









 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO  uBio
uBio 
 Permalink
PermalinkEcología en Bolivia
versão impressa ISSN 1605-2528versão On-line ISSN 2075-5023
Ecología en Bolivia v.47 n.2 La Paz set. 2012
Nota
Estrategias de forrajeo de cuatro especies de picaflores (Aves, Trochilidae) en la ceja de monte yungueña (La Paz, Bolivia)
Foraging strategies of four hummingbird species (Aves, Trochilidae) in "ceja de monte yungueña" (La Paz, Bolivia)
Vanesa Serrudo-Gonzáles1, Daniela Arteaga-Voigt2, Susy Fuentes-Bazán2, Emilia García-Estigarribia1 & Ruddy Luna-Barrón2
1Herbario Nacional de Bolivia, Calle 27 de Cota Cota, Casilla 10077 - Correo Central, La Paz Bolivia,
Email: vanesaserrudo@yahoo.es, Autora de correspondencia
2Centro de Investigación Genética de la Policía Boliviana, Av. Hugo Ernest Nº7404, La Paz, Bolivia
La ceja de monte yungueña está distribuida en Bolivia entre 2.900-4.200 m (Navarro 2002). Se caracteriza por presentar nieblas casi constantes y niveles de precipitación de un bioclima húmedo a hiperhúmedo. Contiene la mayor proporción de especies de aves de rango restringido y el mayor porcentaje de especies ornitológicas insuficientemente protegidas (Herzog et al. 2005).
En este hábitat las cuatro familias más ricas en especies de aves son: Trochilidae, Tyrannidae, Thraupidae y Parulidae. La primera familia ha desarrollado al menos dos estrategias de forrajeo o búsqueda de alimento: el ruterismo y territorialismo (Feinsinger & Colwell 1978). La estrategia de forrajeo rutera consiste en la búsqueda de alimento a través de una ruta más o menos regular donde las flores se encuentran muy dispersas y producen recompensas de néctar acorde al gasto energético (Feinsinger & Colwell 1978, Gill 1988, Garrison & Gass 1999). El territorialismo se define como la defensa de un área por un animal para ganar más acceso exclusivo al recurso, por cuanto la estrategia de forrajeo territorialista consiste en la vigilancia y defensa de territorios en parches de alta densidad de flores (Feinsinger & Colwell 1978, Camfield 2006).
La persecución, según Camfield (2006), representa la respuesta más común de los dueños de los territorios contra los intrusos. Boyden (1978) mostró que los picaflores emplean con más frecuencia este comportamiento cuanto mayor es el grado de amenaza de robo de alimento desde sus parches. El objetivo del presente trabajo es evaluar las estrategias de forrajeo de cuatro especies de picaflores (Trochilidae): Heliangelus amethysticollis, Metallura tyrianthina, Pterophanes cyanopterus y Coeligena violifer en la ceja de monte yungueña, utilizando el número de persecuciones y desarrollando una nueva metodología basada en índices.
El estudio se llevó a cabo desde enero a mayo de 2010 con un esfuerzo de muestreo de 33 horas. Durante estos meses se apreciaron mayores picos de floración y estas especies no defienden territorios destinados a la reproducción. Se trabajó a lo largo de los cuatro primeros kilómetros de la senda Sillutinkara dentro el Parque Nacional y Área Natural de Manejo Integrado (PN-ANMI) Cotapata (SERNAP 2001) desde 2.980-3.460 m de altitud. Se establecieron siete parches florales utilizados por las especies de picaflores, distanciados entre sí por más de 150 m. Dos de estos parches (primero y segundo) se encontraban en una misma matriz compuesta por sólo dos especies de plantas, mientras que los restantes cinco estaban compuestos por más de dos especies de plantas y no pertenecían a una misma matriz. Éstos fueron definidos como parches florales por encontrarse separados uno del otro a una distancia considerable y estar compuestos por especies vegetales que los picaflores visitaban con regularidad.
Cada estrategia de forrajeo (p.e. territorial, rutero) se evaluó mediante: 1) número promedio de persecuciones/h/parche; 2) tiempo promedio de estadía en el parche min/h y 3) presencia de la especie en el parche. Se define como persecución cuando el individuo bajo estudio vuela agresivamente detrás de otro (intra o interespecífico). El tiempo de estadía en el parche es el que el individuo se establece en unos puntos de percheo (este punto no necesariamente es el mismo, puede establecerse hasta en tres puntos distintos); la presencia en el parche se refiere al registro de la especie bajo estudio dentro de los límites del parche. El valor obtenido para cada especie de picaflor fue la sumatoria de cada variable arriba mencionada. Para definir dos comportamientos: territorialista y rutero, se trabajó con tres índices:
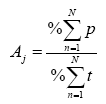
Donde, A = Índice de agresividad, que determina el grado de agresividad de la especie; j = especie; p = número de persecuciones totales/parche/hora/especie; t = tiempo total de estadía en el parche min/hora/especie; n = parche muestreado y N = número total de parches muestreados. Al obtener el número de persecuciones totales de cada especie, todas fueron sumadas para determinar el 100% de las agresiones en el sitio y determinar en porcentaje cuál especie fue la más agresiva.
En el caso del tiempo de estadía en el parche, se consideró como 100% el tiempo de estadía más largo para las cuatro especies, determinándose el porcentaje de las otras por regla de tres:
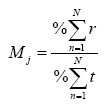
donde, M=índice de movilidad, que determina el grado de movilidad de la especie o grado de ruterismo; j = especie; r = número total de parches en los que la especie fue registrada; t = tiempo total de estadía en el parche (min)/hora/ especie; n = parche muestreado y N = número total de parches muestreados. Para la variable r, se consideró como 100 % la presencia de la especie en los siete parches estudiados:
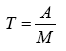
Donde, T=índice de territorialidad, A=índice de agresividad y M=índice de movilidad. El índice T identifica a una especie más territorialista que la otra, cuando su valor se encuentra más cercano a 1. Se utilizó una sumatoria de los datos obtenidos en cada parche porque no existe la seguridad de que cada individuo evaluado se mantenga en un solo parche floral, bajo el supuesto de que podría utilizar a lo largo del tiempo cualquiera de los parches muestreados.
Como resultados, al evaluar los comportamientos separadamente (Tabla 1), se evidenció la tendencia hacia el ruterismo por Metallura tyrianthina y Pterophanes cyanopterus, llegándose a la ausencia de persecuciones en ambas especies.
| Tabla 1. | Comportamientos característicos de individuos territorialistas de la familia Trochilidae |
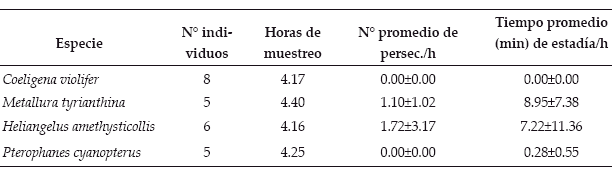 | |
El tiempo de vigilancia entre ambas especies también se asemeja, diferenciándose del de Heliangelus amethysticollis y Coeligena violifer, de entre las cuales la primera mostró una mayor tendencia a permanecer más tiempo en la percha y la segunda un comportamiento más agresivo. Los resultados con los índices empleados (Tabla 2) muestran la tendencia hacia el ruterismo por Metallura tyrianthina y Pterophanes cyanopterus y hacia el territorialismo por Heliangelus amethysticollis y Coeligena violifer.
| Tabla 2. | Índices de agresividad (A), movilidad (M) y territorialiadad (T) de Trochilidae. |
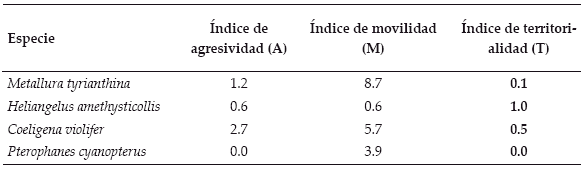 | |
De acuerdo con las características morfológicas estudiadas por Feinsinger & Colwell (1978), C. violifer (de pico y cuerpo largos) debió haber adoptado una estrategia de forrajeo rutera, lo cual no ocurrió. Una posible explicación podría ser la alta disponibilidad de alimento en el primer parche - donde dominaba la especie Desfontainia spinosa (Desfontainiaceae) con un alto número de flores (2.800 flores/día para dos individuos) -repitiéndose de este modo un patrón observado en Rastrorojo (Colombia a 2.200 m), sitio similar a la ceja de monte, donde Coeligena prunellei ocasionalmente estableció un comportamiento de defensa de recurso con Psamisia penduliflora (Ericaceae), especie dominante (Daza et al. 2006).
Una probable explicación del por qué C. violifer presentaría el mayor índice de agresividad se debería a que los sitios donde se hallaban sus territorios eran frecuentados por otras aves, ya que eran sitios de paso. Esto ocasionaría que los individuos se comporten más agresivamente al aumentar la densidad poblacional, factor no evaluado en el presente trabajo; sin embargo, se propone sea considerado en posteriores investigaciones.
Para Pterophanes cyanopterus - una especie con tendencia a la estrategia de forrajeo rutera - se corroboró la información obtenida por Gutiérrez & Rojas (2001), quienes estudiaron el bosque altoandino comprendido entre 3.100-3.500 m con una composición florística similar a la de la ceja. Sin embargo, ellos también mencionaron que en algunas épocas del año los individuos mostraron actividad territorial en flores de Siphocampylus (Campanulaceae) y Puya (Bromeliaceae), géneros presentes en la ceja, por lo cual es posible que se presente este comportamiento de junio a noviembre, meses que no fueron muestreados en este estudio.
Los resultados encontrados en el quinto parche, donde un individuo de H. amethysticollis mostró mayor tiempo de estadía que el otro, podrían deberse a diferencias de edad que determinan la experiencia, como el caso de la especie Calypte anna (Ewald & Rohwer 1980), donde los adultos enfrentan mayor número de encuentros agresivos que los juveniles. Otro factor podría ser la diferencia en el estatus de dominancia, que si bien se ha demostrado a nivel interespecífico con el trabajo de Ewald & Bransfield (1987), donde Archilocus alexandri está subordinado a Calypte anna, podría darse también a nivel intraespecífico.
Analizando los resultados desde una visión fitocentrista, se sugiere que las especies de plantas estarían favorecidas si los picaflores optan por estrategias de forrajeo rutera, porque evitarían una depresión de las poblaciones por endogamia, lo cual fue demostrado por Linhart et al. (1987) con el género Hansteinia (Acanthaceae), polinizado por Lampornis calolaema que es un picaflor territorialista, a diferencia de Razisea (Acanthaceae, que presentó panmixia), polinizado por Phaethornis guy, un picaflor rutero. A esto se suma el que las condiciones climáticas (frío) impulsarían a que las especies de picaflores sean los principales polinizadores al ser visitadores florales homeotermos; por tanto, evaluar el grado de heterocigocidad en estas plantas sería un siguiente paso.
Agradecimientos
Agradecemos a todos aquellos que han contribuido a la revisión del manuscrito, al Instituto de Ecología por el financiamiento, Herbario Nacional de Bolivia, Colección Boliviana de Fauna, NP-ANMI Cotapata, Arely Palabral Aguilera, James L. Luteyn, Sandra Rojas y al grupo Tantasarañani.
Referencias
Boyden, T. 1978. Territorial defense against hummingbirds and insects by tropical hummingbirds. The Condor 80:216-221. [ Links ]
Camfield, A. F. 2006. Resource value affects territorial defense by Broad-tailed and Rufous hummingbirds. Journal of Field Ornithology 77(2): 120-125. [ Links ]
Daza, A., S. Villamarin & J. E. Cely. 2006. Estado poblacional, recursos florales y hábitat de Coeligena prunellei (Trochilidae), ave endémica en peligro de extinción en la Reserva Biológica Cachalú, municipio de Encino (Santander). pp. 138-157 En: Solano N., C. V. (ed.) Memorias del I Seminario Internacional de Roble y Ecosistemas Asociados, Bogotá. [ Links ]
Ewald, P. W. & R. J. Bransfield. 1987. Territory quality and territorial behavior in two sympatric species of hummingbirds. Behavior Ecology Sociobiology 20: 285-293.
Ewald, P. E. & S. Rohwer. 1980. Age, coloration and dominance in nonbreeding hummingbirds. A test of asymmetry hypothesis. Behavior Ecology Sociobiology 7: 273-279.
Feinsinger, P. & R. Colwell. 1978. Community organization among Neotropical nectar-feeding birds. American Zoologist 18 (4): 779-795.
Garrison, J.E. & C. Gass. 1999. Response of a traplining hummingbird to changes in nectar availability. Behavioral Ecology 10(6): 714-725.
Gill, F. B. 1988. Trapline foraging by hermit hummingbirds: competition for an undefended, renewable resource. Ecology 69 (6): 1933-1942. [ Links ]
Gutiérrez, E. & S. V. Rojas. 2001. Dinámica anual de la interacción colibrí-flor en ecosistemas altoandinos del volcán Galeras, sur de Colombia. Tesis para optar a licenciatura en biología, Universidad Nacional de Colombia, Facultad de Ciencias, Departamento de Biología, Bogotá. 108 p. [ Links ]
Herzog, S., Soria, R. & B. Hennessey. 2005. Patrones ecoregionales de riqueza, endemismo y amenaza de la avifauna boliviana: prioridades para la planificación ecoregional. Ecología en Bolivia 40 (2): 27-40.
Linhart, Y., W. Busby, J. Beach & P. Feinsinger. 1987. Forager behavior, pollen dispersal and inbreeding in two species of hummingbirds-pollinated plants. Evolution 41 (3): 679-682.
Navarro, G. & M. Maldonado. 2002. Geografía ecológica de Bolivia: Vegetación y ambientes acuáticos. Centro de Ecología Simón I. Patiño, Santa Cruz. 719 p. [ Links ]
SERNAP (Servicio Nacional de Áreas Protegidas). 2001. Ministerio de Desarrollo Sostenible y Planificación. 2da edición. La Paz. 218 p. [ Links ]
Nota recibida en: Abril de 2012.
Manejada por: Ramiro Pablo López
Aceptada en: Agosto de 2012.