










Services on Demand
Journal

Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
 Cited by SciELO
Cited by SciELO Related links
 Similars in
SciELO
Similars in
SciELO  uBio
uBio Share

 Permalink
PermalinkEcología en Bolivia
Print version ISSN 1605-2528On-line version ISSN 2075-5023
Ecología en Bolivia vol.48 no.1 La Paz Apr. 2013
Ranging patterns by the red-footed tortoise - Geochelone carbonaria (Testudines: Testudinidae) - in the Bolivian Chaco
Patrones de desplazamiento de la peta negra - Geochelone carbonaria (Testudines: Testudinidae) - en el Chaco boliviano
Rossy R. Montaño F.1, Erika Cuéllar2, Lee A. Fitzgerald3, Filemón Soria4, Florencio Mendoza4, Romoaldo Peña4, Telmo Dosapey4, Sharon L. Deem5 & Andrew J. Noss*6
1 Museo de Historia Natural Noel Kempff Mercado, Universidad Autónoma Gabriel René Moreno, Casilla 2489, Santa Cruz, Bolivia, rovina7@yahoo.com
2Wildlife Conservation Research Unit, Zoology, University of Oxford, England, erika.cuellar71@gmail.com
3Department of Wildlife and Fisheries Sciences, Texas A&M University, College Station, Texas, USA, lfitzgerald@tamu.edu
4Capitanía de Alto y Bajo Isoso, Casilla 3108, Santa Cruz, Bolivia
5Institute for Conservation Medicine, Saint Louis Zoo, One Government Drive, Saint Louis, Missouri, USA, deem@stlzoo.org
*6Corresponding author: University of Florida, Department of Geography, P.O. Box 117315, Gainesville, FL 32611, 352-338-9016, anoss@ufl.edu
Abstract
In the Bolivian Chaco, the tortoise Geochelone carbonaria is an important reptile to indigenous people for subsistence purposes and in traditional medicine. In Bolivia the species is considered near threatened, while observations suggest it is less abundant near communities and cattle ranches. However, understanding of its biology and ecology is limited. As part of a landscape conservation program with indigenous land and wildlife management, this article describes research on ranging patterns for the species at two long-term research camps in the Kaa-Iya del Gran Chaco National Park. The principal research method is radio-telemetry with 15 individuals over a two-year period. The tortoises occupy home ranges of 50-600 ha, with males using ranges three times larger than females. Individual home ranges overlap, up to 96% between pairs of monitored individuals, with multiple individual ranges overlapping simultaneously, while many males and females shift their ranges between wet and dry seasons. The relatively large ranges for an animal of this size in the Chaco, combined with shifting ranges across seasons, even in protected areas, means that conservation of red-footed tortoises within human-impacted landscapes will require provision of key resources that include appropriate food, water, and shelter where these become scarce. Alternatively, indigenous territories and ranches will need to set aside communal and private reserves exceeding 1,000 ha in order to maintain tortoise populations on their lands.
Key words: Bolivia, Home Range, Kaa-Iya, Radio-Telemetry, Reptiles, Seasonal.
Resumen
En la región del Chaco boliviano la tortuga negra Geochelone carbonaria es un reptil importante para los pueblos indígenas por motivos de subsistencia y medicina tradicional. Sin embargo, el conocimiento de aspectos biológicos y ecológicos es muy pobre. El presente artículo describe áreas de acción para la especie en dos campamentos de investigación a largo plazo del Parque Nacional Kaa-Iya del Gran Chaco. El método principal empleado ha sido la radio-telemetría en quince individuos durante dos años. Las tortugas ocuparon áreas de acción de 50-600 ha, con territorios ocupados por machos unas tres veces más grandes que los territorios ocupados por hembras. Existe solapamiento territorial entre individuos de ambos sexos, hasta 96% entre individuos observados, con múltiples individuos solapados entre sí, y tanto machos como hembras desplazan sus áreas de acción entre épocas seca y húmeda. Las áreas de acción relativamente grandes para un animal de este tamaño en el Chaco, en combinación con cambios de áreas de acción entre época seca y húmeda, inclusive dentro de áreas protegidas, indica que la conservación de la tortuga negra en paisajes impactados por humanos requerirá de la provisión de recursos claves como son alimentos, agua y refugios adecuados cuando estos mismos escaseen. Como alternativa, los territorios indígenas y las propiedades ganaderas deberán resguardar reservas comunales y privadas superando las 1.000 ha para poder mantener poblaciones de tortugas en sus tierras.
Palabras clave: Área de acción, Bolivia, Estacional, Kaa-Iya, Radio-telemetría, Reptiles.
Introduction
Tortoises worldwide are under threat from a variety of human pressures, for example hunting for meat or eggs by humans, or by their domestic and feral animals; the pet trade; habitat loss and habitat fragmentation; pollution; disease; among others (Buhlmann et al. 2009, Rhodin et al. 2011). Efforts to manage and conserve tortoises therefore require baseline information on the status of tortoise species across their geographic distribution (Swann et al. 2002, Smith et al. 2009, Stober & Smith 2010). Ranging behavior can provide information about habitat use: activity, energy balance, resource availability, opportunities for reproduction and social interaction (Nagy & Medica 1986, Moskovits & Kiester 1987, O'Connor et al. 1994, Averill-Murray et al. 2002).
The study was part of a larger project to promote indigenous community management of indigenous lands and protected areas in the Bolivian Chaco. The red-footed tortoise Geochelone carbonaria Spix 1824 (Testudines: Testudinidae) is reported as a game species for subsistence and commercial hunters in Venezuela (Aponte et al. 2003) and Brazil (Peres 2000, Pezzuti et al. 2010). In the Bolivian Chaco, it is an important subsistence resource for the Ayoreo indigenous people (Ayala 1997, Etacore et al. 2000), and its fat is used in traditional medicine by the Isoseño-Guaraní indigenous people (Cuéllar 2000 a, b, Soria & Noss 2000). Preliminary observations and interviews in the Kaa-Iya landscape suggested the species to be much more abundant in protected areas free of hunting and livestock pressure, despite assertions by local Isoseño-Guaraní residents that they did not consume its meat. The species is considered to be near threatened in Bolivia (Cortéz & Rey-Ortíz 2009). It is also important as a prey item for jaguars Panthera oncaa flagship species for the Kaa-Iya landscapein this region (L. Maffei, 2002, pers. com.) and as a seed disperser (Moskovits & Bjorndal 1990, Strong & Fragoso 2006, Wang et al. 2011).
The few field studies on this species have focused on reproduction, diet, seed dispersal, ranging behavior and abundance. Most research has concentrated in humid Amazonian forests (Medemet al. 1979, Moskovits 1985, Moskovits & Kiester 1987, Moskovits 1988, Bjorndal 1989, Moskovits & Bjorndal 1990, Strong & Fragoso 2006), with only one study in seasonally f looded Pantanal savanna (Wang et al. 2011) and one in dry forest (Aponte et al. 2003). The Gran Chaco dry forest represents the southern distribution limit for Geochelone carbonaria, where it faces the most extreme climatic conditions in terms of winter cold and seasonal drought. We therefore evaluated ranging patterns at the protected sites in order to promote conservation measures for the species on indigenous and private ranch lands. This study is the first to report ranging behavior at two widely separated sites in the Gran Chaco ecoregion, based on radio-telemetry tracking. A complementary paper reports on activity patterns and burrow use (Noss et al. in press).
Study area
The Kaa-Iya del Gran Chaco National Park covers 34,400 km2 in eastern Santa Cruz Department (Fig. 1). This is the most diverse and best-conserved portion of the Gran Chaco ecoregion (Taber et al. 1996), with over 50 reptile species recorded including four chelonians: Geochelone carbonaria, G. chilensis, Kinosternon scorpioides and Acanthochelys macrochephala (Gonzáles 1998, 2001). Of these four, G. carbonaria is by far the most abundant and widely distributed across the Kaa-Iya National Park (Montaño 2003, Gonzáles & Montaño 2007, Montaño & Gonzáles 2007, 2008).

We established the Cerro Cortado field camp (19°31'35" S, 62°18'34" W) in 1997 on the boundary between the Kaa-Iya del Gran Chaco National Park and the neighboring Isoso TCO (indigenous territory). We established Ravelo field camp (19°17'43" S, 60°31'10" W) in 2000 in the southeastern portion of the Kaa-Iya National Park, near where Vanzolini (1999) collected his specimens, 15 km from the Paraguay border, and over 170 km from the Cerro Cortado camp. The Chaco alluvial plain lies at an altitude of approximately 300 m, with isolated hills (Cerro Cortado, Cerro Colorado, Cerro San Miguel) at the study sites that rise 100-200 m above the surrounding plain. Average annual rainfall is 550 mm at Cerro Cortado and 700 mm at Ravelo, with a dry season from May to November.
The vegetation at Cerro Cortado is a xeric Chaco forest, with low forests and thorn scrub distributed according to soil types (sand versus clay). The canopy is 3-8 m high with a predominance of Acacia praecox (Leguminosae-Mimosoidae), Ruprechtia triflora (Polygonaceae) and Castela coccinea (Simaroubaceae). Emergents up to 20 m high are Cereus validus, Stetsonia coryne (Cactaceae), Aspidosperma quebracho-blanco, A. pirifolium, Caesalpinia paraguariensis (Leguminosae-Caesalpiniaceae), and Ziziphus mistol (Rhamnaceae). The vegetation at Ravelo is a transitional Chaco-Cerrado dry forest, with a canopy 10-15 m high, rich in vines but poor en epiphytes. The most abundant tree species include Astronium urundeuva (Anacardiaceae), Cordiaglabrata (Boraginaceae), Aspidosperma cf. tomentosum, Caesalpinia pluviosa (Leguminosae-Caesalpiniaceae), Anadenanthera macrocarpa (Leguminosae-Mimosoideae), and Amburana cearensis (Leguminosae-Papilionoideae). Ground cover at both sites includes numerous species of cactus and bromeliads (Navarro 2004).
Methods
Radio-telemetry has been successfully used in several tortoise and turtle species to describe ranging patterns: Kinixys belliana in Tanzania (Bertram 1979), Gopheruspolyphemus in Georgia, USA (Eubanks et al. 2003), G. assizii in the southwestern USA(Swann et al. 2002), Phrynops rufipes in Brazil(Magnusson et al. 1997), Testudo hermanni in Italy (Mazzotti et al. 2002), as well as Geochelone carbonaria and G.denticulata in Brazil (Moskovits & Kiester 1987). In many tortoise studies, telemetry transmitters are attached using epoxy only (Boarman et al. 2008). We decided to combine this method with bolts to in order to avoid detachment by tortoises pushing their way through the dense and thorny Chaco vegetation at our study sties, during tracking periods extending over a year. We anesthetized the tortoises in order to minimize pain from drilling the holes through the scutes, which occasionally induces minor bleeding, and to surgically implant internal thermometers for a parallel study on activity patterns and burrow use. These protocols were approved by the Texas A & M University Institutional Animal Care and Use Committee / Animal Welfare Assurance Program (2000-230).
We anesthetized the animals using an intramuscular (I.M.) injection of either Telazol® (Tiletamine-Zolazepam, 20 mg/kg) or Ketamine (7.5 mg/kg) with Medetomidine (75 mcg/kg), and Isoflurane® administered using a cotton ball coated with 0.25 ml of liquid Isoflurane® and placed within a syringe case and positioned near the nares as previously described by Deem and Fiorello (2002). When the tortoises were fully anesthetized, veterinarians used glue and bolts to attach a Holohil® or ATS® radio transmitter to the dorsal aspect of the carapace. Tortoises were kept in captivity until full recovery from anesthesia, which ranged between 24-72 hours, and released at their capture location.
We then obtained daily locations for the tortoises by tracking the radio signals with a Telonics TM receiver and a Yagi two-element directional antenna. After locating the animal visually, we took the coordinates of the precise location with a GPS receiver, and recorded the date, time and the tortoise's activity. We analyzed all GPS locations for each animal using ArcView 3.5, and estimated seasonal (dry versus wet season) and overall home ranges using minimum convex polygons.
Results
We captured and fixed radio-transmitters on nine individuals (five males and four females) at Cerro Cortado, tracking the animals from January of 2000 to April of 2002 (Table 1). Six animals were tracked continuously for over a year, and four of these for a total of 21 months each. The number of locations per individual ranged from 52-218. In Ravelo we captured six individuals (three males and three females), and followed them from May 2001 to January 2002. However, political difficulties between park authorities and military personnel using the site forced us to abandon work there, and we recorded only 3-12 locations per individual. One animal disappeared (possible radio failure) and another was killed by a predator within the first three months.

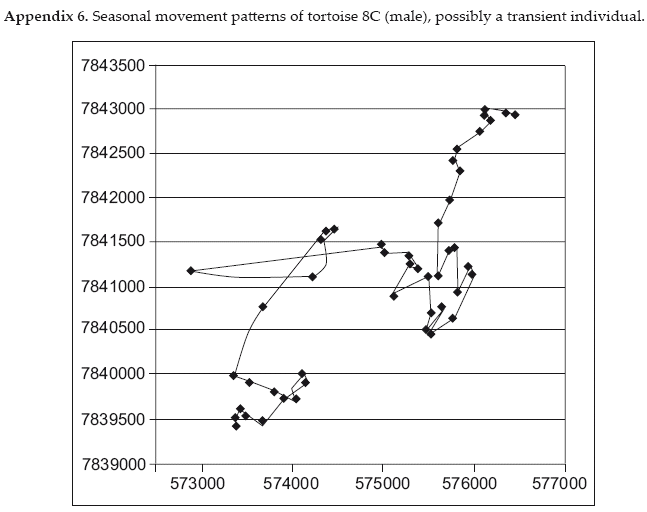
Tortoises at Cerro Cortado occupy overall home ranges from 50-600 ha (Table 1). With only two months of tracking, during the coldest months of the year, the 2 ha observed range for male C7 appears to be an under-estimation. The other four males have home ranges on average three times larger in comparison to four female tortoises 308 versus 103 ha though these differences are not statistically significant (Student's one-tailed t test, p>0.1). With the exception of male C1, who eventually disappeared beyond the study area, and C8 who was only tracked for 6 months but described the largest range of any individual tracked, the tortoises appear to maintain home ranges. The maximum observed distance covered by tortoises, again excluding C7, ranged from 1.3-4.6 km (mean 2.5 km, N=8), with males moving further than females (3.25 vs. 1.825 km, Student's one-tailed t-test, p=0.086).
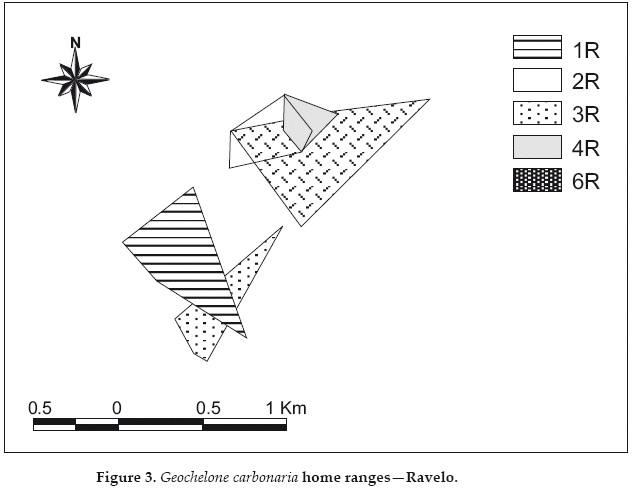
At Ravelo with very few locations, and fewer months of tracking per individual, observed ranges are less extensive: 5.8-38.0 ha (N=5). Maximum observed distances were also less than at Cerro Cortado, from 0.3-1.2 km (mean 0.75 km). The female R3 disappeared in October of 2001, but was encountered again by accident in January of 2003, within the same area where she had first been captured.
We found differences in home range from dry to wet season for each of the six tortoises that were radio-tracked for at least 12 months (Table 2, Student's one-tailed t-test, p=0.068).
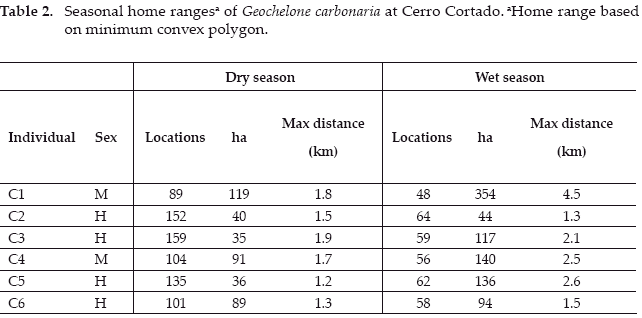
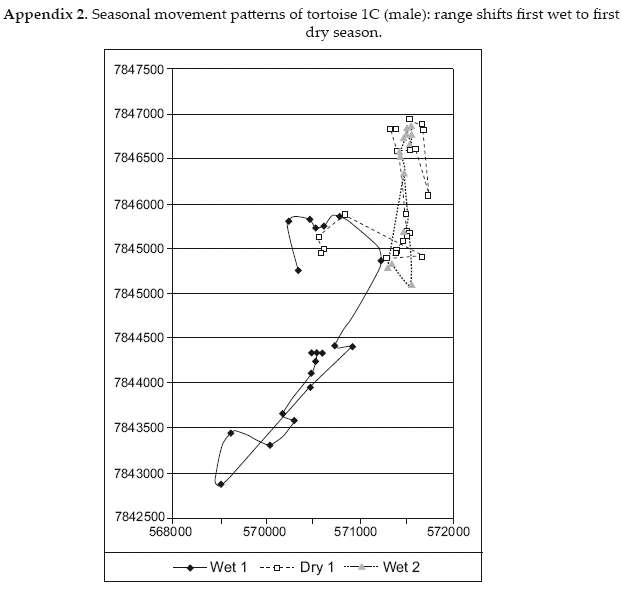
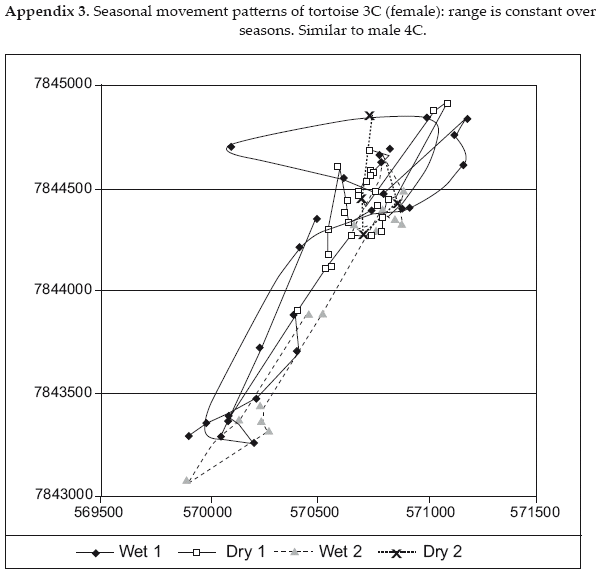
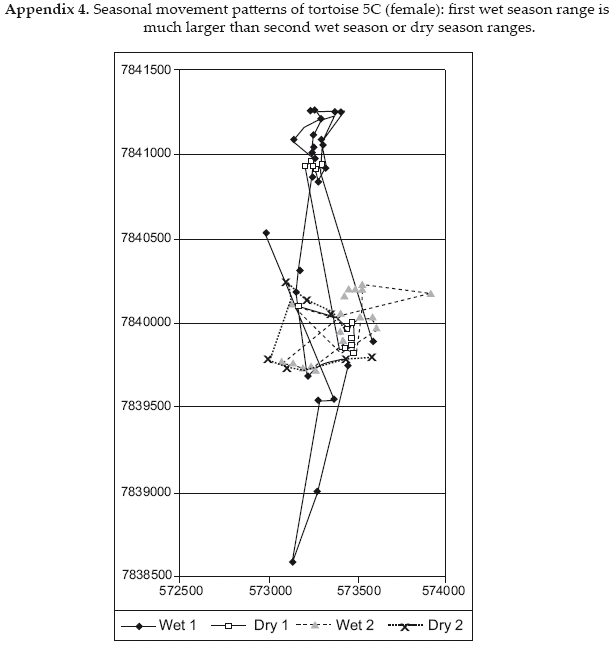
Despite registering more locations during the dry season, individual dry season home ranges were only 26-94 percent of wet season ranges for the same individuals. These data coincide with the evidence described above that the animals continue to be active during the dry season, but at a reduced level. It is notable that only in one case, female C5, the dry season range was a reduced portion of the wet season range. All other tortoises shifted to some degree the ranges they used from one season to the other, though individual variation in ranging patterns was great (Appendices A1-A6).
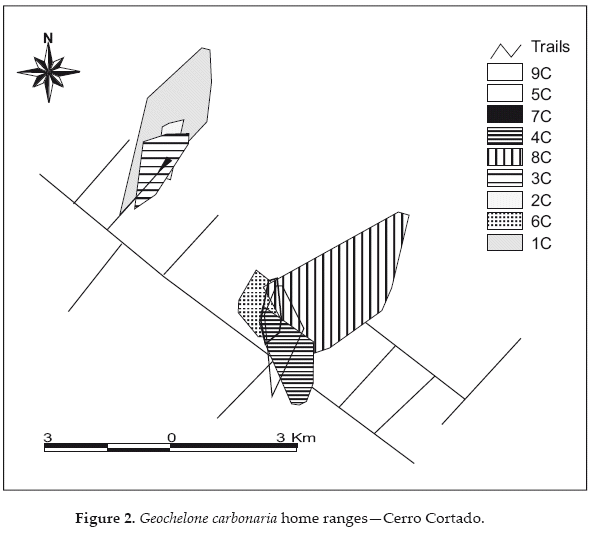
At both sites we observed range overlap among individuals of both sexes (Figs. 2-3). For example, at Cerro Cortado we observed overlaps that represented 40-70% of the respective home ranges of females C2 and C2, 23-29% between females C5 and C6, and 5-61% between males C4/C8/C9. The male C9 shared from 80-96% of his range with females C5 and C6 respectively, while females C2 and C3 shared 83-89% of their ranges with male C1, and female C5 shared 78-90% of her range with males C4 and C8 respectively. In total, we observed range overlaps between four and five neighboring animals at the two portions of the Cerro Cortado study site. We also encountered additional tortoises of both sexes within the ranges of individuals wearing radio-transmitters.
On 592 occasions, tortoises were located on successive days. On 401 of these occasions the tortoises had not moved at all but remained in the same burrow or shelter. The longest single-day displacement for a tortoise, 1,360 m, was registered for female 6C. Male 4C also registered a single-day displacement of over 1,000 m. The longest single-day displacement for a tortoise at Ravelo was 630 m for male 6R. Including all the days without movement, average daily displacement was higher in the wet season (71 m, SD 160) than in the dry season (19 m, SD 94) (Student's one-tailed t-test, p<0.01), and this pattern was followed by both males and females. However, when tortoises did move, single-day displacement was higher in the dry season (mean 192 m, SD 238) than in the wet season (mean 167 m, SD 207), though the differences are not statistically significant, and for females 2C and 3C the opposite was true. Males (N=7 individuals, mean 210 m, SD 219) tended to move further than females (N=4 individuals, mean 183 m, SD 250) in the dry season, but females (mean 178 m, SD 242) moved further than males (mean 150 m, SD 139) in the wet season, though neither of these differences is significant.
Discussion
Home ranges are considered to be areas that animals visit during their daily activities, in which they obtain the resourcesfood, water, mineralsas well as reproductive and social interactions that they require (O'Connor et al. 1994). Male red-footed tortoises in the Chaco utilize larger ranges than do females, presumably as males wander in search of females. This is consistent with reports for the same species at an Amazonian site (Moskovits & Kiester 1987), as well as for the desert tortoise Gopherus assizii, with males moving further in the summer which is the mating season (O'Connor et al. 1994, Averill-Murray et al. 2002).
As at a more benign Amazonian site (Moskovits & Kiester 1987), individual red-footed tortoises at Cerro Cortado differ significantly in ranging patterns (Appendices A1-A6). However, the Chaco's stronger seasonality (temperatures from <0 - +40°C, 6+ months without surface water) may produce more distinct seasonal requirements and induce range shifts as observed in some individuals. Minimum convex polygon home ranges are larger at the Cerro Cortado Chaco site (mean 183 ha, range 2.8-628.9 ha) than at the Ravelo Chaco site (mean 19 ha, range 3.8-38 ha) or the Amazon moist forest of Brazil (mean 26 ha, range 0.63-117.5 ha). This may result in part from the longer period of study in the Cerro Cortado case, 14-21 months for six of the individuals in contrast to six months in the Ravelo case and a maximum of five months in the Brazil case (Moskovits & Kiester 1987). The larger home range may also result from the drier conditions of Cerro Cortado. This is consistent with findings for Testudo hermanni at the northern edge of its range (Mazzotti et al. 2002). We did not find that female tortoises in the Chaco use relatively circular home ranges while males use relatively rectangular, as reported by Moskovits & Kiester (1987). Also contrary to the Brazilian site, we found significant seasonal shifts in home ranges for some individuals.
Conclusion
The tortoises occupy home ranges of 50-600 ha, with males using ranges three times larger than females. Individual home ranges overlap, across and between sexes. Tortoises shift their ranges shift ranges from the dry season to the wet season, and use larger ranges in the wet season. These patterns of overlapping ranges, larger male ranges, and larger ranges in the reproductive season are consistent with other tortoises. However, the relatively large ranges for red-footed tortoises in the Chaco, relative to their ranges in Amazonian habitats, may result from the strong seasonal climate in the Chaco.
Observed ranges are relatively large for an animal of this size in the Chaco, and ranges shift across seasons, in Kaa-Iya's protected areas. Ranges are likely to be even larger within human-impacted landscapes as tortoises seek scarce resources and mates. Indigenous territories and ranches in dry forests where this species occurs will need to provide key resources that include appropriate food, water, and shelter where these become scarce, or alternatively set aside communal and private reserves exceeding 1,000 ha in order to maintain tortoise populations on their lands. Detailed research on tortoise abundance, ranging patterns, resource availability and threats would be required to determine the area necessary to maintain a viable tortoise population in each indigenous territory or private land. Ideally larger sample sizes would support stronger management decisions, if costs and logistics permit.
Acknowledgments
This publication was made possible in part by financial support from the United States Agency for International Development (USAID, Cooperative Agreement No. 511-A-00-01-00005). The opinions expressed here are those of the authors and do not necessarily represent those of USAID. The Wildlife Conservation Society (WCS) and the Capitanía del Alto y Bajo Isoso (CABI) provided further support. CABI's parabiologists and wildlife monitors tracked the tortoises with radios, while also recording data on and marking other tortoises encountered. Lucindo Gonzáles supported the field work and reviewed early versions of the manuscript. Rosa Leny Cuéllar produced the Arcview maps.
References
Aponte, C. G., R. Barreto & J. Terborgh. 2003. Consequences of habitat fragmentation on age structure and life history in a tortoise population. Biotropica 35: 550-554.
Averill-Murray, R. C., B. E. Martin, S. J. Bailey & E. B. Wirt. 2002. Activity and behavior of the Sonoran desert tortoise in Arizona. pp. 135-158. In: van Devender, T.R. (ed.) The Sonoran Desert Tortoise: Natural History, Biology, and Conservation. University of Arizona Press, Tucson, Arizona. [ Links ]
Ayala C., J. M. 1997. Utilización de la fauna silvestre del grupo étnico Ayoréode en la comunidad Tobité, Santa Cruz, Bolivia. Under graduate thesis in biology, Universidad Autónoma Gabriel René Moreno, Santa Cruz. 78 p. [ Links ]
Bertram, B. C. R. 1979. Home range of a hingeback tortoise in the Serengeti. African Journal of Ecology 17: 241-244.
Bjorndal, K. A. 1989. Flexibility of digestive responses in two generalist herbivores, the tortoises Geochelone carbonaria and Geochelone denticulata. Oecologia 78: 317-321. [ Links ]
Boarman, W. I., T. Goodlett, G. Goodlett & P. Hamilton. 1998. Review of radio transmitter attachment techniques for turtle research and recommendations for improvement. Herpetological Review 29: 26-33.
Buhlmann, K. A., T. S. B. Akre, J. B. Iverson, D. Karapatakis, R. A. Mittermeier, A. Georges, A. G. J. Rhodin, P. P. van Dijk & J. W. Gibbons. 2009. A global analysis of tortoise and freshwater turtle distributions with identification of priority conservation areas. Chelonian Conservation and Biology 8:116-149.
Cortéz F., C & G. Rey-Ortíz. 2009. Chelonoidis carbonaria (Spix, 1824): Testudines-Testudinidae. pp. 611-612. In: Aguirre, L. F., R. Aguayo, J. Balderrama, C. Cortéz & T. Tarifa (eds.) Libro Rojo de la Fauna Silvestre de Vertebrados de Bolivia. Ministerio de Ambiente y Agua, La Paz. [ Links ]
Cuéllar, R. L. 2000a. ¿Para qué sirven los animales del monte en Izozog? Capitanía de Alto y Bajo Isoso, Wildlife Conservation Society, Santa Cruz. 29 p. [ Links ]
Cuéllar, R. L. 2000b. Uso de los animales silvestres por pobladores Izoceños. pp. 471-484. In: Cabrera, E., C. Mercolli & R. Resquin (eds.) Manejo de Fauna Silvestre en Amazonía y Latinoamérica. CITES Paraguay, Fundación Moises Bertoni, University of Florida, Asunción. [ Links ]
Deem, S. L. & C. V. Fiorello. 2002. Capture and immobilization of free-ranging edentates. In: Heard, D. (ed.) Zoological Restraint and Anesthesia. Document B0135.1202. International Veterinary Information Service (www.ivis.org), Ithaca, New York. http://www.ivis.org/special_books/Heard/deem/chapter_frm.asp?LA=1 [ Links ]
Etacore, J., A. Higazi, J. Beneria-Surkin & W. R. Townsend. 2000. Yoca iaá utatai: conocimiento Ayoreo de la comunidad el Porvenir: la peta negra. Publicaciones Proyecto de Investigación No. 9. Confederación de Pueblos Indígenas de Bolivia, Santa Cruz. 30 p.
Eubanks, J. O., W. K. Michener & C. Guyer, C. 2003. Patterns of movement and burrow use in a population of gopher tortoises (Gopherus polyphemus). Herpetologica 59: 311-321.
Gonzáles A., L. 1998. La herpetofauna del Izozog. Ecología en Bolivia 31: 45-52.
Gonzáles A., L. 2001. Los anfíbios y reptiles en una zona del Chaco boreal de Santa Cruz, Bolivia: riqueza, composición y biogeografía. Under graduate thesis in biology, Universidad Autónoma Gabriel René Moreno, Santa Cruz. 41 p. [ Links ]
Gonzáles, L. & R. Montaño. 2007. La herpetofauna del Parque Nacional Kaa Iya del Gran Chaco y la Tierra Comunitaria de Origen Isoso, Santa Cruz-Bolivia. Technical Report #193. Wildlife Conservation Society, Museo de Historia Natural Noel Kempff Mercado, Santa Cruz. 8 p. [ Links ]
Magnusson, W. E., A. Cardoso de Lima, V. Lopes da Costa. & R.C. Vogt. 1997. Home range of the turtle, Phrynops rufipes, in an isolated reserve in central Amazonia, Brazil. Chelonian Conservation and Biology 2: 494-499.
Mazzotti, S., A. Pisapia & M. Fasola. 2002. Activity and home range of Testudo hermanni in Northern Italy. Amphibia-Reptilia 23: 305-312.
Medem, F.O., V. Castaño & M. Lugo-R. 1979. Contribución al conocimiento sobre la reproducción y el crecimiento de los "morrocoyes" (Geochelone carbonaria y G. denticulata; Testudines, Testudinidae). Caldasia 12: 497-511.
Montaño F., R. R. 2003. Caracterización de la herpetofauna de la Salina Ravelo, Departamento Santa Cruz, Bolivia. Technical Report #90. Wildlife Conservation Society, Santa Cruz. 25 p. [ Links ]
Montaño, R. & L. Gonzáles. 2007. Diversidad de anfibios y reptiles en el humedal Palmar de las Islas. Technical Report #189. Wildlife Conservation Society, Santa Cruz. 13 p. [ Links ]
Montaño, R. & L. Gonzáles. 2008. El Humedal Palmar de las Islas: una guía ilustrativa. Wildlife Conservation Society, Santa Cruz. 34 p. [ Links ]
Moskovits, D. K. 1985. The behavior and ecology of the two Amazonian tortoises, Geochelone carbonaria and Geochelone denticulata, in northwestern Brazil. PhD Dissertation in Biology. University of Chicago, Chicago. 328 p. [ Links ]
Moskovits, D. K. 1988. Sexual dimorphism and population estimates of the two Amazonian tortoises (Geochelone carbonaria and G. denticulata) in northwestern Brazil. Herpetologica 44: 209-217. [ Links ]
Moskovits, D. K. & A. R. Kiester. 1987. Activity levels and ranging behaviour of the two Amazonian tortoises, Geochelone carbonaria and Geochelone denticulata, in north-western Brazil. Functional Ecology 1: 203-214.
Moskovits, D. K. & K. A. Bjorndal. 1990. Diet and food preferences of the tortoises Geochelone carbonaria and G. denticulata in northwestern Brazil. Herpetologica 46: 207-218.
Nagy, K. A. & P. A. Medica. 1986. Physiological ecology of desert tortoises in southern Nevada. Herpetologica 42: 73-92.
Navarro, G. 2004. Mapa de Vegetación del Parque Nacional y ANMI Kaa-Iya del Gran Chaco. Wildlife Conservation Society, Santa Cruz. 42 p. [ Links ]
Noss, A. J., R. R. Montaño F., F. Soria, S. L. Deem., C. V. Fiorello & L. A. Fitzgerald. In press. Geochelone carbonaria (Testudines: Testudinidae) activity patterns and burrow use in the Bolivian Chaco. South American Journal of Herpetology.
O'Connor, M. P., L. C. Zimmerman, D. E. Ruby, S. J. Bulova & J. R. Spotila. 1994. Home range size and movements by desert tortoises, Gopherus agassizii, in the eastern Mojave desert. Herpetological Monographs 8: 60-71.
Peres, C. A. 2000. Evaluating the impact and sustainability of subsistence hunting at multiple Amazonian forest sites. pp. 31-56. In: Robinson, J.G. & E.L. Bennett (eds.). Hunting for Sustainability in Tropical Forests. Columbia University Press, New York. [ Links ]
Pezzuti, J. C. B., J. P. Lima, D. F. da Silva & A. Begossi. 2010. Uses and taboos of turtles and tortoises along Rio Negro, Amazon Basin. Journal of Ethnobiology 30:153-168.
Rhodin, A. G. J., A. D. Walde, B. D. Horne, P. P. van Dijk, T. Blanck & R. Hudson. 2011. Editorial introduction and executive summary. pp.3-16. In: Turtle Conservation Coalition. Turtles in trouble: The world's 25+ most endangered tortoises and freshwater turtles. IUCN/SSC Tortoise and Freshwater Turtle Specialist Group, Lunenburg, Massachusetts. [ Links ]
Smith, L. L., J. M. Linehan, J.M. Stober, M.J. Elliott & J. B. Jensen. 2009. An evaluation of distance sampling for large-scale gopher tortoise surveys in Georgia, USA. Applied Herpetology 6: 355-368.
Soria M., F. & A. Noss, A. 2000. Herpetofauna de Cerro Cortado con referencias específicas a Tupinambis spp. y Chelonoidis spp. pp. 361-365. In: Cabrera, E., C. Mercolli & R. Resquin (eds.) Manejo de Fauna Silvestre en Amazonía y Latinoamérica. CITES Paraguay, Fundación Moises Bertoni, University of Florida, Asunción. [ Links ]
Stober, J. M. & L. L. Smith. 2010. Total counts versus line transects for estimating abundance of small gopher tortoise populations. Journal of Wildlife Management 74:1595-1600.
Strong, J. N. & J. M. V. Fragoso. 2006. Seed dispersal by Geochelone carbonaria and Geochelone denticulata in northwestern Brazil. Biotropica 38: 683-686.
Swann, D. E., R. C. Averill-Murray & C. R. Schwalbe. 2002. Distance sampling for Sonoran desert tortoises. Journal of Wildlife Management 66: 969-975.
Taber, A., G. Navarro & M. A. Arribas. 1997. A new park in the Bolivian Gran Chacoan advance in tropical dry forest conservation and community-based management. Oryx 31:189-198.
Vanzolini, P. E. 1999. A note on the reproduction of Geochelone carbonaria and G. denticulata (Testudines, Testudinidae). Revista Brasileña de Biología 59: 593-608.
Wang, E., C. I. Donatti, V. L. Ferreira, J. Raizer & J. Himmelstein. 2011. Food habits and notes on the biology of Chelonoidis carbonaria (Spix 1824) (Testudinidae, Chelonia) in the southern Pantanal, Brazil. South American Journal of Herpetology 6:11-19.


Artículo recibido en: Octubre de 2012.
Manejado por: Ignacio de la Riva.
Aceptado en: Enero de 2013.

