











 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkEcología en Bolivia
versión impresa ISSN 1605-2528versión On-line ISSN 2075-5023
Ecología en Bolivia v.48 n.1 La Paz abr. 2013
NOTA
Abundance and species richness of hawkmoths (Lepidoptera: Sphingidae) in the fragmented landscape of Santa Cruz (Bolivia)
Abundancia y riqueza de especies de polillas esfíngidas (Lepidoptera: Sphingidae) en el paisaje fragmentado de Santa Cruz (Bolivia)
Dirk Nikolaus Karger1, Stefan Abrahamczyk1,2 & Michael Kessler1
1Institute of Systematic Botany, University of Zurich, Zollikerstrasse 107, 8008 Zurich, Switzerland, Email: dirk.karger@systbot.uzh.ch Author for correspondence
2Department of Biology, Ludwig Maximilian University Munich, Menzinger Strasse 67, 80638 Munich, Germany
The bosque chiquitano or Chiquitano dry forest in the Department of Santa Cruz (Bolivia) is one of the main ecoregions in the Bolivian lowlands. Although many taxa of the bosque chiquitano are also present in the neighbouring ecoregions of the Gran Chaco and the southwestern Amazon, it is clearly distinguishable by its unique floral and faunal composition and geo-ecological characteristics (Ibisch & Mérida 2004). Bolivia itself belongs to the countries with the highest levels of biodiversity and endemism worldwide (Ibisch & Mérida 2004). Despite its huge biodiversity and unique ecosystems, Bolivia is also threatened by rapid deforestation and conversion of natural landscapes. The area around the Bolivian city of Santa Cruz is especially considered one of the world's main hotspots of deforestation with rates reaching up to 117,000 ha annually (Morales 1996, Achard et al. 1998).
Hawkmoths (Sphingidae) are among the best surveyed Lepidoptera worldwide (Kitching & Cadiou 2000). Detailed information exists about the distribution and ranges for most species (Kitching & Cadiou 2000). There are ca. 1,350 hawkmoth species worldwide distributed on all continents and island groups with the exception of Antarctica. However, the highest diversity of hawkmoth is found in tropical biomes (Kitching & Cadiou 2000). Recent estimations of species richness indicate the presence of 189 species in Bolivia of which at least 111 species occur in the Department of Santa Cruz (Kitching et al. 2001). Hawkmoths are of high importance for the ecosystem acting as pollinators but also as important agricultural pests (Kitching & Cadiou 2000). The present work aims to describe the hawkmoth species richness from the bosque chiquitano's area of Santa Cruz.
We surveyed hawkmoth species richness around Santa Cruz city (17°47'01 S, 63°10'57 W) in the lowlands of Bolivia; at an elevation of 420-450 m. Study sites were located in five forest fragments in and around the Parque Regional Lomas de Arena and the Jardín Botánico Santa Cruz, their sizes are indicated in Table 1. The matrix around the forest fragments was mostly farmland and pasture, with slight deviations (partly experimental fields with different crops around the 137 ha forest fragment). To account for any influence of the forested area around the study sites, the forested area was measured using satellite images taken from Google Earth and georeferenced using Spatial Analyst in ESRI ARCVIEW 3.2a. This data was validated with GPS measurements from the field to ensure that satellite images reflected the conditions at the time of the study. Large tree fall gaps, fields or clearings >10 m in diameter within the fragments were subtracted from the forest area.
Hawkmoths were collected with a light trap consisting of two 20 W tubular fluorescent light tubes with either two fluorescent ultraviolet or fluorescent white light tubes. Lamps were connected to a 12 V car battery powered by a car engine via a voltage converter to ensure stable voltage output. Lamps where placed 50 cm above ground and 1.5 m in front of a white cloth sheet (3x2 m). Additionally, a white cloth sheet (3 x 2 m) was placed on the ground in front of the vertical sheet. The fluorescent light tubes were placed so that most of the light was directed towards the sheets. This installation was placed at the forest edge with the white sheet facing the forest so that only the light reflected by the sheets was visible from the forest. On the side of the sheet facing away from the forest we placed the car to stabilize the trap under windy conditions and to shield the light towards the surrounding habitat matrix.
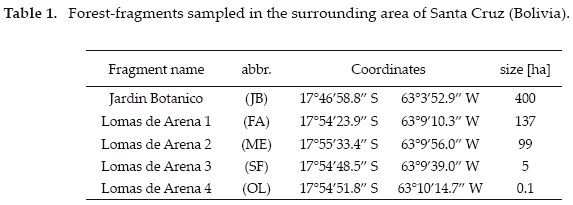
All fragments were surveyed for six days between 18:00 h and 24:00 h, three days each using the ultraviolet and white lamps, respectively. All Sphingidae attracted to the light-trap were caught. Hawkmoths were killed using injections of 1-5 ml of 99% ethanol solution into the metathorax. Location and flight time were noted and the moths were stored for later preparation and species determination. Identifications were made with reference to illustrations and identification keys (D'Abrera 1986, Moré et al. 2005). Nomenclature and classification follows Kitching & Cadiou (2000). The species-area relationship (number of species/fragment size) was analyzed using linear regression in R 2.9.1 (R Development Core Team 2009). Total species richness was estimated using EstimateS (Colwell 2005) (Table 2).
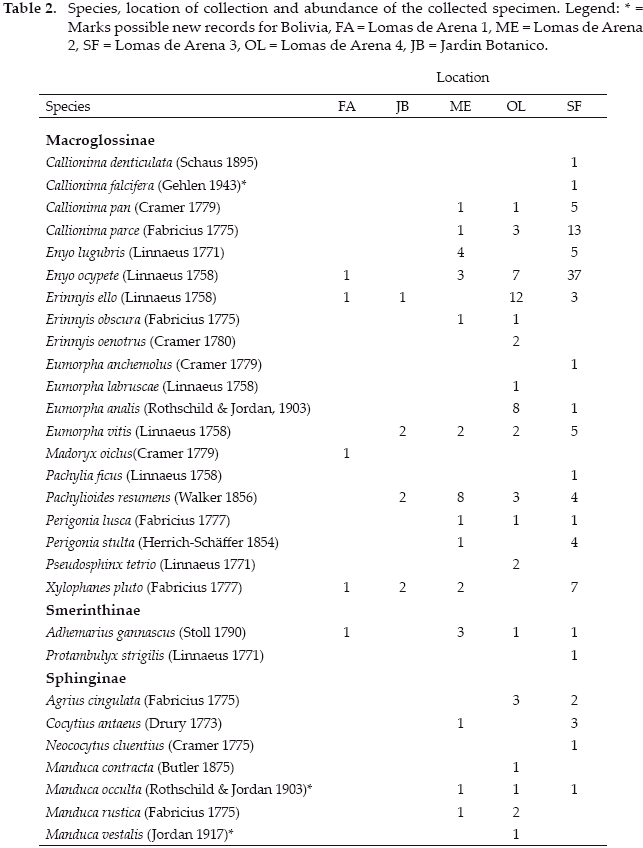
In total, we collected 194 hawkmoths belonging to 29 species and three subfamilies (Table 2). The forest fragment SF showed the highest richness and abundance of Sphingidae while the forest fragment FA showed the lowest abundance and richness (Figs. 1a-b). Macroglossinae formed the major subfamily of Sphingidae at all sampling sites accounting for the majority of species and individuals. Sphinginae were only present at the sampling sites ME, OL, SF while Smerinthinae were only absent at the sampling site JB.
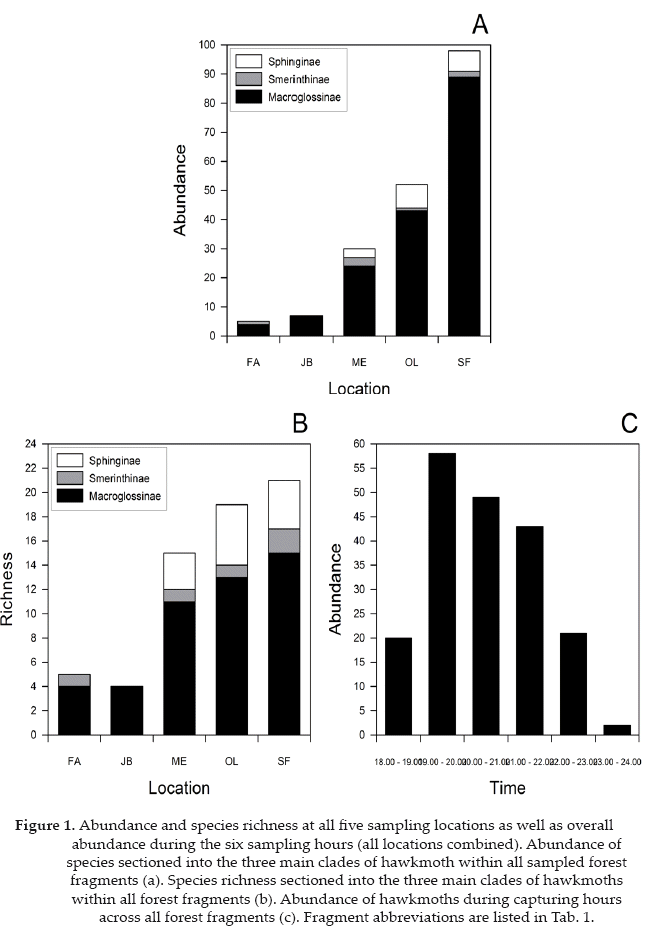
Most species were captured between 19:00 h and 20:00 h. Towards midnight there was a steady decline in individuals captured, with the minimum number between 23:00 h and 24:00 h (Fig. 1c). Abundance of species also steadily declined from 18:00 h to 24:00 h, with the lowest amount of species flying between 23:00 h and 24:00 h.
The number of species decreased with the area of forest around the sampling sites, although this pattern was only marginally significant (linear regression, R2 0.70, p = 0.078) (Fig. 2). However, when we corrected species numbers for uneven sampling intensity between the fragments, the species richness-fragment size relationship became significant (linear regression, R2 = 0.81, p = 0.038). The number of individuals strongly decreased with increasing forest area, but the relationship was not significant (linear regression, R2 = 0.50, p= 0.18).
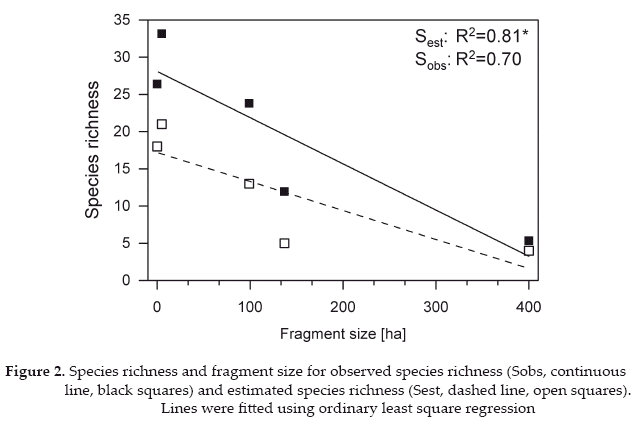
In our rapid assessment, we were able to collect around 26% of the known 112 hawkmoth species from Santa Cruz department (Kitching et al. 2001). This represents only a small fraction of the overall diversity of hawkmoth in the department as noted by Kitching et al. (2001). The low amount of species captured might be caused by the limited timeframe of the study, which did not allow us to capture hawkmoths flying during the rainy season or are more active in the mornings. Three of the species collected: Callionima falcifera (Gehlen 1943), Manduca occulta (Rothschild & Jordan 1903), and Manduca vestalis (Jordan 1917) have not been recorded for Bolivia before (Kitching et al. 2001). The number of hawkmoth species declined markedly from the small to the large forest fragments (Fig. 2). This pattern contradicts the typically observed decline of species richness with increasing fragmentation (e.g., Klein 1989, Creighton et al. 2009), although some other studies have also found increasing numbers for other groups than Sphingidae (e.g., hummingbirds: Stouffer & Bierregaard 1995, polyphagous lepidoptera: Tscharntke et al. 2002). Probably, these differences reflect the degree to which the studied taxon depends on forest habitats. Forest specialists, particularly if they have difficulty moving between fragments, will show negative richness-fragment size relationships, whereas species that can also or even preferentially survive in open habitats, may be favored by forest fragmentation.
Our observations may be explained as a result of the ecology of hawkmoths. Sphingids are strong flyers that forage widely in search of food resources. Forest edges and disturbed forests tend to have a higher abundance of f lowering plants than closed forests (Linhart et al. 1987). Accordingly, small forest fragments are likely to have more food resources for hawkmoths than large patches. Therefore, it might be the case that most of the species caught are mostly non forest species usually living in open habitats, which only forage in the forest.
Previous studies have found limited responses of sphingids to forest degradation (Schulze & Fiedler 2003, Beck et al. 2006). Beck et al. (2006) concluded that species that do not feed as adults are more susceptible to forest degradation than those that feed as adults. In our study, all species recorded belong to the latter group, which may explain the observed pattern. Nevertheless, this does not explain why there are so few species within the large forest fragments. Other factors might play a role here, but due to the limited data we are currently unable to determine any ecological explanation of the observed pattern.
We found that forest fragmentation in our study area does not lead to a reduction of hawkmoth abundances and diversity; rather, it increases in small fragments. We believe that this mainly reflects the high mobility of these large insects and the higher abundance of their food plants along forest edges and in disturbed forests. It must be borne in mind, however, that our study was limited to adults. To which degree these species depend on forest-inhabiting plants as larvae, and hence, how forest fragmentation negatively influences them, needs further investigation.
Acknowledgments
We thank Steffen Reichle, Yuvinca Gareca, Caroli Hamel, Sebastian K. Herzog, Dirk Embert, and Torrey Rodgers for help with the fieldwork. The Botanical Garden of Santa Cruz de la Sierra, Dario Melgar, Fernando Mercardo Bowles, Christian Bomblat, and The Fundación Simón F. Patiño for allowing fieldwork in their premises; and Jürgen Kluge and Rodrigo W. Soria Auza for valuable comments on the manuscript.
References
Achard, F., Eva, H.A. Glinni, P. Mayaux, T. Richards, &H. J. Stibig. 1998. Identification of deforestation hot spot areas in the humid tropics, TREES Series B, European Commission, Luxembourg.100p. [ Links ]
Beck, J., I. Kitching & K. Linsenmair. 2006. Effects of habitat disturbance can be subtle yet significant: Biodiversity of hawkmoth-assemblages (Lepidoptera: Sphingidae) in Southeast-Asia. Biodiversity and Conservation 15: 465-486.
Colwell, R. K. 2005. EstimateS, Version 7.5, statistical estimation of species richness and shared species from samples (Software and User's Guide). Freeware published at http://viceroy.eeb.uconn.edu/estimates [ Links ]
Creighton, J. C., R. Bastarache, M. V. Lomolino & M.C. Belk. 2009. Effect of forest removal on the abundance of the endangered American burying beetle, Nicrophorus americanus (Coleoptera: Silphidae). Journal of Insect Conservation 13:37-43.
D'Abrera, B. 1986. Sphingidae mundi. Hawkmoths of the world. E. W. Classey, Faringdon. 226 p. [ Links ]
Ibisch, P.L. & G. Mérida (eds.) 2004. Biodiversity: the richness of Bolivia. State of knowledge and conservation. Ministry of Sustainable Development. Editorial FAN, Santa Cruz. 644 p. [ Links ]
Klein, B .C. 1989. Effects of forest fragmentation on dung and carrion beetle communities in Central Amazonia. Ecology 70: 1715-1725.
Kitching, I.J. &J. M. Cadiou. 2000. Hawkmoths of the world: an annotated and illustrated revisionary checklist (Lepidoptera: Sphingidae). Cornell University Press, Ithaca. 227 p.
Kitching, I. J., J. Ledezma & J. Baixeras. 2001. The Sphingidae of Bolivia (Insecta: Lepidoptera). Gayana (Concepción) 65: 79-111.
Linhart, Y., P. Feinsinger, J. Beach, W. Busby, K. Murray, W. Pounds, S. Kinshman, C. Guindon, & M. Kooiman. 1987. Disturbance and predictability of flowering patterns in bird-pollinated cloud forest plants. Ecology 68: 1696-1710.
Morales, I. 1996. Memoria explicativa del monitoreo preliminar del desbosque en el Departamento de Santa Cruz. CORDECRUZ, Santa Cruz.100 p. [ Links ]
Moré, M., I.J. Kitching & A. Cocucci.2005. Sphingidae: Esfíngidos de Argentina. Hawkmoths of Argentina.L.O.L.A. (Literature of Latin America), Buenos Aires, 166 p. [ Links ]
R Development Core Team, 2008. R: A language and environment for statistical computing. R Foundation for Statistical Computing. Vienna, Austria. http://www.R-project.org [ Links ]
Schulze, C. &K. Fiedler. 2003. Hawkmoth diversity in northern Borneo does not reflect the influence of anthropogenic habitat disturbance. Ecotropica 9:99-102.
Stouffer, P. & R. Bierregaard. 1995. Effects of forest fragmentation on understory hummingbirds in Amazonian Brazil. Conservation Biology 9:1085-1094.
Tscharntke, T., I. Steff an-Dewenter, A. Kruess& C. Thies. 2002. Characteristics of insect populations on habitat fragments:Amini review. Ecological Research 17:229-239.
Nota recibida en: Noviembre de 2012.
Manejada por: Diana Silva.
Aceptada en: Enero de 2013.

