










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkEcología en Bolivia
Print version ISSN 1605-2528On-line version ISSN 2075-5023
Ecología en Bolivia vol.45 no.2 La Paz Sept. 2010
Artículo
Sistemas de polinización de cinco especies de orquídeas creciendo bajo condiciones de invernadero
Pollination systems of five species of orchids growing under greenhouse conditions
Dennise Quiroga, Marciana Martínez & Daniel M. Larrea-Alcázar*
Departamento de Ciencias, Fundación Amigos de la Naturaleza (FAN-Bolivia), Km. 7 ½ Doble Vía a La Guardia, Telf. +591-3-3556800, Fax. +591-3-3547383, Santa Cruz, Bolivia
*Autor para correspondencia: dlarrea@fan-bo.org, larrea.alcazar@gmail.com
La variedad de formas y colores de las flores de orquídeas es quizás el aspecto más conocido y documentado de estas plantas (Gravendeel et al. 2004). Sus colores llamativos, la marcada simetría bilateral de sus flores, su hermafroditismo (rara vez unisexuado), junto con el desarrollo de estructuras especializadas, como el labelo y sus particulares polinios, han originado que sean comúnmente mencionadas como ejemplos de plantas con sistemas de entrecruzamiento altamente especializados (evidencia indirecta de selección natural, Fægri & van der Pijl 1979). Hoy, sabemos que la alta especialización de sus flores no corresponde en todos los casos a la visita de un solo polinizador, dando lugar a sistemas generalizados con varias especies de polinizadores (Chase & Hills 1992, Herrera & Pellmyr 2002, Fenster et al. 2004). Existen diferencias de una especie a otra en la duración de la antesis y en el periodo de floración que sugieren variaciones en la composición y efecto de los visitantes sobre las flores (Kindlmann & Balounová 1999). Igualmente, otras especies han mostrado ser autocompatibles (e.g. Zimmerman & Aide 1989), lo que sugiere que la autogamia es un sistema de entrecruzamiento que ocurre con cierta frecuencia en el grupo. Estas tendencias han sido poco exploradas en condiciones de invernadero y constituyen aún preguntas abiertas para gran parte de las especies neotropicales.
Para Bolivia, se estima que la riqueza potencial de orquídeas alcanza entre 2.000 y 3.000 especies, de las cuales 1.200 han sido identificadas y descritas (Ibisch et al. 2003, Vásquez & Ibisch 2004a, b). La distribución y riqueza del grupo está fuertemente asociada con la ecorregión de Yungas (Ibisch et al. 2003, Müller et al. 2003), una región que representa cerca del 4% de la superficie del país y que concentra cerca del 60% de las especies conocidas (principalmente especies de las tribus Laeliinae, Polystachyinae, Sobraliinae y Pleurothallidinae, ver Ibisch et al. 2007), de las cuales el 80% es endémico a Bolivia (Vásquez & Ibisch 2004a). Mientras el conocimiento sobre la taxonomía, riqueza y distribución del grupo ha ido en aumento (Vasquez & Ibisch 2000, 2004a, b, Müller et al. 2003, Vásquez et al. 2003, Krömer et al. 2007), junto al interés con fines ornamentales de sus especies, otros aspectos sobre la ecología de sus especies, en especial los sistemas de entrecruzamiento, son aún poco conocidos. Esta información puede ayudar a comprender las pequeñas, discontinuas y dispersas poblaciones que en general presentan sus especies, lo cual parece beneficiar la deriva génica (Ackerman & Ward 1999). Igualmente, esta información puede ser utilizada para favorecer la variabilidad genética de las especies que son cultivadas (Tibbs 2007) o aquellas que son propagadas en condiciones in vitro para su comercialización (J. Quezada et al., datos no publicados).
El objetivo de este estudio fue analizar los sistemas de entrecruzamiento de Cattleya luteola Lind., Brassia thyrsodes Rchb., Prosthechea vespa (Vell.) W.E. Higgins, Schomburgkia undulata Lind. y Xylobium variegatum (Ruiz & Pav.) Garay & Dunst. bajo condiciones de invernadero. El estudio examinó la eficiencia de la polinización cruzada en la formación de frutos y sus posibles implicaciones en las características reproductivas de las especies analizadas. Este es uno de los primeros reportes escritos que examina la biología de la polinización de especies de orquídeas nativas de Bolivia; no obstante, existen experiencias preliminares realizadas con especies del género Masdevallia Ruíz & Pavón (J. Quezada et al., datos no publicados).
Los experimentos de entrecruzamiento fueron realizados entre febrero y diciembre de 2009 para determinar si las flores de las cinco especies estudiadas son autocompatibles. Los experimentos permitieron contrastar especies que pertenecen a distintas subtribus y que, en condiciones naturales, crecen en ecorregiones y pisos de vegetación distintos (Tabla 1).
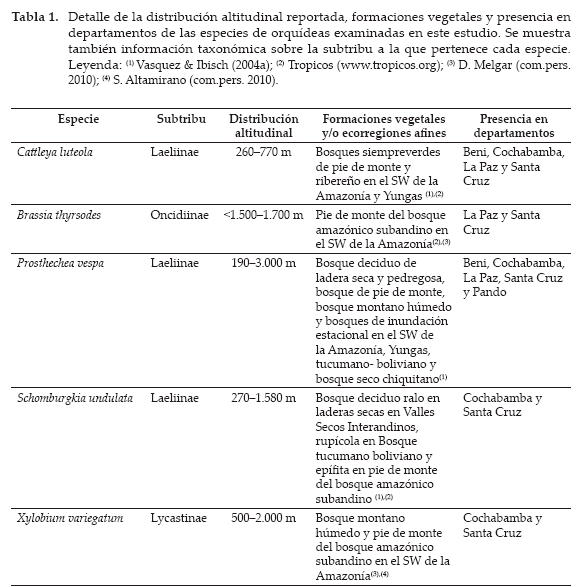
El experimento permitió también registrar datos fenológicos de las especies estudiadas (11 meses). Las cantidades de flores e inflorescencias y el número de individuos y procedencia de los especímenes utilizados en los experimentos son detallados en la Tabla 2.
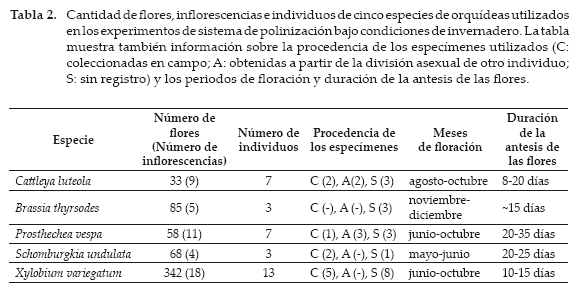
Entre 2-129 flores de cada especie fueron asignadas a los siguientes tratamientos:
1) Autopolinización autónoma (autogamia): Entre 3-129 botones florales fueron protegidos con bolsas de tol y monitoreados hasta que abortaron o formaron frutos;
2) Autopolinización asistida (geitogamia): Entre 8-68 botones florales fueron embolsados en bolsas de tol; cuando se abrieron fueron polinizados manualmente usando polen fresco de la misma flor;
3) Polinización cruzada (xenogamia): Entre 2-12 botones florales fueron protegidos con bolsas de tol; cuando estos se abrieron fueron polinizados manualmente usando polen de flores de inflorescencias de distintos individuos de la misma especie;
4) Polinización “natural” (control): Entre 8-122 botones florales fueron marcados sin embolsar y monitoreados hasta que abortaron o formaron frutos. Este tratamiento no se consideró como estrictamente “natural” debido a que el invernadero restringió la llegada de visitantes florales y que las condiciones climáticas hacia el interior del invernadero no son necesariamente equivalentes a aquellas que se presentan los hábitats donde crecen las especies estudiadas.
Todos los experimentos fueron realizados en el invernadero de la colección científica de Plantas Vivas de la Fundación Amigos de la Naturaleza (FAN-Bolivia). Las especies fueron escogidas en función de la disponibilidad de individuos en el invernadero (min: 3 ind en S. undulata, max: 13 ind en X. variegatum). Debido a la baja cantidad de individuos disponibles para realizar los experimentos de polinización cruzada, para S. undulata se obtuvo polen de flores de individuos fértiles ubicados en el Jardín Botánico Municipal de Santa Cruz. La temperatura en el invernadero, en promedio, osciló entre 18 (agosto) y 26°C (octubre y diciembre) (24°C ± 5.1; media ± DE), mientras que la humedad relativa osciló entre 37 (agosto) y 64 (julio) % (56.8% ± 27.1, datos registrados con dos sensores HOBO (Modelo H08-004-02) que funcionaron durante tres días continuos por cinco meses. Para cada especie, las diferencias entre las proporciones para los tratamientos con frutos formados fueron analizadas mediante pruebas de proporciones. Todos los análisis fueron realizados en el programa Statistica, versión 6.0 (StatSoft 2001).
La floración de las especies estudiadas se inició en mayo (S. undulata) y culminó en diciembre (B. thyrsodes). El periodo de floración fue de solo un par de meses en S. undulata, mientras que en P. vespa y X. variegatum alcanzó los cinco meses. Individualmente, las flores permanecieron abiertas entre ocho (C. luteola) y 35 días (P. vespa) (Tabla 2).
En ninguna de las especies, los experimentos de polinización “natural” formaron frutos. Igualmente, las flores de B. thyrsodes no formaron frutos en ninguno de los tratamientos (Tabla 3).
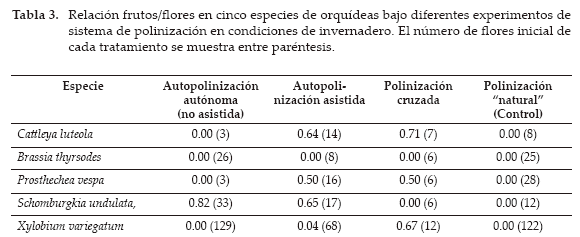
En C. luteola, P. vespa y X. variegatum los experimentos de geitogamia (autopolinización asistida) y xenogamia (polinización cruzada) produjeron frutos. Entre ellas, únicamente para X. variegatum la proporción de frutos formados por polinización cruzada fue significativamente mayor que aquella formada a través de autopolinización asistida (prueba de proporciones; p<0.01), lo que sugiere que presenta un sistema de polinización marcadamente xenogámico. Estas diferencias no fueron detectadas en C. luteola y P. vespa (p=0.75 y p=0.99, respectivamente). Ambas especies parecen presentar sistemas reproductivos combinados o mixtos (geitogamia y xenogamia). La ausencia de formación de frutos en los tratamientos de autogamia apunta a que ambas especies podrían presentar limitaciones de polinizadores para favorecer la formación de sus frutos.
En el caso de S. undulata, los experimentos de autogamia y geitogamia formaron frutos (Tabla 3). La proporción de frutos formados entre ambos tratamientos no fue significativamente distinta (prueba de proporciones; p=0.18), lo que sugiere que no existiría limitación de polinizadores para favorecer la geitogamia en esta especie. La ausencia de formación de frutos en el tratamiento de xenogamia podría, sin embargo, explicarse por el polen utilizado en el experimento, el cual pudo sufrir algún deterioro en su transporte desde el Jardín Botánico de Santa Cruz.
Los resultados sugieren que tres de las cinco especies examinadas formaron frutos vía polinización cruzada. Entre ellas, X. variegatum parece ser una especie fuertemente autoincompatible (xenógama), mientras C. luteola y P. vespa exhibirían sistemas de entrecruzamiento combinados entre autogamia y geitogamia, lo que muestra que ambas especies son parcialmente autocompatibles. Las otras dos especies de orquídeas mostraron sistemas de entrecruzamientos distintos. Mientras S. undulata parece ser una especie principalmente autocompatible (autógama), la formación de frutos en B. thyrsodes por vía sexual parece ser poco probable y, posiblemente, el mantenimiento de sus poblaciones en condiciones naturales dependa de la capacidad de propagación vegetativa de esta especie. No obstante, la ausencia de formación de frutos en esta especie pudo ser resultado del corto periodo de receptividad estigmática. Futuros estudios podrían determinar la relación entre receptividad estigmática y la antesis de las flores de esta especie (< 15 días). Para algunas especies del género Brassia se ha reportado que la polinización es llevada a cabo por avispas del género Pepsis Fabricius (Fam. Pompilidae, Dodson 1965) y que el género en sí se caracteriza por presentar especies con fuerte afinidad a la polinización por engaño (Ospina-Calderón et al. 2007).
Las dos especies que mostraron sistemas reproductivos combinados (C. luteola y P. vespa) son especies epífitas de la subtribu Laeliinae, una de las subtribus con mayor cantidad de especies presentes el país (Vásquez et al. 2003, Vásquez & Ibisch 2004a). Poco se conoce sobre la proporción de las especies de esta subtribu que expresan este tipo de sistema de entrecruzamiento o su relación con el hábito epífito lo que, sin embargo, explicaría parcialmente la alta diversificación de este grupo (sensu Gravendeel et al. 2004). Este tipo de enfoques (sistema de polinización versus hábito) ha sido poco explorado en la investigación de las orquídeas presentes en nuestro país.
En condiciones naturales, las flores de C. luteola son polinizadas por abejas, las que son atraídas por el aroma dulce que desprenden las flores durante el inicio de la mañana (Tibbs 2007). Al parecer esta especie no produce néctar como recompensa para sus visitantes florales (Vásquez & Ibisch 2004a), lo que tiende a ser una estrategia relativamente generalizada en el grupo de las orquídeas que permite visitas a pocas flores pero mayor carga de polen por parte de sus polinizadores (sistema de polinización “por engaño”; Ackerman 1986, Smithson 2006). Al contrario, aunque es conocida la producción de néctar en especies del género Prosthechea (Higgins 2003), su presencia en P. vespa aún no ha sido reportada. Sin embargo, las guías de néctar en sus flores indican su presencia. La autopolinización también ha sido reportada en el género Prosthechea, e incluso algunas especies tienen la estructura floral adaptada para este tipo de reproducción (Higgins 2003). Tanto en C. luteola como en P. vespa, la formación de frutos vía geitogamia es posible, siempre que cuenten con los servicios de visitantes florales que transporten el polen entre las flores de una misma inflorescencia.
Esta potencial demanda de polinizadores al parecer no es necesaria para S. undulata – la otra especie estudiada de la subtribu Laeliinae también de hábito epífito – cuyas flores formaron frutos principalmente por vía autogámica. Al contrario, tal necesidad parece ser altamente indispensable para la formación de frutos en X. variegatum – especie de hábito epífito de la subtribu Lycastinae – que formó frutos esencialmente vía polinización cruzada.
En Perú y Costa Rica la especie es polinizada por abejas del género Trigona (T. amalthea, Van der Pijl & Dodson 1966). Este resultado es por demás sugestivo si consideramos que las especies del género Xylobium se caracterizan por la ausencia de la producción de néctar de sus flores. Este patrón sugiere, nuevamente, que el sistema de polinización “por engaño” no es una excepción en el grupo (sensu Roubik 2000).
En resumen, este estudio muestra que es posible detectar diferentes sistemas de entrecruzamiento – desde estricta hasta parcialmente autocompatibles – en especies de orquídeas que crecen bajo condiciones de invernadero. Es plausible pensar que los resultados presentados pudieran arrojar tendencias distintas si son realizados en el lugar de crecimiento de cada especie. Sin embargo, los resultados muestran la importancia de la polinización cruzada y la autopolinización asistida para la formación de los frutos. Estos datos proporcionan pautas para el manejo y propagación en invernadero de las especies estudiadas y resaltan la necesidad de mayores investigaciones con otras especies que son conservadas o comercializadas bajo esta estrategia. Entre las especies examinadas en este estudio, C. luteola es la especie más usada con estos fines.
Agradecimientos
Este trabajo fue realizado con fondos otorgados por la Embajada Real de los Países Bajos a la Fundación Amigos de la Naturaleza (FAN-Bolivia). Agradecemos a Darío Melgar Gómez, Director del Jardín Botánico Municipal Santa Cruz por facilitar el polen para el experimento de polinización cruzada de una de las especies estudiadas (S. undulata). Pierre Ibisch, Ramiro López y dos revisores anónimos hicieron valiosos comentarios a las versiones preliminares del manuscrito. Saúl Altamirano proporcionó datos importantes para X. variegatum.
Referencias
Ackerman, J. D. 1986. Mechanisms and evolution of food-deceptive pollination systems in orchids. Lyndleyana 1: 108-113.
Ackerman, J.D. & S. Ward. 1999. Genetic variation in a widespread, epiphytic orchid: where is the evolutionary potential? Systematic Botany 24: 282-291.
Dodson, C.H. 1965. Studies in Orchid pollination: the genus Coryanthes. American Orchid Society Bulletin 34: 680-687.
Chase, M.W. & H.G. Hills. 1992. Orchid phylogeny, flower sexuality, and fragrance-seeking. BioScience 42: 43-49.
Fægri, K. & L. van der Pijl. 1979. The principles of pollination ecology. Pergamon Press. Oxford, 244 p. [ Links ]
Fenster, C.B., W.S. Armbruster, P. Wilson, M.R. Dudash & J. D. Thomson. 2004. Pollination syndromes and floral specialization. Annual Review of Ecology, Evolution, and Systematics 35: 375-403.
Gravendeel, B., A. Smithson, F.J. Slik & A. Schuiteman. 2004. Epiphytism and pollinator specialization: drivers of orchid diversity? Philosophical Transactions of The Royal Society, Biological Sciences 359:1523-1535.
Higgins, W.E. 2003. Prosthechea: a chemical discontinuity in Laeliinae. Lankesteriana 7: 39-41.
Ibisch, P.L., S.G. Beck, B. Gerkmann & A., Carretero. 2003. La diversidad biológica: ecoregiones y ecosistemas. Pp. 47-88. En P.L. Ibisch & Mérida, G. (eds.). Biodiversidad: La riqueza de Bolivia. Editorial Fundación Amigos de la Naturaleza (FAN), Santa Cruz, 638 p. [ Links ]
Ibisch, P.L., N. Araujo & C., Nowicki. 2007. Visión de Conservación de la Biodiversidad del Corredor Amboró – Madidi. Editorial Fundación Amigos de la Naturaleza (FAN), Santa Cruz, 414 p.
Herrera, C.M. & O. Pellmyr. 2002. Plant-animal interactions. An evolutionary approach. Blackwell, Oxford, 313 p. [ Links ]
Kindlmann, P. & Z. Balounová. 1999. Flowering regimes of terrestrial orchids: unpredictability or regularity? Journal of Vegetation Science 10: 269-273.
Krömer, T., S.B. Gradstein & A. Acebey. 2007. Diversidad y ecología de epífitas vasculares en bosques montanos primarios y secundarios de Bolivia. Ecología en Bolivia 42: 23-33.
Müller, R., C. Nowicki, W. Barthlott & P.L. Ibisch. 2003. Biodiversity and endemism mapping as a tool for regional conservation planning – case study of the Pleurothallidinae (Orchidaceae) of the Andean rain forests in Bolivia. Biodiversity and Conservation 12: 2005-2024.
Ospina-Calderón, N., M. Diaz-Granados & P.Viveros-Bedoya. 2007. Observaciones de la polinización y fenología reproductiva de Brassia cf. antherothes RCHB.F. (Orchidaceae) en un relicto de selva subandina en la Reserva natural La Montaña. Pontificia Universidad Javeriana, Bogotá, 95 p. [ Links ]
Roubik, D.W. 2000. Deceptive orchids with Meliponini as pollinators. Plant Systematics and Evolution 222: 271-279.
Smithson, A. 2005. Pollinator limitation and inbreeding depression in orchid species with and without nectar rewards. New Phytologist 169: 419-430.
StatSoft, Inc. 2001. STATISTICA (data analysis software system), version 6. [ Links ]
Tibbs, M. 2007. Orchids: A practical guide to care and cultivation. Ball Publishing, London, 160 p. [ Links ]
Van der Pijl, L. & C.H. Dodson. 1966. Orchids flowers, their pollination and evolution. University of Miami Press, Miami, 404 p. [ Links ]
Vásquez, R. & P.L. Ibisch. 2000. Orquídeas de Bolivia/Orchids of Bolvia. Diversidad y estado de conservación/Diversity and conservation status. Vol.1. Pleurothallidinae. Editorial FAN, Santa Cruz, 548 p.
Vásquez, R. & P.L. Ibisch. 2004a. Orquídeas de Bolivia/Orchids of Bolvia. Diversidad y estado de conservación/Diversity and conservation status. Vol.2. Laeliinae, Polystachinae, Sobralinae con actualización y complementación de/with update and complementation of Pleurothallidinae. Editorial FAN, Santa Cruz, 649 p.
Vásquez, R. & P.L., Ibisch. 2004b. Nuevas especies de orquídeas de Bolivia: 4. Revista de la Sociedad Boliviana de Botánica 4: 31-39.
Vásquez, R., P.L. Ibisch & B. Gerkmann. 2003. Diversity of Bolivian Orchidaceae – a challenge for taxonomic, floristic and conservation research. Organism Diversity & Evolution 3: 93-102.
Zimmerman, J.K. & T. M. Aide.1989. Patterns of fruit production in a Neotropical orchid: pollinator vs. resource limitation. American Journal of Botany 76: 67-73.
Nota recibida en: Enero de 2010.
Manejada por: Mónica Moraes R.
Aceptada en: Mayo de 2010.

