rol fundamental en la dinámica de estos bosques.
Palabras clave: Bosques, Clima, Productividad de biomasa aérea (PBA), Secuestro de carbono.
Abstract
Recent studies suggest that climate change will intensify seasonality resulting in prolonged droughts that may drastically reduce carbon storage in Amazonian forests. In order to determine drought impacts on a forest biomass at a local scale, this study allometric equations to estimate changes in tree and forest biomass based on wood density, tree height and monthly diameter growth of trees with DBH ≥ 10 cm in two plots of 1 ha (100 x 100 m) in a humid forest (BH) and a dry forest (BS) in Kenya, Guarayos (Santa Cruz, Bolivia). These estimates were used to compare biomass productivity (PBA) of a wet year (2009) and a dry one (2010). We also compared PBA among seasons (wet, semi-dry, dry, semi-wet seasons). The annual PBA was significantly higher during the wet year (BH = 40.16 kg/ tree/year; BS = 24.21 kg/tree/year) compared to the dry (BH = 35.97 kg/tree/year; BS = 17.71 kg/tree/year) (p < 0.05). Whereas for both types of forests the highest intraseasonal PBA was in the rainy season (2009) and the lowest in the dry one (2010). There were strong relationships between climatic factors with seasonal PBA, specifically, rainfall has a strong positive relationship with productivity BH (BH: rs = 0.91; p < 0.05); (BS: rs = 0.88; p < 0.05). We conclude that prolonged drought significantly reduces biomass productivity, and confirm that climate plays a fundamental role in the above-ground carbon dynamics of these forests.
Keywords: Above - ground biomass productivity (PBA), Climate, Forests, Carbon sequestration.
Introducción
Los bosques amazónicos abarcan un amplio rango climático, con altas precipitaciones en su margen noroeste hasta la estacionalidad en el extremo sur. En el presente, estos bosques son considerados prioritarios, por su papel como reservorios, además de secuestrar importantes cantidades de CO2, amortiguando el cambio climático. Sin embargo, se tienen evidencias que podrían acelerarlo al liberar grandes cantidades de carbono, debido a una mayor mortalidad natural de árboles, provocado por largo periodos de sequías, con lo cual se teme que los efectos contaminantes constituyan un panorama mundial preocupante (Phillips et al. 1998,2009).
Actualmente, se ha reportado que la periodicidad del crecimiento o productividad de los bosques tropicales, por lo general depende de acontecimientos climáticos como la precipitación pluvial, temperatura y humedad (Vargas & Valera 2007). Sin embargo, es difícil caracterizar, evaluar, trazar o predecir su variabilidad. Algunas especies crecen continuamente o al menos hasta cierto grado, mientras que otras exhiben distintos grados de periodicidad, por lo que es necesario medir el crecimiento en años completos (Wadsworth 2000).
Uno de los componentes del bosque más eficientes de medir es la biomasa aérea, definida como la cantidad total de materia orgánica desde la base del árbol, hasta el ápice (incluye follaje, ramas, troncos), y se expresa en peso seco (kg, tn) al horno por unidad de área (Brown 1997). Por lo cual, se considera a la productividad de biomasa aérea como un indicador relevante, a ser medido considerando las fluctuaciones espacio - temporales y explorando el grado de influencia de los factores que las controlan, con lo cual se puede avanzar en el entendimiento de la dinámica y la respuesta de los bosques ante los efectos del cambio climático (Malhi et al. 2004, RMIELP 2005, Mostacedo et al. 2008, Araujo et al. 2013).
En el presente estudio se analizó la variación de la productividad de biomasa aérea durante dos años (2009-2010) y la influencia de los factores climáticos en una zona del sector chiquitano transicional ala Amazonía. Debido a las particularidades microclimáticas, el estudio se enfocó en dos tipos de bosque, el húmedo (con características marcadas de bosques amazónicos) y el seco (con características de bosque seco semideciduos). Se aprovecharon las diferencias climáticas, con precipitaciones pluviales intensas en el 2009 y sequías en el 2010, una variación provocada por el fenómeno "La Niña" (SENAMHI 2010), las mismas que dieron un escenario propicio de análisis. Las preguntas de investigación son: ¿Cómo afectan los periodos de sequía a la productividad de la biomasa aérea?, la precipitación pluvial y la temperatura ¿influyen sobre la productividad de biomasa aérea?
Métodos
Área de estudio
La propiedad Kenia se encuentra al sureste de la provincia Guarayos, departamento de Santa Cruz, Bolivia. El clima es subhúmedo y húmedo mesotermal (Navarro y Maldonado 2002), con una precipitación anual promedio de 1.700 mm y una temperatura promedio de 23.4º C, según datos de la estación metereológica de Kenia (Flores-Valencia en prepa.), de la cual se tiene registro de datos climáticos.
Para efectos de comparación anual los datos fueron agrupados en 2009 (año húmedo), en donde se registró una precipitación pluvial de 1962.7 mm y una temperatura promedio de 23.3 ºC; mientras que en el 2010 (año seco), se tuvo una precipitación pluvial de 1.438 mm y una temperatura promedio de 23.5 ºC. Para la comparación estacional, se clasificaron los datos en función a la intensidad de lluvia que se da en los bosques de la zona (estación metereológica de Kenia), siendo agrupados en estación húmeda HUM (enero a marzo), semiseca S-SECA (abril a junio), seca como SECA (julio a septiembre) y semihúmeda S-HUM (octubre a diciembre). Estas diferencias climáticas fueron tomadas como criterios para agrupar y comparar los datos (Fig. 1).
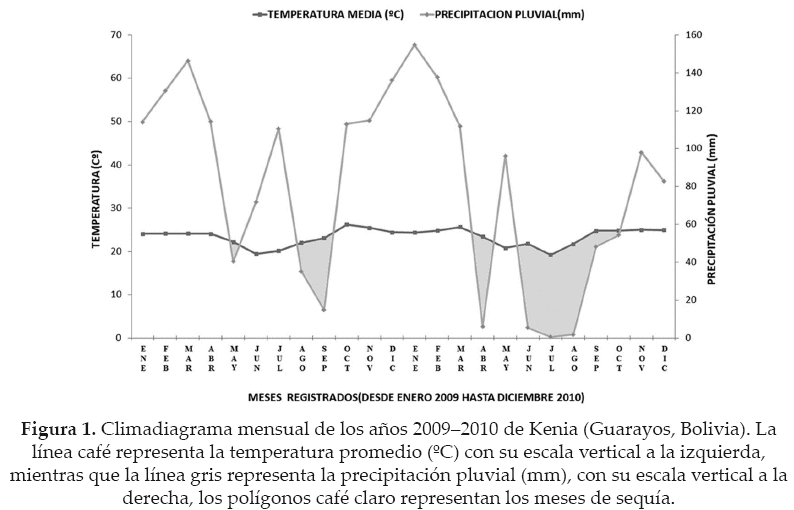
En la zona existen rastros de extracción selectiva de madera (principalmente de Swietenia macrophylla) durante la década de los 80's y 90's, tala con fines ganaderos, además del historial de incendios ocurridos años anteriores (Pinto & Alvarado 2007), lo cual aún se puede evidenciar en los sitios estudiados, dando además lugar a procesos de recuperación.
El área pertenece al sector chiquitano transicional a la Amazonía, presentando una combinación de elementos amazónicos y chiquitanos semideciduos (Navarro 2011). Se han identificado dos tipos de bosque: 1) bosque húmedo: caracterizado por especies siempreverdes, como Ficus glabrata, Cariniana ianeirensis, C. estrellensis, Hura crepitans y Pseudolmedia laevis, con un dosel entre 20-25 m de altura. Se desarrolla sobre suelos mesotróficos moderadamente ricos en bases intercambiables. Presenta un microclima con características de bosque amazónico mesofítico 2) bosque seco: con vegetación semidecidua, como ser Caesalpinia pluviosa, Anadenanthera colubrina, Piptadenia viridiflora y Centrolobium microchaete, con un dosel de 10-16 m de altura media, formado sobre suelos pedregosos expuestos, poco profundos y excesivamente drenados. Por sus características, estos suelos se desecan estacionalmente de forma más severa que los suelos más profundos, repercutiendo en una menor altura de dosel y mayor temperatura al interior del sotobosque, en relación al bosque húmedo mencionado anteriormente (Navarro 2011, Flores-Valencia en prep.).
Diseño de muestreo y toma de datos
Se utilizaron dos parcelas permanentes de monitoreo (PPM) instaladas en el 2008 por el personal del Museo de Historia Natural Noel Kempff Mercado, como parte del proyecto Red Amazónica de Inventarios Forestales (RAINFOR). Las mismas presentaban un muestreo estratificado al azar, una en un bosque húmedo (PPM A) y otra en un bosque seco (PPM B), cada una con una superficie de 1 hectárea, es decir 100 x 100 m (Fig. 2).
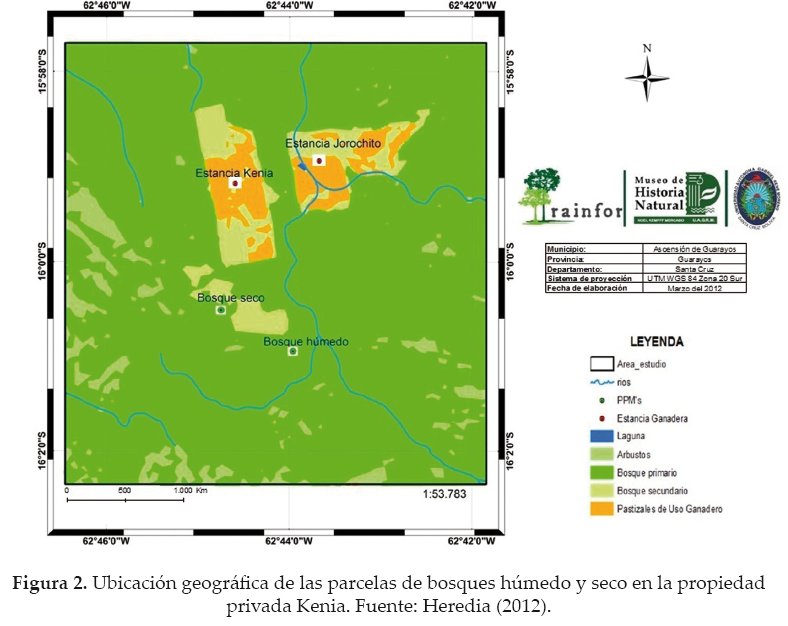
En las parcelas, a todos los individuos con DAP ≥ 10 cm se le instaló una banda dendrométrica (instrumento que se ajusta a la forma del tronco para detectar pequeños cambios en el diámetro del árbol, entre intervalos de medición), la misma que se encontraba a 1.4 m, 10 cm por encima del DAP. De forma mensual, se midió el incremento a todos los árboles, utilizándose un calibrador electrónico de alta precisión (centésima de milímetro) para detectar pequeños cambios en el crecimiento de los árboles y lianas a través de la banda dendrométrica, además de reclutar a nuevos individuos que entren a la categoría de diámetro ≥ 10 cm.
Se calculó la altura total en una oportunidad y se obtuvo la densidad de la madera de las especies, consultando la base de densidades de madera para los bosques amazónicos, publicada por Chave et al. (2006). Los datos climáticos de precipitación pluvial y temperatura fueron registrados de forma diaria de la estación meteorológica de RAINFOR, distante a aproximadamente 3 km de ambas parcelas. El levantamiento de datos fue realizado de forma continua, durante 24 meses, comenzando en enero del 2009 hasta diciembre de 2010. Todo el proceso de diseño de muestreo, toma y análisis de datos fue realizado siguiendo la metodología establecida por RAINFOR (Araujo et al. 2013).
Análisis de datos
Estimación de la biomasa aérea
Para los análisis estadísticos se utilizó el programa SPSS v.19, con un nivel de significancia del 95% (p < 0.05). Para obtener la biomasa aérea mensual por árbol, se consideró el diámetro a la altura del pecho, la densidad y la altura total como variables constantes, mientras que el incremento diamétrico fue la variable de diferenciación entre meses. Se utilizaron las siguientes ecuaciones (Tabla 1) de Chave et al. (2005) para árboles y la de Orrego & del Valle (2003) para lianas.

Evaluación de la PBA
Al obtenerse la biomasa aérea mensual de cada árbol, la tasa de aumento (crecimiento) de biomasa de cada mes se consideró como la productividad de biomasa aérea (PBA) mensual.
Comparación interanual de la PBA: Para evaluar la variación de la PBA ante diferentes escenarios climáticos, se agrupó la PBA mensual obtenida en el año húmedo (2009) y año seco (2010) obteniéndose la PBA anual, siendo contrastados con la prueba no paramétrica de Wilcoxon (Z).
Comparación interestacional de la PBA: Para evaluar la variación de la PBA en periodos de tiempo más cortos, se agrupó la PBA mensual según la estación climática (PBA estacional) e inferenciados con el análisis de varianza de medidas repetidas, junto al comparador de Bonferroni, el cual es usado para identificar diferencias estadísticas significativas entre grupos. Cabe resaltar que cada árbol se tomó como una unidad de muestreo. Se presenta la PBA de todos los individuos muestreados en cada parcela, como PBA total, tanto anual como estacional, los cuales fueron evaluados de manera descriptiva.
Dinámica y mortalidad de biomasa aérea
Para tener una visión general del comportamiento de la biomasa aérea, se estimó la dinámica anual de biomasa aérea, en cada parcela, siguiendo la presente ecuación:
BAF = BAI + Sobrev + Reclutas - Mortalidad
Donde, BAF = biomasa aérea final, BAI = biomasa aérea inicial, Sobrev = PBA en árboles sobrevivientes, Reclutas = biomasa de los árboles reclutas y Mortalidad=biomasa muerta.
La mortalidad de los individuos es traducida como pérdida de biomasa aérea para objeto de análisis. En el levantamiento de campo, un árbol se consideraba muerto, luego de una inspección en la cual se verificaba que sus tejidos internos están totalmente secos. Como las mediciones eran mensuales, se verificaba en cada monitoreo el estado de sanidad de los individuos. La pérdida de biomasa aérea por mortalidad, a través del tiempo, entre años y estaciones, fue analizada con el test Chi cuadrado (x2).
Relación de los factores climáticos con la PBA
Para evaluar la sensibilidad del bosque a los factores climáticos, se relacionó el cambio en la PBA estacional con la precipitación pluvial registrada durante dicho intervalo de tiempo, mientras que para la relación con la temperatura, esta fue promediada. Para su análisis se utilizó el coeficiente de correlación de Spearman.
Resultados
Evaluación de la PBA
Comparación interanual de la PBA
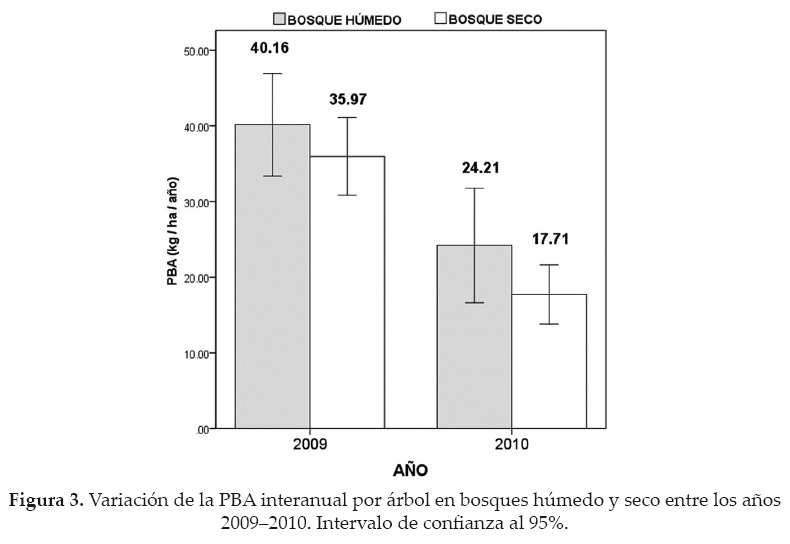
En la parcela de bosque húmedo, el número de árboles monitoreados mensualmente fue de 427, mientras que en la parcela de bosque seco, fueron 388. El análisis de varianza aplicado a la PBA interanual por árbol (Fig. 3), en el bosque húmedo, indica que la productividad fue significativamente mayor en el 2009 (año húmedo), con una media de 40.16 kg/árbol/año, en relación al 2010 (año seco) donde se obtuvo una media de 24.21 kg/árbol/año (Z = -7.39; p < 0.05). El mismo efecto se pudo evidenciar para el bosque seco, con diferencias significativas en la PBA entre ambos años, disminuyendo de 35.97 a 17.17 kg/árbol/año (Z = -11.64; p < 0.05).
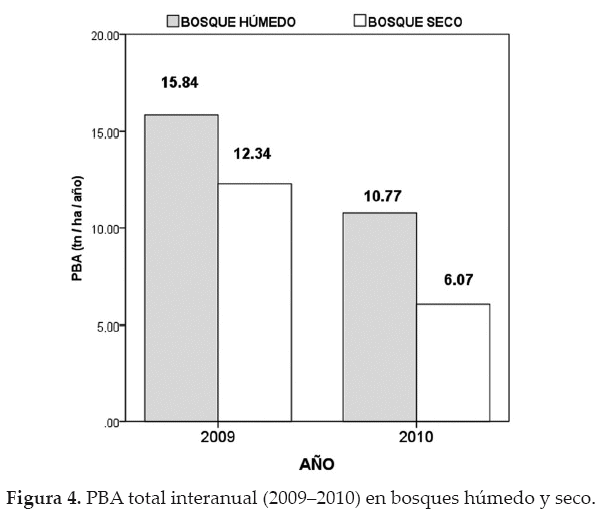
De forma concordante, la PBA total también tuvo una drástica reducción durante el periodo de sequía (Fig. 4), tanto en el bosque húmedo, con un descenso en la productividad que varió de 15.84 tn/ha/año (2009) a 10.77 tn/ha/año (2010); como en el bosque seco, con disminución de 12.34 tn/ha/año (2009) a6.07 tn/ha/año (2010).
Comparación interestacional de la PBA
Mediante la análisis inferencial, se determinó que la PBA interestacional por árbol en el bosque húmedo presentó diferencias significativas (F = 24.84; p < 0.05) entre las ocho estaciones, afirmando que los árboles presentaron su mayor productividad durante la estación húmeda del 2009 (19.29 kg/árbol/3 meses), siendo similar solamente con la estación húmeda del 2010 (13.01 kg/árbol/3 meses) (p = 0.09), ya que con las demás estaciones presenta diferencias significativas (p < 0.05). En contraposición, la menor PBA estacional por árbol se dio durante las estaciones secas del 2009 (2.22 kg/árbol/3 meses) y del 2010 (1.34 kg/árbol/3 meses), con similitud en su productividad (p = 0.90), pero con diferencias significativas con las otras estaciones (p < 0.05) (Fig. 5).
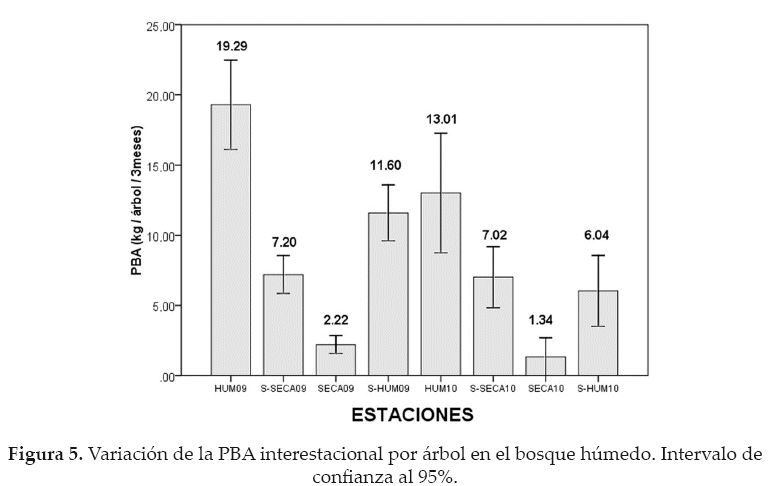
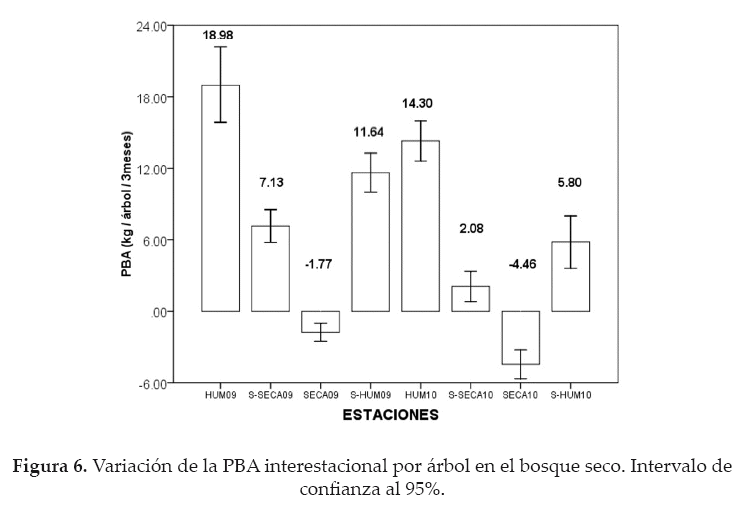
La PBA interestacional, por árbol en el bosque seco, presentó diferencias significativas (F = 57.14; p < 0.05). Los árboles alcanzaron su mayor productividad durante la estación húmeda del 2009 (18.98 kg/árbol/3meses), mostrando diferencias significativas con la productividad de las demás estaciones (p < 0.05); en contraste se pudo evidenciar que los árboles presentaron PBA estacionales negativas durante las estaciones secas del 2009 (-1.77 kg/árbol/3meses) y del 2010 (-4.46 kg/ árbol/3meses), con diferencias significativas entre sí y entre las demás estaciones (p < 0.05) (Fig. 6).
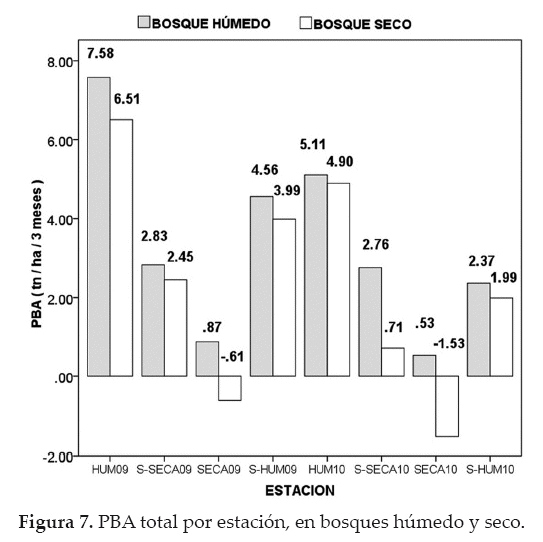
En ambos bosques, la mayor PBA total por estación se dio durante la estación húmeda del 2009, con 7.58 tn/ha/3 meses y 6.51 tn/ha/3 meses, respectivamente. Relativamente, ambos tuvieron su menor PBA total, durante la estación seca del 2010, con valores de 0.53 tn/ha/3 meses en el bosque húmedo y -1.53 tn/ha/3 meses en el bosque seco, lo cual se traduce en una reducción importante de la PBA en ambos tipos de bosques (Fig. 7).
Dinámica y mortalidad de biomasa aérea
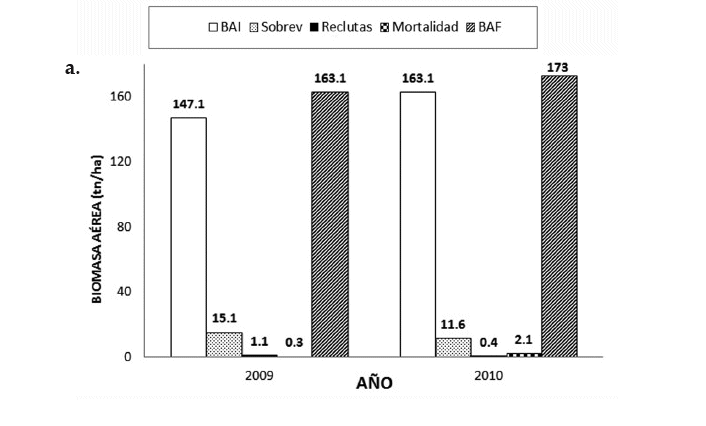
En el bosque húmedo, durante el 2009, la biomasa aérea en su primer medición presentó 147.1 tn/ha, lo que acumulado a PBA en árboles sobrevivientes, biomasa de árboles reclutas y la baja o pérdida por mortalidad, nos dio una biomasa aérea de 163.1 tn/ha, con un balance positivo durante ese año. Sin embargo, en 2010 la PBA en árboles sobrevivientes (11.6 tn/ha/año) como la biomasa de árboles reclutas (0.4 tn/ha) fue menor respecto al 2009. Hubo una mayor pérdida de biomasa aérea (2.1 tn/ha), no obstante, el balance general para el 2010 fue positivo, aunque menor en relación al anterior año (Fig. 8a).
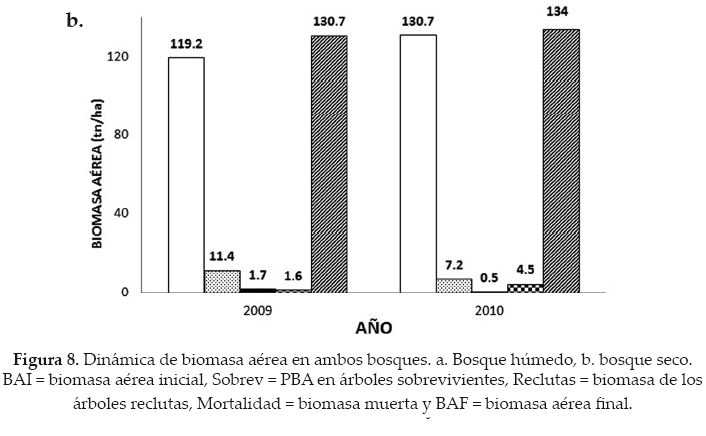
El bosque seco en el 2009 presentó una biomasa aérea en su primer medición de 119.2 tn/ ha, que junto a la PBA de árboles sobrevivientes y la biomasa de árboles reclutas, menos la pérdida de biomasa aérea por mortalidad, nos dio una biomasa aérea de 130.7 tn/ha. Al igual, que en el bosque húmedo, en el bosque seco la ganancia de biomasa fue menor en el 2010, ya que tanto la PBA de árboles sobrevivientes, como la biomasa de árboles reclutas disminuyó drásticamente, además de presentar una mayor pérdida por mortalidad. Sin embargo, se pudo observar un balance positivo con una biomasa final de 134 tn/ha (Fig. 8b).
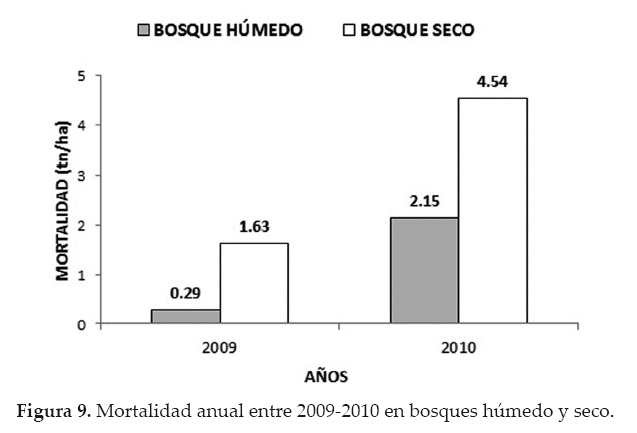
La pérdida de biomasa aérea por mortalidad, en el bosque húmedo, durante el 2009 fue de 0.29 tn/ha; sin embargo, esta cifra se elevó significativamente en el 2010 a 2.15 tn/ha (x2 = 6013.8; p < 0.05). También, en el bosque seco se observó el mismo efecto, es decir, una pérdida baja de biomasa aérea durante el 2009 (1.63 tn/ ha), aunque elevándose significativamente (x2= 6161.0; p<0.05) en el 2010 con 4.54 tn/ha (Fig. 9).
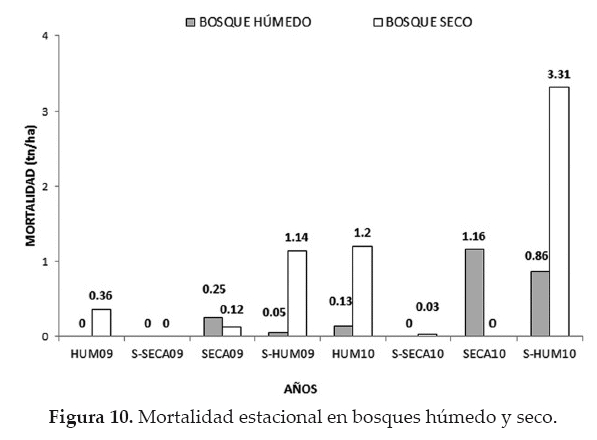
La mortalidad estacional en el bosque húmedo tuvo su mayores acumulaciones en la estaciones seca (1.16 tn/ha) y semihúmeda (0.86 tn/ha) del 2010, presentando diferencias significativas (x2 = 9780.0; p < 0.05) en la mortalidad entre las estaciones. La mortalidad estacional en el bosque seco fue más alta en las estaciones semihúmeda (3.31 tn/ha) y húmeda (1.20 tn/ha) de 2010, mostrando también diferencias significativas en la mortalidad entre las estaciones (x2 = 30815.0; p < 0.05) (Fig. 10).
Relación de los factores climáticos con la PBA
Bosque húmedo
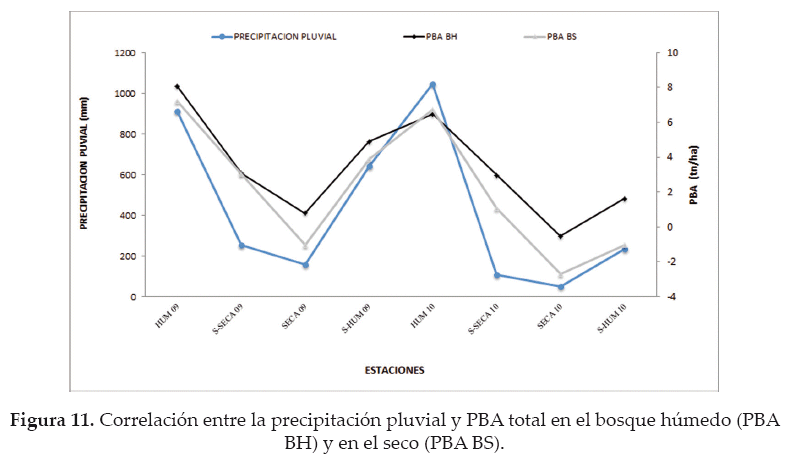
En el bosque húmedo, la relación entre la precipitación pluvial y la PBA total estacional fue estadísticamente significativa (rs = 0.91; p < 0.05); con una correlación fuerte y positiva (Fig.11). La relación entre la temperatura media y la PBA presenta una tendencia gráfica similar, sin embargo esta no presenta diferencias significativas (rs = 0.62; p = 0.09).
Bosque seco
La asociación entre la precipitación pluvial y la PBA total por estación, fue positiva, fuerte y significativa (rs = 0.88; p < 0.05), evidenciándose que la precipitación se relaciona con la PBA en el bosque seco (Fig. 11). De igual manera, en el bosque seco la asociación entre la temperatura media y la PBA estacional fue positiva, pero sin diferencias significativas (rs = 0.51; p = 0.20), demostrándose estadísticamente que estos factores son independientes.
Discusión
Productividad de biomasa aérea
La PBA interanual por árbol, para ambos tipos de bosque, se vio afectada por disminuciones significativas entre años, tal como lo afirma Phillips et al. (2009), quienes aseveran que los bosques amazónicos presentaron altas tasas de productividad de biomasa durante muchos años. Sin embargo, durante el periodo de sequía de 2005 provocado por el fenómeno de "El Niño" influyó para un descenso significativo en la productividad de biomasa, efecto que coincide con el descenso de productividad que se tuvo en 2010, producto de la sequía provocada por el fenómeno climático "La Niña". A pesar del descenso provocado por la sequía, ambos bosques mantuvieron un balance de productividad positivo.
Al analizar de forma específica, la PBA estacional por árbol presentó disminuciones significativas en ambos bosques, en especial durante las estaciones secas en el bosque seco, con valores negativos en la productividad, tal como se documenta en otros estudios (Baker et al. 2003, Soto 2007). Un efecto documentado por Valladares (2005) quien afirma que en condiciones de déficit hídrico los árboles y en especial los deciduos, pueden tener tasas de asimilación de CO2 cercanos a cero, además de una reducción en el transporte de agua en el xilema, inhibiendo el crecimiento y provocando plasmólisis celular, lo que se traduce en una reducción temporal del diámetro. Otro factor, que pudo haber influido en la productividad negativa de los árboles, son los suelos pedregosos, poco profundos y excesivamente drenados que presenta el bosque seco, lo que determina que estos bosques tengan menor altura, desecándose estacionalmente de forma más severa que en los suelos más profundos y menos pedregosos, como los bosques húmedos (Navarro 2011).
La PBA total por año en ambas parcelas presentó valores más altos en relación a los obtenidos para bosques amazónicos, como los de Baker et al. (2004), con una PBA total entre 0.8 y 2.36 tn/ha/año; Phillips et al. (2009) con 0.65-1.93 tn/ha/año o los de Araujo et al. (2006) con valores entre 2.65 tn/ha/año y 0.65 tn/ha/año. Esta productividad elevada puede deberse a un proceso de recuperación que presentan ambos bosques luego de los incendios (un acontecimiento periódico en los bosques de Guarayos), siendo el último registrado en 2004 (Pinto & Alvarado 2007) y que pudo ser constatado al evidenciarse rastros de fuego en algunos árboles estudiados. Sin embargo, se espera que a medida que el bosque vaya alcanzando su madurez o recuperándose de los efectos del incendio, irá disminuyendo sus tasas de productividad (Wadsworth 2000). Sumado a eso, el efecto fertilizante provocado por las altas concentraciones de CO2 presentes en la atmósfera, podría estar incidiendo levemente sobre el aumento de dichas tasas (Mahli et al. 1998, Phillips et al. 1998, Baker et al. 2004).
Dinámica y mortalidad de biomasa aérea
Mostacedo et al. (2008), estimaron una biomasa aérea entre 160-170 tn/ha (para un bosque subhúmedo y uno seco) valores que para el bosque húmedo guardan relación (173 tn/ha), aunque difiere del obtenido para el bosque seco (134 tn/ha). Un contraste atribuido a diferencias dasonométricas, ya que Mostacedo et al. (2008) presenta un bosque seco chiquitano maduro y sin efectos descritos de quemas, con valores mayores de área basal (19.7 m2/ha) y número de árboles (437 árboles/ha), en relación al investigado (17 m2/ha; 388 árboles/ha). Sin embargo, Araujo et al. (2006) dan resultados de 260.6 tn/ha para un bosque húmedo y 207.85 tn para un bosque seco en el Parque Nacional Noel Kempff Mercado, ambos valores mayores a lo obtenido en nuestra investigación, pero concordante con los resultados obtenidos por Dauber et al. (2000) guardan relación con lo obtenido por este estudio, con valores para la Amazonía (78-228 tn/ha) y para la Chiquitanía (73-157 tn/ha).
En el bosque húmedo y seco, la dinámica de biomasa aérea presentó cambios significativos entre años, con drásticos descensos de reclutamiento y productividad, con una mortalidad elevada, tanto en las estaciones como en el año seco, principalmente. De igual forma, Allen (2008) y Phillips et al. (2009) documentaron que durante las sequías del 2005 muchos bosques tropicales y principalmente el amazónico, evidenciaron marcadas disminuciones de crecimiento, bajos niveles de reproducción y altos tasas de mortalidad. De seguir en aumento estos fenómenos climáticos, los bosques amazónicos presentarán cambios importantes a nivel de comunidades establecidas, llegando a modificar de forma preocupante la composición de los individuos juveniles, además de invertir un sumidero de carbono a largo plazo (Valladares 2005, Phillips et al. 2009).
Relación de la productividad de biomasa aérea y los factores climáticos
La productividad de biomasa aérea mostró una buena relación con la precipitación pluvial en ambos bosques, lo cual concuerda con el estudio del proyecto FLUXNET (2010), quienes afirmaron que las precipitaciones son de vital importancia para los bosques a nivel mundial.
La falta de agua es el principal factor limitante de la productividad vegetal, ya que el estrés hídrico obliga al cierre estomático de las hojas con el fin de reducir la pérdida de agua, lo que lleva consigo a una reducción de la fotosíntesis y por lo tanto al de la productividad, por lo cual el crecimiento se concentra durante las estaciones lluviosas (Schulz 1960, Perez 2001).
Los resultados procesados en este estudio indican que a pesar de observarse una tendencia gráfica, la productividad de biomasa aérea es independiente a la influencia de la temperatura promedio, sin embargo, diversos estudios (Paruelo et al. 1999, Wadsworth 2000, Valladares 2005) sugieren que al variar la temperatura media, se ven afectados la respiración y fotosíntesis, por lo cual la producción primaria neta de los ecosistemas se altera de forma progresiva. De igual manera, los científicos del proyecto FLUXNET (2010) resaltan la importancia de la temperatura, ya que aseveran que a mayor temperatura las plantas emiten mayor CO2 por respiración. En este contexto, se hace necesario realizar evaluaciones locales con un rango temporal mayor, para poder tener un mejor entendimiento de este factor en nuestros bosques.
Conclusiones
La productividad de biomasa aérea de los árboles estudiados en Kenia (Guarayos, Bolivia) tanto en el bosque húmedo, como en el bosque seco, fue significativamente menor durante los años y estaciones secas. Asimismo, se evidenció que la precipitación pluvial tuvo una alta relación con la productividad. Por lo tanto, se puede afirmar que la precipitación pluvial es un factor importante para el crecimiento de los árboles, pudiendo determinar la magnitud de las tasas de producción en el área estudiada.
El área tiene un potencial importante como bosque secuestrador de carbono, determinado por sus altas tasas de productividad que varió de 15.84 tn/ha/año (2009) a 10.77 tn/ha/año (2010) en el bosque húmedo; y de 12.34 tn/ ha/año (2009) a 6.07 tn/ha/año (2010) en el bosque seco. Sin embargo, la reducción de la disponibilidad de agua por la intensa sequía es considerada como el principal responsable de la reducción de la productividad y una posible causa para el aumento de las tasas de mortalidad. Es decir, con estos resultados se evidencia la sensibilidad que tienen los bosques, pudiéndose convertir en fuentes temporales de emisión de CO2, aunque en menor magnitud que lo observado en otras investigaciones, ya que ambos bosques mantuvieron un balance positivo en las tasas de biomasa aérea, a pesar de los efectos de la sequía.
Agradecimientos
Esta investigación fue financiada por la Red Amazónica de Inventarios Forestales (RAINFOR). Nuestros agradecimientos a Marisol Toledo, Betty Flores y Teresa Ruiz de Centurión por sus importantes revisiones y sugerencias. Al Herbario del Oriente Boliviano (USZ) del Museo Noel Kempff Mercado por el apoyo logístico. Asimismo al Sr. René Castedo (Propietario) y a Pedro Tomicha y familia por permitirnos el trabajo en su propiedad y a todos los tesistas e investigadores del proyecto RAINFOR que participaron en la recolección de datos y aporte de ideas. Agradecemos a Mónica Moraes y a los revisores anónimos quienes contribuyeron en mejorar la versión final de este trabajo.
Referencias
Allen, C. D. 2008. Muerte regresiva del bosque inducida por el clima: ¿un fenómeno mundial en aumento?. Unasylva 231/232(60): 43-49.
Araujo, A., L. Arroyo, T. Killeen & M. Saldias. 2006. Dinámica del bosque, incorporación y almacenamiento de biomasa y carbono en el Parque Nacional Noel Kempff Mercado. Ecología en Bolivia 41(1): 24-45.
Araujo-Murakami, A., C. Doughty, D. Metcalfe, J. Silva-Espejo, L. Arroyo, J. Heredia M. Flores, R. Sibler, L. M. Mendizabal, E. Pardo-Toledo, M. Vega, L. Moreno, V. D. Rojas-Landivar, K. Halladay, C. A.J. Girardin, T. J. Killeen & Y. Malhi. 2013. The productivity, allocation and cycling of carbon in forests at the dry margin of the Amazon forest in Bolivia. Plant Ecology & Diversity DOI: 10.1080/17550874.2013.798364
Baker, T., D. Burslem & M. D. Swaine. 2003. Associations between tree growth, soil fertility and water availability at local and regional scales in Ghanaian tropical rain forest. Journal of Tropical Ecology 19:109-125.
Baker, T., O. Phillips, Y. Malhi, S. Almeida, L. Arroyo, A. Fiore, T. Erwin, N. Higuchi, T. Killeen, S. Laurance,W. Laurance, S. Lewis, A. Monteagudo, D. Neill, P. Núñez, N. Pitman, N. Silva & R. Vásquez. 2004. Increasing biomass in Amazonian forest plots. The Royal Society 359: 353-365.
Brown, S. 1997. Estimating biomass and biomass change of tropical forests. Forestry Paper, FAO, Roma. 55 p. [ Links ]
Chave, J., C. Andalo, S. Brown, M. A. Cairns, J. Q. Chambers, D. Eamus, H. Folster, F. Fromard, N. Higuchi, T. Kira, J. P. Lescure, B. W. Nelson, H. Ogawa, H. Puig, B. Riera & T. Yamakura. 2005. Tree allometry and improved estimation of carbon stocks and balance in tropical forests. Oecologia 145: 87-99.
Chave, J.C., H.C. Muller Landau, T.R. Baker, T. A. Easdale, H. Ter Steege & C. Webb. 2006 Regional and phylogenetic variation of wood density across 2,456 Neotropical tree species Ecological Applications 16: 235-236.
Dauber, E., J. Terán & R. Guzmán. 2000. Estimaciones de biomasa y carbono en bosques naturales de Bolivia. Superintendencia Forestal, Santa Cruz. 32 p.
Flores-Valencia, M. en prep. Estructura y composición de los bosques húmedos y seco semideciduo en el sector Kenia, Guarayos. Tesis de grado, Universidad Gabriel René Moreno, Santa Cruz.
FLUXNET 2010. A new balance for the global carbon balance. Max - Planck - Gesellschaft, Munich. http://www.mpg.de/616832/pressRelease201007041 (Visitado el 5 de enero de 2011). [ Links ]
Heredia, J. 2012. Productividad de necromasa vegetal e influencia del clima en dos tipos de bosques (Provincia Guarayos, Santa Cruz, Bolivia). Tesis de licenciatura en biología, Universidad Autónoma Gabriel René Moreno, Santa Cruz. 78 p. [ Links ]
Malhi, Y., T.R. Baker, O.L. Phillips, S. Almeida, E. Alvarez, L. Arroyo, J. Chave, C. Czimczik, A. Difiore, N. Higuchi, T.J. Killeen, S. G. Laurance, W.F. Laurance, S. L. Lewis, L. M. Mercado Montoya, A. Monteagudo, D. A. Neill, P. Nuñez Vargas, S. Patiño, N. A. Pitman, C. A. Quesada, R. Salomao, J. N. M. Silva, A. T. Lezama, R. V. Martinez, J. Terborgh, B. Vinceti & J. Lloyd. 2004. The above-ground coarse wood productivity of 104 Neotropical forest plots. Global Change Biology 10: 563-591.
Mostacedo, B., Z. Villegas, J.C. Licona, A. Alarcón, C .Leaño, M. Peña & L. Poorter. 2008. Dinámica de la biomasa en áreas de manejo forestal sujetas a diferentes intensidades de aprovechamiento. Documento Técnico Instituto Boliviano de Investigación Forestal (3): 1-43.
Navarro, G. & M. Maldonado.2002. Geografía ecológica de Bolivia: Vegetación y ambientes acuáticos. 4ta. Edic. Editorial Centro de Ecología Simón I. Patiño - Departamento de Difusión, Cochabamba. 718 p.
Navarro, G. 2011. Clasificación de la vegetación de Bolivia. Centro de Ecología Difusión Simón I. Patiño. Santa Cruz, 713 p.
Orrego, S. A. & J. I. del Valle. 2003. Existencias y tasas de incremento neto de la biomasa y del carbono en bosques primarios intervenidos y secundarios. pp 215-242. En: Orrego, S. A. J. I. del Valle & F. H. Moreno (eds.). Medición de la Captura de Carbono en Ecosistemas Forestales Tropicales de Colombia: Contribuciones para la mitigación del cambio climático. Universidad Nacional de Colombia sede Medellín, Centro Andino para la Economía en el Medio Ambiente, Bogotá. [ Links ]
Paruelo, J.M., W.K. Lauenroth, I.C. Burke & O.E. Sala. 1999. Grassland precipitation use efficiency across a resource gradient. Ecosystems 2: 64-69.
Pinto, C. & E. Alvarado. 2007. Efectos del fuego en bosque de producción forestal en Bolivia. Documento técnico Instituto Boliviano de Investigación forestal (IBIF), Santa Cruz. 24 p.
Phillips, O. L., Y. Malhi, N. Higuchi, W. Laurance, P. Núñez, M. Vásquez, S. Laurance, L. Ferreira, M. Stern, S. Brown & J. Grace. 1998. Changes in the carbon balance of tropical forest: evidence from long-term plots. Science 282: 439-442.
Phillips, O. L., E. Luiz Aragao, S. Lewis, J. Fisher, J. Lloyd, G. López-González, Y. Malhi, A. Monteagudo, J. Peacock, C. Quesada, G. Van Der Heiden, S. Almeida, L. Arroyo, G. Aymard, T. Baker, O. Banki, L. Blanc, D. Bonal, P. Brando, J. Chave, A. Alves De Oliveira, N. Dávila, C. Czimezik, T. Feldpausch, N. Higuchi, P. Meir, C. Mendoza, A. Prieto, F. Ramirez, M. Schwars, J. Silva, M. Silveira, J. Stropp, R. Vázques, P. Zelazowski, S. Andelman, A. Andrade, K. Chao, E.Honorio, T. Killen, W. Laurance, A. Peña, N. Pitman, P. Vargas, A. Rudas, R. Salamao, N. Silva, J. Terborgh & A. Torres-Lezama. 2009. Drought sensitivity of the Amazon rainforest. Science Magazine 323:1344-1347.
RMIELP (Red Mexicana de Investigación Ecológica a Largo Plazo). 2005. Patrones y control de la productividad primaria en los ecosistemas. Documento fundacional (fragmento). Versión preliminar, México D.F. 12 p. [ Links ]
Schulz, J.P. 1960. The vegetation of Surinam. pp 87-98. En: Schulz, J.P. (ed.) Ecological Studies on Rain Forest in Northern Surinam. Hoord-Hollandsche Vitgeners Maats-chappij, Ámsterdam.
SENAMHI (Servicio Nacional de Meteorología e Hidrología - Bolivia). 2010. Boletín informativo y pronóstico Nro. 2 sobre evolución de condiciones "La Niña" 2010 - 2011. Ministerio de Medio Ambiente y Agua, La Paz. 13 p.
Soto, D. 2007. Evaluación de la productividad de la Biomasa aérea arbórea en tres tipos de bosques en la reserva privada "Los Volcanes". Tesis de licenciatura en ingeniería forestal, Universidad Autónoma Gabriel René Moreno, Santa Cruz. 72 p.
Valladares, F. 2005. Ecología del bosque mediterráneo en un mundo cambiante. Ministerio de Medio Ambiente, Madrid. 587 p.
Vargas, L. & A. Varela. 2007. Producción de hojarasca de un bosque de niebla en la reserva natural La Planada (Nariñó, Colombia). Universitas scientiarum edición especial I,12: 35-49.
Wadsworth, F. H. 2000. Producción forestal para América tropical. USDA Forest Service, Washington DC. 603 p.
Artículo recibido el: 03 de Julio de 2014.
Manejado por: Mónica Moraes R.
Aceptado el: 17 de Septiembre de 2014.

