










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
 Citado por SciELO
Citado por SciELO Links relacionados
 Similares en
SciELO
Similares en
SciELO  uBio
uBio Compartir

 Permalink
PermalinkEcología en Bolivia
versión impresa ISSN 1605-2528versión On-line ISSN 2075-5023
Ecología en Bolivia v.47 n.2 La Paz sep. 2012
Artículo
Depredación post-dispersión de Guettarda viburnoides (Rubiaceae) en sabanas neotropicales del noreste de Bolivia: su efecto a pequeña escala espacial
Post-dispersal seed predation of Guettarda viburnoides (Rubiaceae) in Neotropical savannas, of NE Bolivia: its effect at a small spatial scale
Francisco Saavedra1, Andrea Loayza2 & Mónica Moraes R.1
1 Herbario Nacional de Bolivia, Instituto de Ecología, Universidad Mayor de San Andrés, Casilla 10077 - Correo Central, La Paz, Bolivia
Email: panncho@gmail.com Autor de correspondencia
2 Departamento de Biología, Facultad de Ciencias, Universidad de La Serena & Instituto de Ecología y Biodiversidad (IEB), Casilla 599, La Serena, Chile
Email: andrea.loayza@mac.com
Resumen
La depredación de semillas post-dispersión juega un papel crítico en la estructura y dinámica de poblaciones vegetales. Por lo tanto, identificar patrones de depredación de semillas y determinar los procesos que los generan son fundamentales para entender la organización de comunidades vegetales. En este estudio evaluamos la depredación post-dispersión de Guettarda viburnoides (Rubiaceae) en relación a la distancia al árbol madre y al hábitat. Hipotetizamos que la depredación de semillas será mayor 1) cerca del árbol madre que lejos de éste, 2) en parches de vegetación leñosa que árboles aislados o en áreas de pastizal. El patrón de depredación obtenido representa el patrón clásico de depredación en especies tropicales, donde la mayoría de las semillas es más depredada debajo del árbol madre que a distancias intermedias. Con relación al hábitat, la depredación de semillas fue mayor en parches de vegetación en comparación con árboles solitarios y sabanas. Asumimos que especies del género Pheidole son potencialmente los depredadores más importantes de G. viburnoides por su hábito alimenticio y presentar mayor abundancia. Este patrón de depredación de semillas apoya la hipótesis de Janzen y Connell y sugiere que a nivel de comunidades, en el sitio de estudio, los parches de vegetación y árboles solitarios en la sabana son hábitats con mayor depredación y actividad de hormigas.
Palabras clave: Depredación de semillas, Distancia-dependencia, Estación Biológica del Beni, Hábitat, Pheidole.
Abstract
Post-dispersal seed predation plays a critical role in the structure and dynamics of plant populations. Therefore, to identify patterns of seed predation and determine the processes that generate them are fundamental to understand the organization of plant communities. In this study we evaluated the post-dispersal seed predation of Guettarda viburnoides (Rubiaceae) in relation to distance from the parent tree and habitat. We hypothesize that seed predation will be greater 1) near the mother tree away from this, 2) woody vegetation patches of isolated trees, or in areas of grassland. The predation pattern obtained represents the classic pattern of predation in tropical species, where most seeds are preyed under the tree stem at intermediate distances. In relation to habitat seed predation was higher in patches of vegetation compared with solitary trees and savannas. We assume that species of the genus Pheidole are potentially the most important predators of G. viburnoides for their food habit and abundance. This pattern of seed predation supports the hypothesis of Janzen and Connell and suggests that at the community level in the study site, patches of vegetation and trees are solitary in the savanna habitats with higher predation and ant activity.
Keywords: Distant-dependent seed predation, Estación Biológica del Beni, Habitat, Pheidole.
Introducción
La depredación de semillas es uno de los procesos que causa mayor mortalidad en plantas vasculares y, por lo tanto, es un proceso clave para el reclutamiento de plantas (Janzen 1971, Wenny 2000, Hulme 2002, Hulme & Benkman 2002). Debido a que las semillas, al contrario de las plantas adultas, no pueden volver a crecer o alterar su composición química para incrementar su probabilidad de sobrevivencia, la mortalidad de semillas causada por la depredación post-dispersión es considerada como uno de los mayores limitantes para el reclutamiento de nuevos individuos (Hulme 2002, Hulme & Benkman 2002, Schupp 1990, Hulme 1998). Más aun, la depredación de semillas puede influenciar la sobrevivencia y la distribución espacial de las plantas (Orrock et al. 2006) y ejercer un fuerte efecto tanto sobre la estructura de comunidades vegetales, como sobre las dinámicas poblacionales de plantas (Janzen 1970, Schupp 1992, Maron & Simms 2001, Beckage & Clark 2005).
En áreas tropicales, se han reportado pérdidas de semillas superiores al 75% (Howe et al. 1985, Schupp 1988, Chapman 1989). Sin embargo, la presión causada por la depredación de semillas varía considerablemente entre diferentes hábitats (p.e. Jones et al. 2003, Loayza et al. 2011). Esta variación puede deberse a varios factores que incluyen: 1) variabilidad en la disponibilidad de semillas asociada a distintos hábitats (Jones et al. 2003, García et al. 2005), 2) diferencias en el grado en que distintos hábitats son adecuados para los depredadores (Janzen 1971), 3) variabilidad en los patrones de forrajeo de los depredadores (Avgar et al. 2008) y 4) variación espacial en las defensas de las semillas (Louda et al. 1987).
A pesar de las diferencias observadas entre hábitats, un patrón de mortalidad común en áreas tropicales es la mayor pérdida de semillas cerca de la planta madre o de plantas conespecíficas que lejos de ellas (Wenny 2000, Wyatt & Silman 2004, Norghauer et al. 2006). Como resultado, el área cercana a la planta madre (o a conespecíficas) presentará poco reclutamiento de plántulas (Janzen 1970, Connell 1971). Este patrón de reclutamiento, conocido como el patrón de Janzen y Connell es descrito por una curva donde hay pocos reclutas debajo o cerca de conespecíficos, y a medida que incrementa la distancia aumenta también el número de reclutas, hasta que a mayores distancias de conespecíficos el número de reclutas vuelve a caer debido abajas densidades de semillas.
A pesar de que el patrón de Janzen y Connell ha sido ampliamente estudiado en bosques tropicales (p.e. Harms et al. 2000, Wenny 2000), se conoce relativamente poco acerca de los mecanismos que determinan el patrón de reclutamiento de especies de plantas en sabanas tropicales. Estos ecosistemas se caracterizan por ser mosaicos de vegetación compuestos por distintos tipos de hábitats entre los que están los pastizales, fragmentos de bosque, parches de vegetación leñosa y árboles aislados (Ferreira et al. 2011). Consecuentemente, en sabanas la depredación post-dispersión de semillas va a estar influenciada tanto por la distancia a la planta madre, pero también por el hábitat. Por lo tanto, estos ecosistemas brindan una oportunidad para examinar de forma conjunta como interactúan estos dos factores.
En este estudio examinamos la depredación de semillas de Guettarda viburnoides (Rubiaceae) en sabanas inundables de la Estación Biológica del Beni en Bolivia. Específicamente, evaluamos cómo varía la depredación post-dispersión en relación a: 1) la distancia al árbol madre y, 2) el hábitat. Hipotetizamos que la depredación de semillas será mayor 1) cerca del árbol madre que lejos de este, 2) en parches de vegetación leñosa que árboles aislados o en áreas de pastizal.
Métodos
Área de estudio
Realizamos este estudio entre enero y julio de 2006 en las sabanas de la Reserva de la Biosfera Estación Biológica del Beni (RB-EBB), ubicada al sudoeste del departamento del Beni, en las provincias General Ballivián y Yacuma entre los municipios de San Borja y Santa Ana de Yacuma (14º30'- 14º50' Latitud Sur; 66º40'-65º50' Longitud Oeste; extensión aproximada de 2.400 ha; con altitud aproximada de 200 m) (Miranda 2000). El clima es pluviestacional húmedo; con una precipitación media anual de 1.900 mm y una temperatura promedio anual de 26ºC (máx. 38C, mín. 11ºC) (PL-480 &ANCB1998).
La vegetación dominante de las sabanas está conformada por asociaciones de pastos higromorfos, xeromorfos hasta pantanosos conformados por gramíneas, ciperáceas, varias familias de hábito acuático (ej. Araceae, Azollaceae, Marantaceae y Lentibulariaceae) y algunas especies arbustivas (ej. (Davilla nítida, Helicteres guazumifolia y Vernonia). Inmersos en las sabanas, se encuentran árboles aislados, parches de vegetación leñosa e islas de bosque. Los parches de vegetación leñosa constituyen áreas pequeñas (<200 m2, A.P. Loayza obs. pers.) con un dosel de hasta 10 m, donde se han establecido especies leñosas, que pertenecen principalmente a géneros de plantas dispersadas por animales como por ejemplo Guettarda (Rubiaceae), Virola (Myristicaceae), Piper (Piperaceae), Miconia (Melastomataceae), Solanum (Solanaceae), Pourouma (Urticaceae) y varias especies de palmeras (Loayza & Knight 2010, Loayza et al. 2011). Por otro lado, las islas de bosque son unidades aisladas de bosque (entre 1-20 ha), que presentan especies características de bosques tropicales y tienen un dosel de hasta 25 m (Comiskey et al. 2000).
Especie de estudio
Guettarda viburnoides Cham. y Schlecht. (Rubiaceae) es un árbol pequeño de hasta 6 m de altura que está distribuido desde Brasil hasta Paraguay en bosques semi-deciduos y sabanas (Taylor et al. 2004). En la RB-EBB G. viburnoides es una especie conspicua que crece predominantemente en parches de vegetación leñosa. Esta especie florece desde octubre hasta enero y tiene frutos maduros desde abril hasta julio, periodo en el cual se realizaron los experimentos de depredación. Los frutos son unas drupas sub-globosas con un endocarpo leñoso que mide entre 7-15 mm (Loayza et al. 2011). Cada endocarpo contiene 3-7 semillas, pero para la planta el endocarpo es la unidad de dispersión.
Depredación de semillas
Para evaluar la depredación de semillas realizamos inicialmente un experimento piloto con exclusiones metálicas, huelleros y bancos de semillas no protegidos para determinar si las mayores tasas de depredación se debían a vertebrados o a invertebrados. Encontramos poca remoción de endocarpos de G. viburnoides por vertebrados, pero altos niveles de depredación por invertebrados, principalmente hormigas (F. Saavedra datos no publicados). Por lo tanto, asumimos que los depredadores principales de esta especie eran principalmente hormigas. Para determinar si la depredación de semillas de G. viburnoides varía en relación a la distancia respecto al árbol focal seleccionamos 15 árboles adultos de la población ubicados cada uno en un parche de vegetación leñosa en la sabana. Los árboles seleccionados estaban separados entre sí por al menos 100 m. En cada árbol ofrecimos cuatro grupos de 10 endocarpos (depositados sobre mallas metálicas sobre el piso) ubicados a lo largo de un transecto de 40 m trazado en una dirección al azar a partir de la base de cada árbol. Los grupos de endocarpos se ubicaron de la siguiente manera: un grupo debajo del árbol, un grupo a los 10 m, un grupo a los 25 m y uno a los 40 m. Siguiendo la metodología de Wenny (2000), censamos los endocarpos día por medio durante la primera semana y semana por medio durante cuatro semanas. En cada censo reemplazamos con nuevos endocarpos aquellos que fueron removidos y registramos el número de endocarpos depredados. Asumimos que un endocarpo estaba muerto si era removido o si presentaba al menos una perforación; es decir si al menos una semilla dentro del endocarpo estaba deprededa. Este método da un valor conservador de la tasa de depredación pero ha sido exitosamente utilizado para comparar de manera no sesgada la depredación entre distintos hábitats (Loayza et al. 2011).
Para determinar cómo varía la depredación de semillas de G. viburnoides entre hábitats seleccionamos cuatro hábitats en el área de estudio: 1) parches de vegetación leñosa con un adulto de G. viburnoides 2) parches de vegetación leñosa sin individuos de G. viburnoides 3) árboles solitarios en la sabana y 4) hábitat de sabana. Cada uno de los hábitats fue replicado 15 veces (Ntotal=60) y en cada réplica establecimos un grupo de 10 endocarpos. Censamos los endocarpos día por medio durante la primera semana y semana por medio durante cuatro semanas. En cada censo se reemplazó con nuevos endocarpos aquellos que fueron removidos y se registró el número de endocarpos depredados.
Densidad de hormigas
Debido a que los resultados del ensayo piloto revelaron altos niveles de depredación por hormigas (F. Saavedra datos no publicados), evaluamos si la densidad de hormigas varía en función al árbol madre. Esto constituye un sucedáneo para determinar si los potenciales depredadores de G. viburnoides se distribuyen de manera no aleatoria en relación a la distancia al árbol madre. Para esto, seleccionamos los mismos 15 árboles de la población donde se instalaron los experimentos de depredación, y en cada árbol instalamos cuatro grupos de cinco trampas pit-fall desde la base del árbol hasta los 40 m. Las trampas fueron dispuestas de la siguiente manera: cinco trampas debajo del árbol, cinco trampas a los 10 m, 5-25 m y 5-40 m (Fig. 1). Adicionalmente, determinamos si la densidad de hormigas varía con relación al hábitat instalando un grupo de cinco trampas pit-fall en cada una de las 15 réplicas de los hábitats seleccionados (parches con G. viburnoides, parches sin G.viburnoides, árboles solitarios y sitios en sabana). Las trampas pit-fall consistían de vasos plásticos de 120 ml, que fueron llenados tres cuartas partes con una mezcla de detergente, agua y alcohol (Romero & Jaffe 1989).
Los especímenes colectados en las trampas se recogían cada 24 horas, durante 10 días.
En el laboratorio, las hormigas fueron clasificadas y separadas en morfoespecies según el color, forma y tamaño. Asumimos que las morfoespecies representadas por muy pocos individuos no eran depredadores numéricamente importantes (al ser poco abundantes). Posteriormente las morfoespecies más abundantes fueron identificadas hasta nivel de género utilizando la clave taxonómica de Fernández (2003). Los géneros determinados se clasificaron posteriormente en función al hábito alimenticio y para este estudio consideramos únicamente a las hormigas de hábito granívoro como potenciales depredadores.
Finalmente, para estimar la densidad de potenciales depredadores en cada distancia y tipo de hábitat calculamos una abundancia ajustada (AA) mediante la ecuación:
![]()
Donde, A = abundancia o el número total de individuos de cada especie y O = frecuencia de cada especie en el número total de trampas (Lindsey & Skinner 2001).
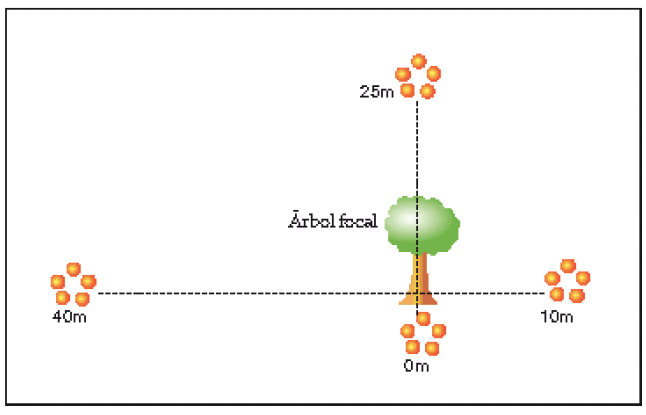 |
| Figura 1. Disposición de las trampas pit-fall instaladas en cada árbol. |
Análisis estadísticos
Para evaluar si la depredación de endocarpos varía con la distancia y con el tipo de hábitat utilizamos modelos lineales generalizados (MLGs: Cr awley 1993) en el que consideramos la distancia o el hábitat como factores principales, y el número de semillas depredadas al final del experimento (día 40) por distancia y/o habitat como variables de respuesta. En ambos análisis consideramos una distribución de errores de Poisson. Para examinar cómo varía la abundancia de hormigas con la distancia a los árboles de G. viburnoides y con el hábitat utilizamos ANOVAS. Para estos análisis transformamos la variable de respuesta (abundancia de hormigas) a LN10.
Resultados
Depredación de semillas
La tasa de depredación de semillas de G. viburnoides varió significativamente con la distancia al árbol de G. viburnoides (x2Wald= 17.45, gl=3, P=0.001). Específicamente, encontramos mayores niveles de depredación debajo del árbol (0 m) (1.47±0.40 endocarpos,) que a los 10 m (0.76±0.30 endocarpos) (P=0.008) y a los 40 m de éste (0.68±0.28 endocarpos) (P=0.002), no existiendo diferencias a los 25 m del árbol madre (0.45±0.17 endocarpos)(P=0.08). Estos resultados sugieren que la depredación de semillas de G. viburnoides es distancia-dependiente.
La depredación de semillas varió entre hábitats (x2Wald=21.77, gl=3, P=0.001), siendo mayor en parches de vegetación con árboles de G. viburnoides (8.86±0.21 endocarpos) que en parches de vegetación sin árboles de G. viburnoides (3.4±0.12 endocarpos) (P=0.002), que en árboles solitarios (3.33±0.13 endocarpos) (P=0.002) y que en áreas de sabana (4.13±0.13 endocarpos) (P=0.01).
Densidad de hormigas
Capturamos un total de 3.011 individuos, pertenecientes a 14 géneros de hormigas (Tabla 1). Entre éstos, el género granívoro Pheidole fue el que presentó mayor abundancia en las trampas pit-fall, por lo que asumimos que esta especie es potencialmente un depredador importante de G. viburnoides.
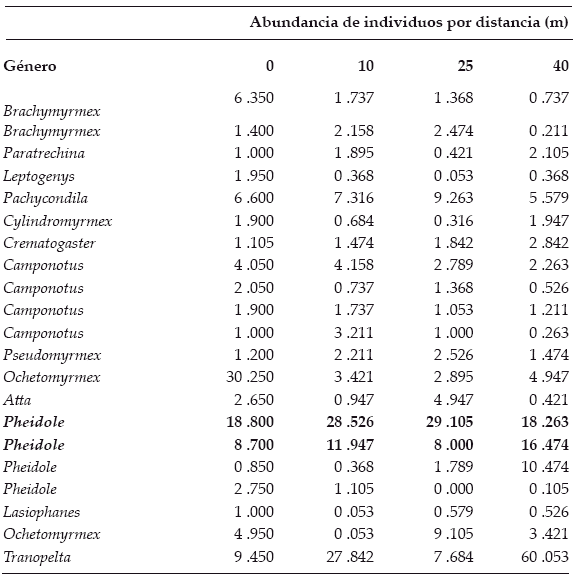 | |
| Tabla 1. | Abundancia promedio de hormigas por distancia y género (promedio ± E.E). Los valores más altos son resaltados en negrita. |
La densidad de individuos de las dos morfoespecies de Pheidole más comunes en las trampas no varió con relación a la distancia, Pheidole sp.1 (ANOVA F(3;2,29) P=0.11) y Pheidole sp.2 (ANOVA F(3;0,34) P=0.79) (Fig. 2).
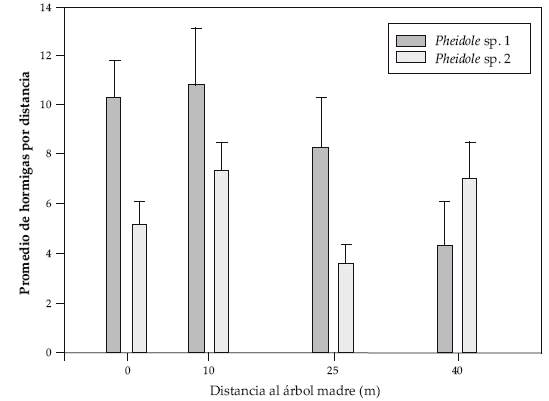 | |
| Figura 2. | Abundancia promedio de hormigas (identificados hasta nivel de género) a diferentes distancias del árbol madre (promedio ± E.E). |
Sin embargo, con relación al hábitat la abundancia de hormigas para Pheidole sp.1 varió significativamente (ANOVA F(3,3,01), P= 0.04) entre árboles aislados (6.80±1.53) y áreas abiertas de sabana (1.89±0.66) (Games-Hovell test P= 0.01) siendo similar en parches con G. viburnoides (9.93±4.56) (P=0.76) y parches sin G. viburnoides (3.60±1.15) (P=0,13). Por el contrario, para Pheidole sp.2 no se detectaron diferencias en su abundancia entre los distintos hábitats (ANOVAF(3,1,65), P=0.20)(Fig.3).
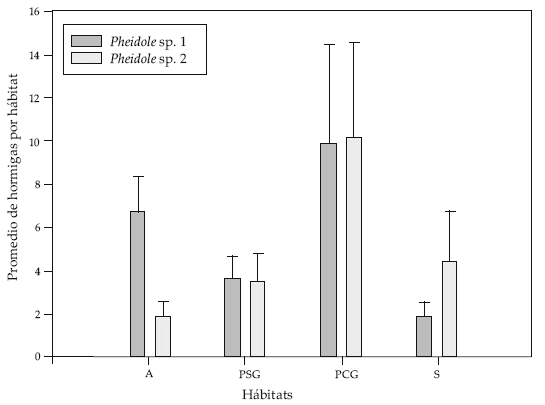 | |
| Figura 3. | Abundancia promedio de hormigas (identificados hasta género) en diferentes hábitats (promedio ± E.E). Donde, A= árbol, PSG = parche sin G. viburnoides, PCG parche con G. viburnoides y S = sabana. |
Discusión
El patrón de depredación encontrado en este estudio revela que los niveles de depredación de semillas de G. viburnoides fueron más altos debajo del árbol madre, que lejos de éste. Este resultado sugiere un efecto distancia-dependiente y concuerda con otros trabajos en los trópicos (Augspuger 1982, Levey & Byrne 1993, Schupp 1995, Pizo & Oliveira 1998, Wenny 2000, Wyatt & Silman 2004, Norghauer et al. 2006, Takeuchi & Nakashizuka 2007) que respaldan el modelo de Janzen (1970) y Connell (1971). El patrón encontrado para G. viburnoides se ve respaldado por factores que son los responsables de generar dicho patrón. Uno de estos factores es la alta densidad de semillas disponibles debajo del árbol madre por efecto directo de la sombra de semillas o indirectamente por la remoción de frutos causada por sus principales dispersores (Laman 1996). En el área de estudio el 91.7% de las semillas es dispersada hasta los 10 m del árbol madre (lluvia de semillas) por aves frugívoras, como Cyanocorax cyanomelas (Loayza &Knight 2010; Saavedra 2008). Esto se traduce en una gran cantidad de semillas disponibles cerca o debajo del parental que atrae a numerosos consumidores de semillas (Janzen 1970, 1971, Wilson & Traveset 1992, Wenny 2000, Levin et al. 2003). Sin embargo, no se detectaron diferencias significativas en la abundancia de depredadores (Pheidole) respecto a la distancia, lo cual contrasta la teoría de atracción de los depredadores hacia sitios con alta concentración de semillas.
En base a observaciones de campo y experimentos piloto, creemos que lo más probable es que hormigas granívoras del género Pheidole sean los depredadores más importantes de esta especie, pero no se descarta la existencia de otros. En términos de abundancia, el género Pheidole es el más diverso y abundante en los trópicos (Holldobler & Wilson 1990, Belstermeyer et al. 2000) tendiendo a concentrar su actividad cerca del árbol en fructificación (Janzen 1970, Takeuchi & Nakashizuka 2007). Encontramos que los depredadores de G. viburnoides consumen las semillas en los sitios donde han sido encontradas. Este comportamiento se ve reflejado en la forma cómo eran consumidas las semillas, donde, un 65% de los endocarpos ofrecidos en los experimentos presentaban orificios por donde las semillas habían sido consumidas (F. Saavedra datos no publicados). Avgar et al. (2008) encontraron que las características de forrajeo de dos especies de hormigas granívoras pueden explicar y pronosticar la estructura espacial de plantas. De la misma manera, estudios en sistemas tropicales encontraron que especies del género Pheidole varían en su forma de consumir las semillas. Mientras unas especies trasladan las semillas al nido (Levey & Byrne 1993; Passos & Oliveira 2002), otras consumen las semillas en el lugar donde las encuentran (Pizo & Oliveira 1998, Passos & Oliveira 2002), movilizando una gran cantidad de individuos (Carroll & Janzen 1973). Al igual que el trabajo de Pizo & Oliveira (1998), observamos una gran cantidad de hormigas consumiendo las semillas en los sitios donde fueron encontradas. Por lo tanto, es posible proponer que el comportamiento alimenticio de los depredadores podría ser otro factor más, detrás del patrón de depredación de G. viburnoides. Con relación al hábitat los resultados encontrados, concuerdan con otros estudios en sabanas tropicales, donde se encontró que la pérdida de semillas debido a la actividad de depredadores fue mayor debajo de arbustos altos comparados con pastizales abiertos, debido a la desproporcional llegada de semillas a los diferentes tipos de hábitats (Holl 2002). Esto podría deberse a que hormigas neotropicales del género Pheidole tienden a ocupar ambientes con abundante hojarasca y materia orgánica donde construyen sus nidos y obtienen alimento (Holldobler & Wilson 1990; Reyes-Lopez etal. 2003). Nosotros encontramos mayor abundancia de hormigas (Pheidole sp.1) en árboles aislados que áreas de sabana y al igual que en nuestro sistema de estudio, Dunn (2000) y Reyes-Lopez et al. (2003) concluyeron que la riqueza y abundancia de hormigas es mayor en árboles aislados que en áreas abiertas. Esto demuestra que nuestro sistema de estudio los depredadores de semillas encontrados pueden jugar un rol crucial en determinar la estructura espacial y composición de comunidades vegetales.
Conclusiones
El patrón de depredación encontrado en este estudio sugiere un efecto distancia-dependiente, que apoya la hipótesis de Escape de Howe & Smallwood (1982) y que la estructura de parches, árboles solitarios y sabana del hábitat influyen sobre la abundancia de hormigas a pequeña escala. La densidad de hormigas (Pheidole spp.) no determina el patrón espacial de depredación de semillas de Guettarda viburnoides.
Agradecimientos
Queremos dar las gracias a Nataniel Burgos, Juanita Balderrama y Alberto Balderrama por el apoyo en la realizaron del trabajo de campo. A Flavia Montaño y Daniel Larrea por su ayuda en el trabajo de clasificación e identificación de hormigas. Al personal de la Estación Biológica del Beni (EBB) por el apoyo logístico. Este estudio fue financiado por el Instituto de Ecología de la Universidad Mayor de San Andrés (UMSA).
Referencias
Augspurger, C. K. 1982. A cue f or synchronous flower. pp. 133-150 En: Leigh, J., E. Giles, Rand A. S. &D. M. Windsor (eds) Ecology of a Tropical Forest Seasonal Rhythms and Long-term Changes. Smithsonian Institution Press, Washington DC. [ Links ]
Avgar, T., I. Giladi, & R. Nathan. 2008. Linking traits of foraging animals to spatial patterns of plants: social and solitary ants generate opposing patterns of surviving seeds. Ecology Letters 11: 224-234.
Beckage, B. &J.S. Clark. 2005. Does depredation contribute to tree diversity? Oecologia 143: 458-469.
Bestelmeyer, B. T., D. Agosti, L. E. Alonso, C. R. Brandao, W. Brown, J.H.C. Delabie & R. Silvestre. 2000. Field techniques for the study of ground-dwelling ants: an
overview, description, and evaluation. pp. 122-144. En: Agosti, D., J. Majer, L. Alonso & T. Schuktz (eds) Ants: Standard Methods for Measuring and Monitoring Biodiversity. Smithsonian Institution, Washington DC.
Carroll, C. R. & D. H. Janzen. 1973. Ecology of foraging by ants. Annual Review of Ecology and Evolution and Systematics 4: 231-257.
Chapman, C. A. 1989. Primate seed dispersal the fate of dispersed seeds. Biotropica 21:148-154. [ Links ]
Comiskey, J. A., F. Dallmeier & G. Aymard. 2000. Floristic composition and diversity of forested habitats in the Estación Biológica del Beni. pp. 89-112. En: Herrera-MacBryde, O., F. Dallmeier, J. A. Comiskey & C. Miranda (eds.) Biodiversidad, Conservación y Manejo en la Región de la Reserva de la Biosfera Estación Biológica del Beni, Bolivia. SI/ MAB Series 4. Smithsonian Institution, Washington DC. [ Links ]
Connell, J. H. 1971. On the role of natural enemies in preventing competitive exclusion in some marine animals and in rain forest trees. pp. 298-312. En: Den Boer, P.J. & G.R. Gradwell (eds). Dynamics of Populations. Center for Agricultural Publication and Documentation, Wageningen. [ Links ]
Crawley, M. J. 1993. GLIM for Ecologists. Blackwell Scientific Publications, Oxford. 379 p. [ Links ]
Dunn, R. R. 2000. Isolated trees as foci of diversity in active and fallow fields. Biological Conservation 95: 317-321. [ Links ]
Fernández, F. (ed.). 2003. Introducción a las hormigas de la región neotropical. Instituto de Investigación de Recursos Biológicos Alexander Von Humbolt, Bogotá. 456 p.
Ferreira, A. V., M. E. Bruna & H. L. Vasconcelos. 2011. Seed predators limit plant recruitment in Neotropical savannas. Oikos 120:1013-1022.
García, D., J. R. Obeso & I. Martínez. 2005. Spatial concordance between seed rain and seedling establishment in bird-dispersed trees: does scale matter? Journal of Ecology 93: 693-704.
Harms, K. E., S. J. Wright, O. Calderón, A. Hernández & E. Allen Herre. 2000. Pervasive density-dependent recruitmentenhances seedling diversity in a tropical forest. Nature 404:493-495.
Holl, K. D. 2002. Effect of shrubs on tree seedling establishment in an abandoned pasture. Journal of Ecology 90:179-187. [ Links ]
Holldobler, B. & O. Wilson. 1990. The ants. Belknap Press of Harvard University Press, Cambridge. 774 p.
Hulme, P. E. 1998. Post-dispersal seed predation and seed bank persistence. Seed Science Research 8: 513-519 [ Links ]
Hulme, P. E. 2002. Seed-eaters: dispersal, destruction and demography. pp. 257-273. En: Levey, D. J., W. R. Silva & M. Galetti (eds.), Seed Dispersal and Frugivory: Ecology, Evolution and Conservation. CAB International, Wallingford. [ Links ]
Hulme, P. E. &C. W. Benkman, 2002. Granivory. pp. 132-154. En: Herrera, C. & O. Pellmyr (eds.) Plant-animal Inter actions: An E volutionary Approach. Blackwell, Oxford. [ Links ]
Howe, H. F., E. W. Schupp & L. C. Westley. 1985. Early consequences of seed dispersal for a Neotropical tree (Virola surinamensis). Ecology 66:781-791.
Janzen, D. H. 1970. Herbivores and the number of tree species in tropical forests. The American Naturalist 104: 501-528. [ Links ]
Janzen, D. H. 1971. Seed predation by animal. Annual Review of Ecology and Evolution and Systematics 2: 465-492. [ Links ]
Jones, F. A., C. J. Peterson & B. L. Baines. 2003. Seed predation in Neotropical pre-montane pastures: Site, distance and species effect. Biotropica 35:219-225.
Laman, T. G. 1996. Fig seed shadows in a Bornean rain forest. Oecologia (Berl.) 107: 347-355. [ Links ]
Levey, D. & M. Byrne. 1993. Complex ant-plant interactions: rain forest ants as secondary dispersers and post-dispersal seed predators. Ecology 74:1802-1812.
Levin, S. A., H.C. Muller-Landau, R. Nahan & J. Chave. 2003. The ecology and evolution of seed dispersal: a theoretical perspective. Annual Review of Ecology Evolution and Systematics 34: 575-604.
Lindsey, P. A. & J. D. Skinner. 2001. Ant composition and activity patterns as determinedby pitfall trapping and other methods in three habitats in the semi-arid Karoo. Journal of Arid Environments 48: 551-568.
Loayza, A. P. & T. Knight. 2010. Seed dispersal by pulp consumers, not "legitimate" seed dispersers, increases Guettarda viburnoides population growth. Ecology 91:2684-95.
Loayza, A. P., B. A. Loiselle & R. S. Rios. 2011. Context-dependent recruitment of Guettarda viburnoides in a Neotropical forest-savannamosaic. American Journal of Botany 98:1317-1326.
Louda, M. A., M. A. Farris & M. J. Blua. 1987. Variation in methylglucosinolate and insect damage to Cleome serrulata (Capparaceae) along a natural soil moisture gradient. Journal of Chemical Ecology 13: 569-581.
Miranda, C. L. 2000. La Reserva de la Biosfera Estación Biológica del Beni, Bolivia. pp. 21-25. En: Herrera-MacBryde, O., F. Dallmeier, B. MacBryde, J. L. Comiskey & C. Miranda (eds.) Biodiversity, Conservation and Management in the Region of the Beni Biological Station Biosphere Reserve, Bolivia. Smithsonian Institution/Monitoring and Assessment of Biodiversity Program (SI/MAB), Washington DC. [ Links ]
Maron, J. R. & E. L. Simms. 2001. Rodent-limited establishment of bush lupine: field experiments on the cumulative effect of granivory. Journal of Ecology 89:578-588.
Norghauer, J., J. R. Malcolm, B. L. Zimmerman & J. M. Felfili. 2006. An experimental test of density- and distant-dependent recruitment of mahogany (Swietenia macrophylla) in southeastern Amazonia. Oecologia (Berlin) 148:437-446.
Orrock, J.L., D. J. Levey, B. J. Danielson & E. I. Damschen. 2006. Seed predation, not seed dispersal, explain the landscape-level abundance of an early-successional plant. Journal of Ecology 94: 838-845.
Passos, L. &P. S. Oliveira. 2002. Interaction between ants and fruits of Guapira opposite (Nyctaginaceae) in a Brazilian sandy plain rainforest: ant effects on seeds and seedlings. Oecologia 139: 376-382.
Pizo, M. A. & P. S. Oliveira. 1998. Interaction between ants and seeds of a non myrmechorous Neotropical tree, Cabralea canjerana (Meliaceae), in the Atlantic forest of southeast Brazil. American Journal of Botany 85: 669-674.
PL-480&ANCB(Academia Nacional de Ciencias de Bolivia). 1998. Plan Quinquenal EBB 2000. Estrategia hacia un futuro sostenible. La Paz. 27 p. [ Links ]
Reyes-Lopez, J., N. Ruiz & J. Fernandez-Haeger . 2003. Community structure of ground- ants: T role of single trees in a Mediterranean pastureland. Acta Oecologica 24: 195-202.
Romero, H. & K. Jaffe. 1989. A comparison of methods for sampling ants (Hymenoptera, Formicidae) in savannas. Biotropica 21(4): 348-352.
Saavedra, F. 2008.Patrones de reclutamiento -dispersión, sobrevivencia y establecimiento- de Guettarda viburnoides (Rubiaceae) En sabanas de la Reserva de la Biosfera Estación Biológica del Beni (RB-EBB). Tesis de licenciatura en biología, Universidad Mayor de San Andrés, La Paz. 79 p. [ Links ]
Schupp, E.W. 1990. Annual variation in seedfall, postdispersal predation, and recruitment of a Neotropical tree. Ecology 71: 504-515. [ Links ]
Schupp, E. W. 1992. The Janzen-Connell model for tropical tree diversity: population implications and the importance of spatial scale. American Naturalist 140: 126-130. [ Links ]
Schupp, E. W. 1995. Seed-seedling conflicts, habitat choise, and patterns of plant recruitment. American Journal of Botany 82: 399-409. [ Links ]
Schupp, E. W. 1988. Seed and early seedling predation in the forest understory and in treefall gaps. Oikos 51:71-78. [ Links ]
Takeuchi, Y. & T. Nakashizuka. 2007. Effect of distance and density on seed/seedling fate of two dipterocarp species. Forest Ecology and Management 247:167-174.
Taylor, C., P. G. Delprete, A. Vincentini, R. Cortés, D. Zappi, C. Persson, C. Bestetti Costa & E. Araujo de Anunciação. 2004. Rubiaceae. Pp. 497-847. En: Steyermark, A., P. E. Berry, K. Yatskievych & B. K. Holst (eds.) Flora of the Venezuelan Guayana, vol. 8: Poaceae - Rubiaceae, Missouri Botanical Garden Press, St. Louis. [ Links ]
Wenny, D. G. 2000. Seed dispersal, seed predation, and seedling recruitment of a Neotropical montane tree. Ecological Monographs 70: 331-351. [ Links ]
Wilson, M. F. & A. Traveset. 1992. The ecology of seed dispersal. Pp. 85-110. En: Fenner, M. (ed.) Seeds: The Ecology of Regeneration in Plant Communities. 2da. edic. CAB International, Wallingford. [ Links ]
Wyatt, J. & M. R. Silman. 2004. Distance-dependence in two Amazonian palms: effects of spatial and temporal variation in seed predator communities. Oecologia 140:26-35.
Artículo recibido en: Noviembre de 2011.
Manejado por: Javier Lopez de Casenave
Aceptado en: Mayo de 2012.

