











 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkEcología en Bolivia
versión impresa ISSN 1605-2528versión On-line ISSN 2075-5023
Ecología en Bolivia v.47 n.1 La Paz abr. 2012
Artículo
Survival and growth of big-leaf mahogany (Swietenia macrophylla King) seedlings in two provenance trials in Bolivia
Supervivencia y crecimiento de plántulas de mara (Swietenia macrophylla King) en dos ensayos de procedencia en Bolivia
Ernesto Escalante1,2*, Valentín Pando1, Pedro Saravia3 & Felipe Bravo1,2
1Instituto Universitario de Investigación en Gestión Forestal Sostenible, Universidad de Valladolid -Instituto Nacional de Investigación y Tecnología Agraria y Alimentaria (INIA), Av. de Madrid 44, 34004 Palencia, España.
*Corresponding author: E-mail address: escalante.ernesto@gmail.com
2Departamento de Producción Vegetal y Recursos Forestales, Universidad de Valladolid, Av. de Madrid 44, 34004 Palencia, España.
3Carrera de Ingeniería Forestal, Universidad Autónoma Gabriel René Moreno, Facultad de Ciencias Agrícolas, El Vallecito Km. 9 carretera al Norte, CC. 702, Santa Cruz, Bolivia.
Abstract
The conservation of Bolivian big-leaf mahogany (Swietenia macrophylla King.) forests requires of provenance trials to identify potential seed sources. Survival and early growth of seedlings are regarded as key variables in provenance trials. Two provenance trials were installed in an open secondary forest. Five Bolivian mahogany provenances, which belong to the pre-Andean Amazon (A and SC), Chiquitano-Amazon transitional (B and CH) and the Amazon (C) ecoregions, were evaluated. Mixed models with repeated measurement data were used to analyze survival rate, height growth and root collar diameter of seedlings, as well as seedling vigour. After 12 months of plantation, survivorship of all provenances was much lower at site 1 than at site 2. Height growth was relatively uniform among provenances and sites, ranging from 33-47 cm and from 63-101 cm, respectively. After 19 months at site 2, the provenances from Chiquitano-Amazon transitional (B=84%, N=288) and pre-Andean Amazon (SC=79.5%, N=288) showed the highest survival rates, whereas differences in height growth varied from 150 to 216 cm. In general, root collar diameter growth was lower at site 1 than at site 2. Among provenances, B and CH were significantly different in diameter growth after 4 months at site 1, however, no differences were observed at 12 months among all provenances. At site 2, diameter growth showed that provenances B and CH did better after 4 months while CH did better after 12 and B after 19. In summary, the provenance B from Chiquitano-Amazon transitional showed better characteristics than the rest of tested provenances (84% survival, 135.1 cm/year height growth and 34.7 mm/year root-collar diameter). Our findings provide useful information on survival and growth performance of Bolivian mahogany in plantations on secondary forests.
Keywords: Ecoregion; Height growth; Root collar diameter; Survival rate; Tropical timber species.
Resumen
Se instalaron dos ensayos de procedencia en un bosque secundario abierto. Se evaluaron cinco procedencias de mara (Swietenia macrophylla King) pertenecientes a la ecoregión pre-andina amazónica (A y SC), transición chiquitano-amazónica (B y CH) y amazónica (C). Se utilizaron modelos mixtos con datos de medidas repetidas para analizar la tasa de sobrevivencia, crecimiento en altura y diámetro raíz de cuello de plántulas, como también el vigor de las plántulas. La supervivencia varió entre las procedencias y fue inferior en el sitio 1 que en el sitio 2, después de 12 meses de la plantación. El crecimiento en altura fue relativamente uniforme entre procedencias y sitios, variando desde 33-47 cm y desde 63-101 cm, respectivamente. Después de 19 meses en el sitio 2, la procedencia de la ecoregión de transición chiquitano-amazónica (B=84%, N=288) y pre-andina amazónica (SC=79.5%, N=288) mostraron altas tasas de supervivencia, mientras que las diferencias de crecimiento en altura variaron desde 150-216 cm, respectivamente. En general, el crecimiento del diámetro a la raíz del cuello fue menor en el sitio 1 que en el sitio 2. Entre procedencias, la procedencia B y CH fueron significativamente diferentes después de cuatro meses en el sitio 1; sin embargo, no se observaron diferencias entre el resto de las procedencias a los 12 meses. En el sitio 2, el crecimiento en diámetro fue mejor para las procedencias B y CH a los cuatro meses, mientras CH lo hizo a los 12 y B a los 19. En resumen, la procedencia B de la ecoregión de transición chiquitano-amazónica mostró mejores características que el resto de las procedencias ensayadas (sobrevivencia 84%, crecimiento en altura 135.1 cm/año y diámetro raíz de cuello 34.7 mm/año). Nuestros resultados proporcionan una información útil sobre la supervivencia y el crecimiento de las plantaciones de mara en un bosque secundario abierto.
Palabras claves: Crecimiento en altura, Diámetro raíz de cuello, Ecoregión, Especie maderable tropical, Tasa de sobrevivencia.
Introduction
In forestry, provenance research provides comprehensive knowledge about adaptation of the species to specific locations and provides information on genetic and environmental variability (Callaham 1963). Big-leaf mahogany, Swietenia macrophylla King, Meliaceae family (hereafter referred as Mahogany), is a valuable tropical timber tree that is naturally distributed over regions of Central and South America, from Mexico to Bolivia. Mahogany's natural populationinPeru,BoliviaandBrazilisrestricted to southern areas of the Amazonian forests. In Bolivia, mahogany occurs over reduced areas of the Amazon, pre-Andean Amazon and the Chiquitano-Amazon transitional ecoregion (Mejía et al. 2008). Mahogany populations are threatened in South America, as natural populations have been severely affected by several factors, including deforestation (Gillies et al. 1999) and selective logging (Veríssimo et al. 1995, Krometter et al. 2004) which have negatively impacted on the genetic structure of populations (Gillies et al. 1999, Novick et al. 2003). All mahogany populations are included in Appendix III of the Convention on International Trade in Endangered Species of Wild Fauna and Flora (CITES) in countries of Central America (Costa Rica and Mexico) and South America (Brazil, Bolivia, and Peru). In Bolivia, this species has been included in the CITES list since 1998 and in the Appendix II since 2003, which promotes legal harvesting and non-detrimental to the survival of the species in its natural environment. Mahogany logging began in Bolivia in the late 1960s (Krometter et al. 2004) and was held until the mid 1990s (Bascopé 1992) promoted by the high price of timber in international markets. During that period, commercial logging in Bolivia was highly focussed on mahogany (Mostacedo & Fredericksen 1999) and accounted for 90% of total Bolivian exports (Merry & Carter 2001). As a result of this selective harvesting, the genetic constitution of mahogany populations may have been adversely affected (Patiño Valera 1997). Even though very little research on forest management has been traditionally undertaken in Bolivia, interest has increased lately, probably due to new management strategies implemented in forestry. A new forestry law (Forestry Law Nº 1700) established the legal minimum felling size in Bolivia at 70 cm diameter at breast height, and had a very positive effect on mahogany conservation (Mejía et al. 2008), but usage and conservation of forest genetic resources are still being discussed. Therefore, the efforts for the conservation of forest resources in Bolivia are still seen as insufficient.
Previous works on mahogany have been focussedon timber production (Varmola& Carle 2002, Wadsworth &González 2008), silvicultural systems (Browder et al. 1996, Mayhew et al. 2003), natural regeneration (Gullison & Hubbell 1992, Mostacedo & Fredericksen 1999) and pathologies (Weaver & Bauer 1986, Pérez-Salicrup & Esquivel 2008). Small plantations of mahogany have been established across its natural distribution range, mainly in local communities and privately owned land, but these are often poorly managed. Sites selected for trials are areas known to belong to the previous mahogany distribution range thus considered as potential sites for the plantation of this species. In the Brazilian Amazon three mahogany planting regimes are found among small-scale family farmers, degraded fallow enrichment, agroforestry association, and pure stand-plantation (Browder et al. 1996), whereas line-plantations are common in Puerto Rico (Weaver & Bauer 1986).
Many researchers have reported on the difficulties and requirements for natural regeneration of mahogany (Gullison & Hubbell 1992, Veríssimo et al. 1995, Gullison et al. 1996, Grogan et al. 2003). Alternatively, outplanting programs may be needed to ensure successful regeneration of this valuable species (Park et al. 2005). However, in order to obtain success with hardwood plantations in the tropics and sub-tropics (Varmola & Carle 2002, Piotto et al. 2004, Wadsworth & González 2008), either as mixed or pure plantations (Piotto et al. 2004) appropriate knowledge about the species involved is required. Thus, understanding the response of mahogany provenances is central to forestry, particularly in sustainable forest management and conservation. The success of the establishment of plantations depends on plantation design and adaptability of the provenances to be used (Callaham 1963, Wormald 1992). Unfortunately, very little information on experiences about provenance trials of mahogany can be found in the literature. Trials were established to assess differences in seedling performance of five provenances towards the conservation and improvement of big-leaf mahogany (S. macrophylla). In this study we evaluate survival, early tree-seedling growth, root collar diameter, and vigour of planted seedlings of five mahogany provenances at two experimental sites and at three dates after plantation.
Methods
Study site
Experiments were carried out at the southern part of El Chore Forest Reserve of Permanent Production (hereafter referred to as the Chore Reserve), located 120 km north of Santa Cruz, Bolivia (Fig. 1). Field trials were established at two sites: Pulpito (16°58'S, 63°57'W); hereafter referred to as site 1 and Chapaco (16°56'S, 63°43'W); hereafter referred to as site 2 located 25 km apart within the Chore Reserve. Within an annual mean temperature of ~24°C and an annual mean precipitation of 1212.7 mm/ year (since 1961-1990) there is no considerable difference in climate between the two sites. The life-zone ecology of the Chore Reserve has been classified as tropical rain forest in the northern area and subtropical rain forest in the southern section (Holdridge 1967). The sites have an average elevation of 400 m (range 320-460 m), flat topography, and a well-drained mineral soil, which is appropriate for mahogany as it grows well on a wide range of soils (Lamb 1966).
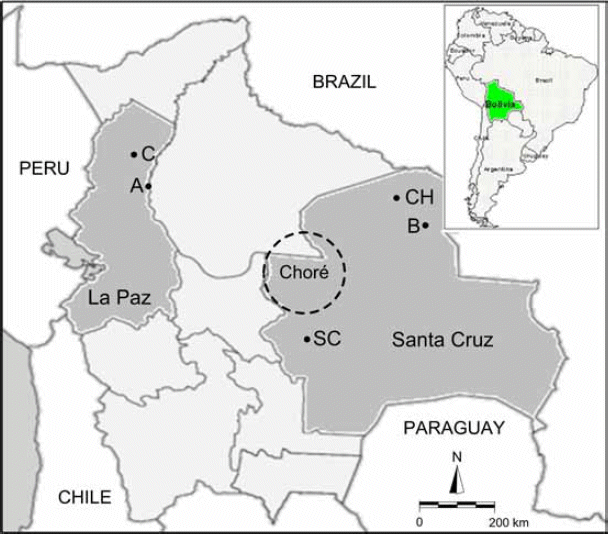 | |
| Figure 1. | Geographical location of the five provenances studied from three ecoregions of Bolivia. Locations of the provenance trials of mahogany plantations at two experimental sites (dashed line of the circle). Details of provenances are given in table 1. |
| Table 1. | Mahogany (S. macrophylla King) provenances and site characteristics where the seeds were collected. Elevation (m), mean annual temperature (degree), and total annual precipitation (mm) obtained from the closest meteorological stations where seeds of mahogany were collected. |
 | |
The Chore Reserve includes forest species of medium commercial value, such as Hura crepitans (Euphorbiaceae), Ficus spp (Mor aceae) or Terminalia oblonga (Combretaceae) (Dauber et al. 1999, Pattie et al. 2003). The estimated abundances of S. macrophylla and Cedrela odorata, two of the main commercial species in the area, were 0.46 tree/ha and 0.53 tree/ha respectively (Dauber et al. 1999). The logging pressure in the Chore Reserve has increased because of intensive timber harvesting on primary forests, swidden fallow, resulting from rotational swidden agriculture on secondary forests surrounding the reserve. Recently, Pinto (2006) reported on the evolution of the landscape, deforestation rates and conservation condition in the Chore Reserve.
Seed collection
Seeds were collected from healthy mahogany trees located along the Pre-Andean Amazon, Chiquitano-Amazon transitional, and the Amazon ecoregions (Dauber et al. 2003, Mejía et al. 2008) in Bolivia. The main characteristics of these three ecoregions are: the Pre-Andean Amazon ecoregion covers large areas over the mid-elevation premontane humid forests; the Chiquitano-Amazon transitional ecoregion is a transitional area between the humid climate of the Amazonian region and the dry climate of the Chaco, with a seasonal climate; and the Amazon ecoregion is found in the lowland rainforest located in the southern part of the Amazonian forests, commonly described as tropical lowland humid forests. Seeds were collected from felled or standing trees. Seed provenances and sites collected are illustrated in fig. 1. Seedlings were grown for 12 months in a nursery, and then outplanted to field trials. Before plantation, mean height of seedlings were 22.5 cm (range 20.1-25.3 cm) and 24.2 cm (range 19.8-26.7 cm) at sites 1 and 2 respectively.
Experimental design
Seedlings were planted at the two sites under open secondary forest conditions. A total of 71 subplots were replicated in contiguous blocks at the time of planting, of which 47 were located in site 1 and 24 in site 2. The mahogany seedlings were planted at 5 m x 5 m spacing (36 seedlings per subplot). A total of 2,556 mahogany seedlings were planted in both field trials. Giant Chinaberry tree (Melia azedarach L. var. gigantea; Meliaceae), which is a fast-growing and commercial species, was inter-planted in the middle of four mahogany seedlings (25 seedlings per subplot). Five mahogany provenances were established at site 1 and four at site 2 owing to insufficient collection of seeds and limited germination of seedlings. Treatments followed a randomized block design in the field. No fertilizers or insecticides were applied at any time.
Field measurements
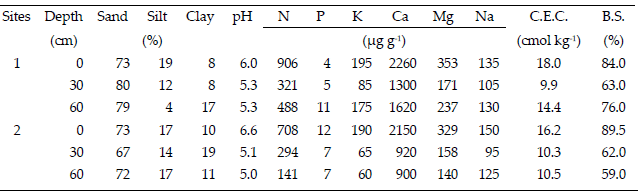
Survival, total height, RCD (basal diameter) and vigour of mahogany seedlings were measured at 4 and 12 months after plantation in site 1 and at 4, 12, and 19 months in site 2. Seedling height was considered as the distance measured from the root collar to the apical shoot tip. Survival rate was measured as the proportion of living seedlings per number of planted seedlings. Mean seedling height was calculated by subplots in each site. RCD was measured in two perpendicular directions for each seedling using a vernier caliper after planting. Vigour was evaluated based on morphological characters of seedlings according to Malleux (1982), where 1 is a good value and more than 1 indicated that the seedling vigour is reducing. In this study, the effect of M. azedarach seedlings has not been taken into account because its main purpose was to evaluate the performance range of mahogany seedlings among provenances. Soil samples were collected from two representative spots at each site at three depth levels (0,30 and 60 cm). Minimum post-plantation maintenance was carried out, but this was eventually abandoned after 4 months.
| Table 2. | Soil texture and chemical composition of soils from two experimental sites, named Pulpito (site 1) and Chapaco (site 2). Data represent mean values from two samples at each depth level. Legend: C.E.C.=Cation Exchange Capacity; B.S.=Base Saturation. |
 | |
Statistical analyses
Differences among provenances regarding seedlings survival, height, root collar diameter, and vigour were analyzed using a linear mixed model (LMM) in the PROC MIXED procedure (SAS Institute Inc 2004, Cary, NC). An analysis of variance was carried out using localities (sites), provenances, blocks and measurements as fixed effects. We did not use random effects due to the unstructured dataset.Themodelization was performed using the restricted maximum likelihood (REML) approach and an unstructured variance-covariance matrix by sites. Independence of observations and normality of the error residuals were assumed in the model. Least-square (LS) mean values by provenances, subplots and sites were calculated. The significance level for all tests was set to P<0.05. Survival data was used as a continuous variable.
Results
Survival rate
Seedling survival rates were significantly different among measurement dates (Table 3; fig. 2). From 4-12 months, survival of seedlings reduced significantly from 72.2-26.9% at site 1, and from 91.7-78% at site 2. After 12 months at site 1, survival rates of provenances C (39.7%) and CH (44.2%) were significantly higher than the rest. After 12 months at site 2, seedlings survival ranged from 66-84.7%. Among provenances, seedling survival was higher for B (84.7%) and SC (80.9%) provenances, but no significant differences were observed between them. Furthermore, survival rates among provenances from the Pre-Andean Amazon ecoregion were not statistically different (A and SC; table 3). However, differences between provenances B and CH (Chiquitano-Amazon transitional ecoregion) were highly significant (P=0.0036). After 19 months at site 2, overall survival rate for all provenances was 76.3% (Table 3). Provenances A (68.1%) and CH (62.5%) had poorer survival rates than B and SC provenances (84% and 79.5% respectively), but no significant differences between each pair of provenances were observed (Fig. 2).
| Table 3. | Least square means of survival rate, height, root collar diameter (RCD), and vigour by provenances (mean ± SE). Tests were performed at the 5% level of significance. Distinct capital letters in the rows indicate significant differences among sites and/or measurements corresponding to each provenance, respectively. Distinct lowercase letters in the columns indicate significant differences among provenances. The asterisk (*) indicates the overall mean values by sites. |
 | |
| Figure 2. | Survival rates of mahogany seedlings among provenances and sites over the studied period. The provenances tested belong to the following ecoregions of Bolivia, Pre-Andean Amazon (A and SC), Chiquitano-Amazon transitional (B and CH), and Amazon (C). |
 | |
Height growth
Differences in mean-height of seedlings among provenances were significant in both sites (P<0.0001) (Table 3; fig. 3). After four months (site 1), mean height of provenances was 29.5±0.3 cm. There were significant differences in height among provenances, although no significant differences were found between the two pairs of provenances (A, CH and C,SC), which were similar. At site 2, height of provenance B (51.2±2.1 cm) was significantly higher than the rest. After 12 months (site 1) height was on average 38.9±0.9 cm, and only provenance B (47.4±2 cm) showed a highly significant difference with the rest. At site 2, height growth among provenances varied from 62.8-100.6 cm, with a mean of 81.3±3.6 cm for all provenances. At this period, provenances A and B were taller than CH and SC provenances, and were significantly different among them. Mean height growth in provenances B and A were 100.6±6.2 cm and 83.2±8.8 cm respectively. Overall, seedlings of provenance B were taller at site 2 than at site 1 after 12 months, followed by the seedlings of provenance A at site 2 and SC at site 1. Finally, remarkable differences in seedling height growth among mahogany provenances were observed at 19 months, although no statistically significant differences between provenances A, B and CH were found (Table 3; fig. 3). Total height ranged between 150±15.5 cm (SC) and 216.1±15.5 cm (B).
| Figure 3. | Height growth of mahogany seedlings among provenances and sites over the studied period. The provenances tested belong to the following ecoregions of Bolivia, Pre-Andean Amazon (A and SC), Chiquitano-Amazon transitional (B and CH), and Amazon (C). Different lowercase letters indicate the significant differences between mean values among provenances tested at each time point and sites (1, 2). |
 | |
Root collar diameter growth
As shown in table 3, there were significant differences among sites for RCD (P<0.0001). Among provenances (Fig. 4), none of them showed significant differences after 12 months at site 1, however, the CH provenance was significantly high at site 2 (site 1: ranged from 5.9-7 mm and site 2: ranged from 10.9-29 mm). After 19 months at site 2, RCD ranged from 33.8-55.5 mm. The root collar diameter growth for the B provenance (55.5±3.6 mm) was significantly higher from the rest. However, provenances A, CH and SC were no statistically different among them.
| Figure 4. | Root-collar diameter of mahogany seedlings among provenances and sites over the studied period. The provenances tested belong to the following ecoregions of Bolivia, Pre-Andean Amazon (A and SC), Chiquitano-Amazon transitional (B and CH), and Amazon (C). Different lowercase letters indicate the significant differences between mean values among provenances tested at each time point and sites (1, 2). |
 | |
Seedling vigour
There were significant differences in seedling vigour between site 1 and 2 after four months, as shown in table 3. At site 1, seedling vigour decreased between two periods studied, although only provenances CH and SC were significantly different after 12 months. After 4, 12 and 19 months at site 2, there were no differences between the overall mean of seedlings vigour, and the vigour index for all provenances was improved in each period (Table 3; fig. 5). As illustrated in fig. 5, the vigour indices of seedlings were relatively similar among provenances, and were not significant among them, at 12 and 19 months after plantation in site 2.
| Figure 5. | Vigour index of mahogany seedlings among provenances and sites over the studied period. The provenances tested belong to the following ecoregions of Bolivia, Pre-Andean Amazon (A and SC), Chiquitano-Amazon transitional (B and CH), and Amazon (C). Different lowercase letters indicate the significant differences between mean values among provenances tested at each time point and sites (1, 2). |
 | |
Finally, soils were characterized as sandy loam and slightly acidic. With respect to major nutrients such as calcium (Ca+) and potassium (K+) there were higher values in site 1 than in site 2. Concentrations of exchangeable bases (Ca+, Mg+, K+ and Na+) showed similar results for both sites, but the concentrations of N+, Ca+ and Mg+ decreased with soil depth and sites (Table 2).
Discussion
Survival, early growth, root collar diameter and vigour Early survival and vigorous growth are essential to ensure mahogany establishment (Lamb 1966, Gullison et al. 1996). In our study, the proportion of seedlings surviving at the latest census (19 months, site 2) was 76.3% (Table 3). Our results are partly consistent with other findings in natural forests, for instance, survival rate was quite similar to that reported for Brazilian (61-64% in gaps and 25% in the forest understory after 42 months) and lower than in Mexican (87.4% in new felling gaps after 20 months) natural forests (Dickinson & Whigham 1999, Grogan et al. 2003). On the contrary, in a closed forest of Pará (Brazil), survivorship of big-leaf mahogany after natural regeneration was 18.2% after 10 months (Grogan et al. 2006), and in Quintana Roo (Mexico) only 5% of mahogany seedlings survived under the canopy 58 months after establishment (Snook & Negreros-Castillo 2004). These studies showed few encouraging results for natural forests, probably due to large differences in growth rates for individual trees between regions (Gullison & Hubbell 1992, Gullison et al. 1996). In a comparative study between slow and fast growth tropical species carried out in Costa Rica, survival rates and height of S. macrophylla evaluated 68 months after plantation were lower in pure stands (62% and 4.92 m) than in mixed plantations (76.2% and 5.10 m) although they did not differ significantly among them (Piotto et al. 2004). An evaluation of mahogany survival made in Quintana Roo (Mexico), found on average only 22% of seedlings survived until the third year, probably because of poor seed quality and deficient planting (Negreros-Castillo & Hall 1996). On experimental plantations, line plantings of mahogany were established in 1974, 1979 and 1980 in Luquillo (Puerto Rico), resulting in surviving seedlings percentages of 66.7, 66.6 and 58.7, respectively (Weaver &Bauer 1986). Survival of S. macrophylla in a secondary dry forest in the Guanacaste Conservation Area (Costa Rica) was found to be 35% after 3 years after establishment (Gerhardt 1996). In our sstudy, herbaceous plants and shrubs species could have influenced survival rate at site 1, as young Swietenia and Cedrela trees are highly intolerant to overhead shade (Lamb 1966), both species require accessibility to light through canopy openings or clearings to regenerate (Gullison & Hubbell 1992, Dickinson & Whigham 1999). As described before, our survival results from site 2 were higher in relation to experiences developed in natural forests and mahogany plantations, but lower when compared to results from site 1. On the other hand, our experiments (site 2) resulted in survival rates over 70%, which can make it possible to turn mahogany small-scale planting into a financially viable operation according to simulations developed in Brazil (Browder et al. 1996).
The observed differences in height growth and RCD among provenances were evident from the beginning of the trials (Table 3; figs. 3 and 4). Height growth varies with age (Snook 2003), size class (Gullison et al. 1996), site quality (Lamb 1966), andinpart, due to genetic variation among mahogany populations (Ward & Lugo 2003). A progeny trial of mahogany in Upala (Costa Rica) showed an overwhelming height growth (Navarro & Hernández 2004). The authors indicated that growth can be considered fast when RCD increases by 20 mm/year and height over 1 m/year. Our results revealed that RCD increased above 34.7 mm/year and over 130 cm/year height (B provenance), and could, therefore be classified as fast growing (site 2). In the Luquillo mountains (Puerto Rico), mean annual height was in the range of 0.7-1.0 m, depending on topography, with growth rates being lower on ridges (Weaver & Bauer 1986). Mahogany trees in plantations typically exhibit diameter growths that exceed 1 cm/year during the early years (Lamb 1966). In a study conducted in Mexico where small-scale plantations of mahogany (0.5-2 ha) were established in areas formerly used for shifting agriculture, tree height growth was 0.5 m/year (Pérez-Salicrup & Esquivel 2008). The magnitude of the differences differed among provenances (mainly seedling height). Differences in RCD, for example, were highly significant (P<0.0001, provenance B only). Light experiments conducted in Sri Lanka (Mahroof & Edirisinghe 2001) reported a higher RCD of S. macrophylla seedlings in open houses than in shade houses, two years after planting. They found high survival rates of S. macrophylla when growing in shade houses irrespective of the shape levels (ranged from 91-98.1%). No significant differences in final height were observed under the three regimes (nine months after planting) and RCD responded to light availability under artificial shade. Among the provenances studied, our results can be summarized by saying that provenance B showed very high survival, height and RCD values, although the variability increased over time. Overall, in our field plantation trials, seedling survivorship varied among provenances, and broadly speaking, higher values were obtained at site 2 (Fig. 2).
Mahogany provenances for forest plantations
Provenance B from Chiquitano-Amazon transitional ecoregion demonstrated a striking performance in survival rate (84%), growth (135.1 cm/year), and RCD (34.7 mm/year) at site 2,19 months after plantation. Many difficulties on gaps after logging, large density variation of seedlings in gaps, or low survival rates of mahogany seedlings, were commonly reported in natural forests (Quevedo 1986, Gullison et al. 1996, Snook & Negreros-Castillo 2004, Grogan et al. 2006), therefore, an alternative for promoting investment on forest plantation is to produce high-quality seedlings, based on tested provenances and followed by appropriate silvicultural and management practices. To improve deterioration of mahogany seedling (Gillies et al. 1999, Krometter et al. 2004), based on our results, provenance B would be a logical candidate for seed planting or seed orchards, and these results could be useful for initial material selection.
Height growth and survival rates were higher for site 2, but further consideration on the observed variability, especially in height is needed. Future trials will also improve our understanding of mahogany provenances in order to effectively manage and conserve the species' genetic resources (Gillies et al. 1999). Successfulforestplantations require monitoring, planning and implementation of treatments to ensure tree seedling establishment in the open plantation or enrichment understory. Our results showed consistent early establishment of mahogany seedlings at site 2 after 19 months. Over the study period, no Hypsiphylla grandella attacks were reported, probably this may have been caused by the vigorous growth of seedlings (site 2), although the attacks over a 0.5 m height were reported (Griffiths 2001). McCarter (1986) indicated that apparent resistance to short borer attack was influenced by seedling vigour. As observed at site 1, the low height during seedling development could have positive effects to avoid the short borer attacks.
The most successful mahogany plantation efforts should be those concentrating investment in areas where nutrient-rich soils are available to boost early seedling growth (Grogan et al. 2003). Although there were not differences in soil texture (sandy loam) in our study (Table 2), Gerhardt (1996) observed that soil texture influences soil water content in both deciduous (clay loam) and semi-evergreen (loam) sites with mahogany plantations in Costa Rica. In our study (site 1), the lower performance of S. macrophylla seedlings could be probably attributed to physical soil properties (soil aeration and moisture retention), therefore these factors could have adversely affected seedling growth and survival. In relation to morphology, mahogany seedlings showed small differences in leaf damage, leaf number and foliar conditions in two soil types in Pará, Brazil (Norghauer et al. 2008), which may have conditioned the vigorously of seedlings. Therefore, it could be necessary to include this variable in future provenance trials to investigate the relationships between seedling performance and soil conditions.
The plantation with native species can help recoverdegradedsecondaryforestsandpromote the rehabilitation of degraded lands, although recovery takes a long time (Kartawinata 1994). Forest managers should, therefore, use well-adapted provenances to ensure the success of the plantation, but additional research in this matter is still needed. Our results suggest that some mahogany provenances (B and CH) seem to be well adapted to the Amazon region (Chore Reserve) and are certainly commercially promising alternatives (Bascopé 1992, Dauber et al. 2003). Additionally, variations in mahogany early growth and survival could be studied in multi-location provenance tests. Given that provenance trials were done in forestry areas and within the species distribution, it would be valuable to undertake further research on other plantation systems such as enrichment planting, mixed-plantations or agroforestry. Until more is known on the response of mahogany plantations, reforestation efforts should focus on enhanced mahogany provenances based upon our initial results.
Acknowledgements
This research was partially supported by the Johnny Mahogany Seed Program in Bolivia. We thank FUNDEFOREST (Fundación para el Desarrollo Forestal) formerly named PROMABOSQUE (Programa de Manejo de Bosques) for seedlings production in the nursery. We would like to thank R. Sierra and B. Turrión, who provided helpful comments on provenances and soils approach. We also thank J. Azevedo and I. Etxebeste for their comments on the manuscript. E. Duran and P. Rodriguez for assistance in data collection and trial establishment. S. Vaca, H. Morales, and D. Taxi, who helped with the fieldwork and L. Gonzales for her collaboration in the soil sample collection.
References
Bascopé, F. 1992. Mahogany (Swietenia macrophylla King) in Bolivia: Its sites, industrial development and actual situation. Mahogany Workshop: Review and Implications of CITES. Tropical Forest Foundation, Washington, DC. 3-4 February 1992. [ Links ]
Browder, J. O., Trondoli Matricardi, E. A. & Abdala, W. S. 1996. Is sustainable tropical timber production financially viable? A comparative analysis of mahogany silviculture among small farmers in the Brazilian Amazon. Ecological Economics 16(2): 147-159.
Callaham, R. Z. 1963. Provenance research: investigation of genetic diversity associated with geography. Unasylva 18(2-3): 40-50.
Dauber, E., Guzmán, R. & Terán, J. (eds.). 1999. Potencial de los bosques naturales de Bolivia para producción forestal permanente. Superintendencia Forestal, Santa Cruz. 39 p.
Dauber, E., Terán, J. & Guzmán, R. 2003. Estimaciones de biomasa y carbono en bosques naturales de Bolivia. Revista Forestal Iberoamericana 1(1): 1-10.
Dickinson, M. B. & Whigham, D. F. 1999. Regeneration of mahogany (Swietenia macrophylla) in the Yucatan. International Forestry Review 1(1): 35-39.
Gerhardt, K. 1996. Effects of root competition and canopy openness on survival and growth of tree seedlings in a tropical seasonal dry forest. Forest Ecology and Management 82(1-3): 33-48.
Gillies, A. C. M., Navarro, C., Lowe, A. J., Newton, A. C., Hernández, M., Wilson, J. & Cornelius, J. P. 1999. Genetic diversity in Mesoamerican populations of mahogany (Swietenia macrophylla), assessed using RAPDs. Heredity 83(6): 722-732.
Griffiths, M. W. 2001. The biology and ecology of Hypsipyla shoot borers. pp. 74-80. In: Floyd, R.B. & Hauxwell, C.(eds.).Hypsipyla Shoot Borers in Meliaceae: Proceedings of an International Workshop. Australian Centre for International Agricultural Research, Kandy. [ Links ]
Grogan, J., Ashton, M. S. & Galvão, J. 2003. Big-leaf mahogany (Swietenia macrophylla) seedling survival and growth across a topographic gradient in southeast Pará, Brazil. Forest Ecology and Management 186(1-3): 311-326.
Grogan, J., Galvão, J., Simões, L. & Veríssimo, A. 2006. Regeneration of big-leaf mahogany in closed and logged forest of southeastern Pará, Brazil. pp. 193-208. In: Lugo, A. E., Figueroa, J. C. & Alayón, M. (eds.). Big-Leaf Mahogany: Genetics, Ecology, and Management. Springer Verlag, New York.
Gullison, R. E. & Hubbell, S. 1992. Regeneración natural de lamara (Swietenia macrophylla) en el Bosque Chimanes, Bolivia. Ecología en Bolivia 19: 43-56.
Gullison, R. E., Panfil, S. N., Strouse, J. J. & Hubbell, S. P. 1996. Ecology and management of mahogany (Swietenia macrophylla King) in the Chimanes Forest, Beni, Bolivia. Botanical Journal of the Linnean Society 122(1): 9-34.
Holdridge, L. R. (ed.). 1967. Life zone ecology. 2nd Edition. Tropical Science Center, USD A, San José. 206 p.
Kartawinata, K. 1994. The use of secondary forest species in rehabilitation of degraded forest lands. Journal of Tropical Forest Science 7(1): 76-86.
Krometter, R. F., Martinez, M., Blundell, A. G., Gullison, R. E., Steininger, M. K. & Rice, R. E. 2004. Impacts of unsustainable mahogany logging in Bolivia and Peru. Ecology and Society 9(1): 12 [on line] http://www.ecologyandsociety.org/vol19/iss11/art12.
Lamb, F. B. (ed.). 1966. Mahogany of tropical America: Its ecology and management. University of Michigan Press (Ann Arbor), Michigan. 220 p.
Mahroof, R. M. & Edirisinghe, J. P. 2001. Effect of quantity and quality of light on survival, growth and morphology of mahogany (Swietenia macrophylla (King)) seedlings. Journal of the National Science Foundation of Sri Lanka 29(1-2): 1-13.
Malleux, J. (ed.). 1982. Inventarios forestales en bosques tropicales. Universidad Nacional Agraria, Lima. 414 p.
Mayhew, J. E., Andrew, M., Sandom, J. H., Thayaparan, S. & Newton, A. C. 2003. Silvicultural systems for big-leaf mahogany plantations. pp. 261-277. In: Lugo, A. E., Figueroa, J. C. & Alayón, M. (eds.). Big-Leaf Mahogany: Genetics, Ecology, and Management. Springer Verlag, New York. [ Links ]
McCarter, P. S. 1986. The evaluation of the international provenance trials of Cordia alliodora and Cedrela spp. Annual Report to the UK Overseas Development Administration. Oxford Forestry Institute, Oxford. 32 p. [ Links ]
Mejía, E., Buitrón, X., Peña-Claros, M. & Grogan, J. 2008. Big-leaf mahogany (Swietenia macrophylla) in Bolivia, Peru and Brazil. Case study 4: Swietenia macrophylla. En. International Expert Workshop on CITES Non-Detriment Findings, Mexico, DF. 17-22 November 2008. [ Links ]
Merry, F. D. & Carter, D. R. 2001. Factors affecting Bolivian mahogany exports with policy implications for the forest sector. Forest Policy and Economics 2(3-4): 281-291.
Mostacedo, B. & Fredericksen, T. S. 1999. Regeneration status of important tropical forest tree species in Bolivia: assessment and recommendations. Forest Ecology and Management 124(2-3): 263-273.
Navarro, C. & Hernández, G. 2004. Progeny test analysis and population differentiation of Mesoamerican mahogany (Swietenia macrophylla). Agronomia Costarricense 28(2): 37-51.
Negreros-Castillo, P. & Hall, R. B. 1996. First-year results of partial overstory removal and direct seeding of mahogany (Swietenia macrophylla) in Quintana Roo, Mexico. Journal of Sustainable Forestry 3(2): 65-76.
Norghauer, J. M., Malcolm, J. R., Zimmerman, B. L. & Felfili, J. M. 2008. Experimental establishment of big-leaf mahogany (Swietenia macrophylla King) seedlings on two soil types in native forest of Pará, Brazil. Forest Ecology and Management 255(2): 282-291.
Novick, R. R., Dick, C. W., Lemes, M. R., Navarro, C., Caccone, A. & Bermingham, E. 2003. Genetic structure of Mesoamerican populations of big-leaf mahogany (Swietenia macrophylla) inferred from microsatellite analysis. Molecular Ecology 12: 2885-2893.
Park, A., Justiniano, M. J. & Fredericksen, T. S. 2005. Natural regeneration and environmental relationships of tree species in logging gaps in a Bolivian tropical forest. Forest Ecology and Management 217(2-3): 147-157.
Patiño Valera, F. 1997. Recursos genéticos de Swietenia y Cedrela en los Neotropicos: Propuestas para acciones coordinadas. Food and Agriculture Organization, Roma. 58 p.
Pattie, P., Nuñez, M. & Rojas, P. 2003. Valoración de los bosques tropicales de Bolivia. Documento Técnico 130/2003. BOLFOR, Santa Cruz. 44 p. [ Links ]
Pérez-Salicrup, D. R. & Esquivel, R. 2008. Tree infection by Hypsipyla grandella in Swietenia macrophylla and Cedrela odorata (Meliaceae) in Mexico's southern Yucatan Peninsula. Forest Ecology and Management 255(2): 324-327.
Pinto, J. 2006. Evolución del paisaje y estado de conservación de la Reserva Forestal El Choré. Kempffiana 2(1): 45-56.
Piotto, D., Víquez, E., Montagnini, F. & Kanninen, M. 2004. Pure and mixed forest plantations with native species of the dry tropics of Costa Rica: a comparison of growth and productivity. Forest Ecology and Management 190(2-3): 359-372.
Quevedo, L. H. 1986. Evaluación del efecto de la tala selectiva sobre la renovación de un bosque humedo subtropical en Santa Cruz, Bolivia. Tesis de maestría, CATIE Universidad de Costa Rica, Turrialba. 221 p. [ Links ]
Snook, L. K. 2003. Regeneration, growth, and sustainability of mahogany in Mexico's Yucatan forests. pp. 169-192. In: Lugo, A. E., Figueroa, J. C. & Alayón, M. (eds.). Big-Leaf Mahogany: Genetics, Ecology, and Management. Springer Verlag, New York. [ Links ]
Snook, L. K. & Negreros-Castillo, P. 2004. Regenerating mahogany (Swietenia macrophylla King) on clearings in Mexico's Maya forest: the effects of clearing method and cleaning on seedling survival and growth. Forest Ecology and Management 189(1-3): 143-160.
Varmola, M. I. & Carle, J.B. 2002. The importance of the hardwoods plantations in the tropics and sub-tropics. International Forestry Review 4(2): 110-121.
Veríssimo, A., Barreto, P., Tarifa, R. & Uhl, C. 1995. Extraction of a high-value natural resource in Amazonia: the case of mahogany. Forest Ecology and Management 72(1): 39-60.
Wadsworth, F. H. & González, E. 2008. Sustained mahogany (Swietenia macrophylla) plantation heartwood increment. Forest Ecology and Management 255(2): 320-323.
Ward, S. & Lugo, A. 2003. Twenty mahogany provenances under different conditions in Puerto Rico and the U.S. Virgin Islands, pp. 103-117. In: Lugo, A., Figueroa, J. C. & Alayón, M. (eds.). Big-Leaf Mahogany: Genetics, Ecology, and Management. Springer Verlag, New York.
Weaver, P. L. & Bauer, G. P. 1986. Growth, survival and shoot borer damage in mahogany plantings in the Luquillo Forest in Puerto Rico. Turrialba 36(4): 509-522.
Wormald, T. J. 1992. Mixed and pure forest plantations in the tropics and subtropics. Food and Agriculture Organization, Rome. 166 p. [ Links ]
Artículo recibido en: Noviembre de 2011.
Manejado por: Blanca León
Aceptado en: Marzo de 2012.

